

核糖体工程通过在细胞水平上改善蛋白质合成来重塑培养肉生产。核糖体是细胞的蛋白质工厂,对于生产定义肉质地和营养价值的肌动蛋白、肌球蛋白和其他蛋白质至关重要。然而,标准细胞系并未针对大规模肉类培养所需的高生产力进行优化。
关键进展包括:
- 优化的核糖体RNA变体: 筛选含有1.7 × 10⁷个变体的文库显示出增加翻译活性的潜力。
- 正交核糖体: 这些工程化的核糖体专门生产特定蛋白质,如肌球蛋白,而不干扰正常的细胞功能。
- 密码子优化: 根据核糖体偏好调整mRNA序列,蛋白质表达量提高了多达72倍。
- 肌肉因子信号传导: 像IL-15和肌动素这样的蛋白质在肌肉分化过程中增强核糖体生物合成和蛋白质合成。
在平衡能量需求、维持细胞稳定性和将生产规模扩大到工业水平方面仍然存在挑战。例如,核糖体过度活跃可能导致蛋白质错误折叠或代谢压力,而生物反应器中的营养扩散限制会限制组织生长超过200微米。解决这些问题需要将核糖体工程与先进的生物加工策略相结合。
本文探讨了这些方法如何塑造培养肉的未来以及必须克服的障碍以实现商业可行性。
核糖体与蛋白质生物合成:入门指南
哺乳动物细胞中的核糖体结构和功能
核糖体是蛋白质合成的核心,将mRNA序列翻译成功能性蛋白质。在哺乳动物细胞中,核糖体被分类为80S颗粒,由两个亚基组成:40S小亚基,负责解码mRNA,以及60S大亚基,负责催化肽键的形成。翻译过程包括三个主要步骤:起始, 识别起始密码子;延伸, 氨基酸依次添加到正在增长的多肽链中;以及终止, 当达到终止密码子时发生。
大亚基的两个特定区域对于工程应用尤为重要:肽酰转移酶中心 (PTC), 促进肽键形成,以及出口通道, 新合成的多肽通过此通道退出[3].
掌握这些核心机制对于探索如何优化核糖体性能以提高培养肉生产至关重要。
为什么蛋白质生物合成对培养肉很重要
蛋白质合成的效率是培养肉开发中的一个关键因素,特别是在体外肌生成过程中。这个过程将肌肉卫星细胞(MSCs)转化为富含收缩蛋白如肌动蛋白和肌球蛋白的多核肌纤维。核糖体在这一转化中起着核心作用[4].
“大约需要八万亿个肌肉细胞才能从一个容量为5,000升的传统生物反应器中生产出1公斤的蛋白质”[5]
这一惊人的需求突显了即使是核糖体效率的小幅提高也能显著提升产量,直接影响培养肉的商业可行性。
随着细胞的成熟,它们的核糖体活动会发生变化。在增殖阶段,MSCs优先进行快速分裂。然而,在分化的三到五天后,重点转向合成收缩蛋白的成人同工型,并促进细胞融合成肌管[4]. 这一转变由特定的信号分子或肌肉因子调节。
例如,白细胞介素-15 (IL-15)促进肌球蛋白重链 (MyHC) 蛋白的积累,同时减少蛋白质降解,作为肌肉发育过程中的关键合成代谢因子 [4]. 同样,Myonectin通过PI3K/Akt/mTOR信号通路增强蛋白质合成来支持肌肉生长 [4]. 了解这些信号通路如何影响核糖体活动对于设计满足生产需求的可扩展细胞系至关重要。这些见解为后续章节中讨论的工程策略奠定了基础。
当前核糖体工程研究
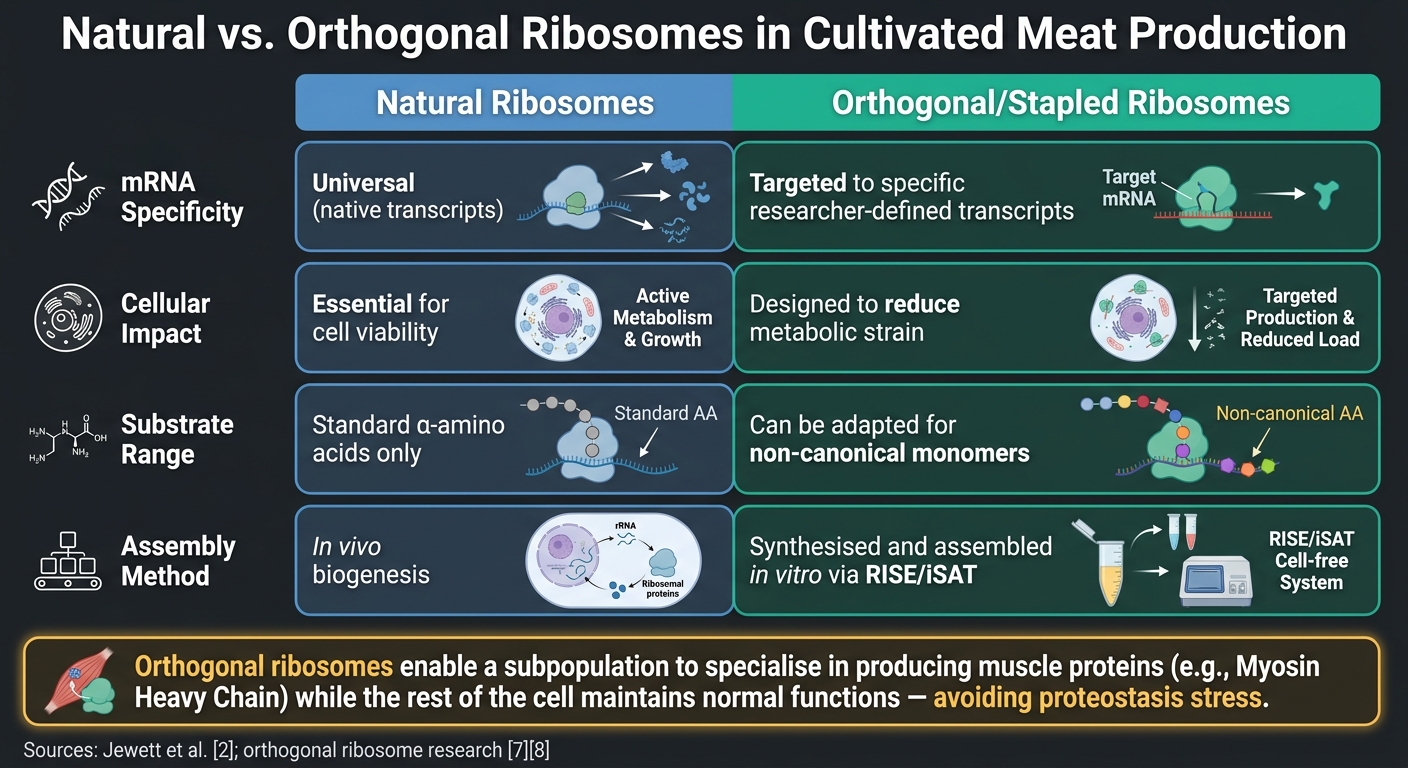
天然核糖体与正交核糖体在培养肉生产中的应用
核糖体生物发生与翻译控制
核糖体生物发生是细胞构建新核糖体的过程,是一个高度调控且耗能的活动。在哺乳动物细胞中,它占据了细胞代谢输出的很大一部分。仅翻译过程就可能消耗多达75%的细胞总能量预算 [8], 使其成为最耗资源的细胞过程之一。
当核糖体分配效率低下时——例如,当核糖体在早期编码区域停滞时——会造成瓶颈,减少自由核糖体的可用性,最终限制蛋白质的生产。计算模型显示,通过工程化处理仅100个基因可以提高酵母(Saccharomyces cerevisiae)的核糖体分配35%,以及在 大肠杆菌中提高 57%[8]. 。这些发现对优化哺乳动物细胞中的核糖体动态具有直接影响,特别是在培养肉行业中,能源效率和蛋白质产出至关重要。
培养肉背景下的核糖体工程
核糖体工程的进展现在正应用于培养肉生产,建立在核糖体生物发生的基础知识之上。即使是未直接在肌肉细胞中进行的研究,也在为培养肉细胞系提供相关见解。
2020年12月,Hadas Zur 和 Tamir Tuller 来自特拉维夫大学展示了 核糖体交通工程 (RTE)在提高生长率和蛋白质产量方面的潜力。使用 CRISPR-Cas9, 他们在RPO21和 CYS4的坡道区域(密码子11-50)中引入了同义突变在 酿酒酵母. 中。结果双突变体表现出改进的对数生长期生长和细胞密度。然而,研究人员警告说,在二相生长和静止期,翻译优化与生长率之间的关系减弱,此时翻译以外的因素成为限制因素[8]. 这一见解对于设计培养肉生产中的分化协议特别重要。
2020年2月,Michael Jewett的团队在西北大学验证了RISE(体外合成和进化核糖体) 方法。该技术涉及筛选大约1.7 × 10⁷核糖体RNA变体 [2] . 通过完全在活细胞外操作,RISE绕过了致命核糖体突变所施加的限制,这些突变无法在体内研究。
“体外方法克服了细胞活力的限制,使得探索致命核糖体突变成为可能。” - Michael Jewett等人 [2]
另一个对培养肉有前景的创新是使用正交核糖体. 这些工程化的核糖体-mRNA对独立于细胞的天然翻译机制运作。这使研究人员能够将核糖体活动集中在特定目标上,例如对肌肉质地至关重要的肌球蛋白重链(MyHC)亚型,而不干扰基本的细胞过程 [6]. 比较研究强调了正交核糖体相对于天然核糖体的优势:
| 特征 | 天然核糖体 | 正交/订书钉核糖体 |
|---|---|---|
| mRNA 特异性 | 通用(天然转录本) | 针对特定研究人员定义的转录本 [6] |
| 细胞影响 | 对生存至关重要 | 设计以减少代谢负担 [7] |
| 底物范围 | 标准 α-氨基酸 | 可适用于非标准单体 [7] |
| 组装 | 体内生物发生 | 在体外通过RISE/iSAT合成和组装[2] |
这里的关键点是正交核糖体使一部分核糖体专门生产肌肉蛋白,如MyHC,而细胞的其余部分保持正常功能。这避免了蛋白质稳态压力的风险,这种压力可能在整个翻译系统被推动过度生产特定蛋白质时出现。
提高核糖体性能的策略
增加核糖体生物合成
增加核糖体数量是增强蛋白质生产的直接方法,已有两种主要方法引起了关注。第一种方法涉及修改核糖体RNA(rRNA)基因的表观遗传状态以提高其翻译能力。
"核糖体RNA基因的表观遗传工程增强了蛋白质生产。" - Santoro R., Lienemann P., Fussenegger M. [1]
第二种方法利用PI3K/Akt/mTOR信号通路。肌肉因子如IL-15、肌动素和鸢尾素激活该通路,在肌管成熟过程中驱动核糖体生物合成,如前所述。
然而,核糖体生产的增加必须与细胞的代谢能力仔细平衡,因为核糖体合成是活细胞中最耗能的过程之一[1].
一旦核糖体数量增加,重点就转向确保它们完全参与翻译。
改善翻译起始和延长
最大化所有核糖体的活性是至关重要的,因为即使在生长优化的细胞中,仍有15-20%的核糖体处于非活跃状态[9]. 这代表了培养肉细胞系中未开发能力的显著储备。
翻译延长的速度取决于两个因素:核糖体的固有速度和积极参与翻译的核糖体比例[9]. 为了优化这些,保持培养基中高水平的氨基酸是至关重要的。此外,工程化细胞系以稳定核糖体蛋白有助于保护rRNA免于错误折叠和降解,从而减少在高峰生长条件下rRNA的典型10%损失[9].
一旦核糖体活性达到最大化,优化mRNA序列成为加速蛋白质合成的下一步。
mRNA优化和密码子使用
核糖体的性能高度依赖于它们处理的mRNA的质量。密码子优化将目标蛋白的编码序列调整为与宿主物种特定的tRNA库对齐 - 例如牛、猪或鱼。这种对齐防止了延长过程中核糖体的停滞,并增加了对关键肌源性蛋白如MyoD和Myf5的吞吐量。
除了密码子优化,转录调节确保了细胞内rRNA和mRNA水平的适当平衡。这些组件之间的任何不匹配都可能造成瓶颈,降低整体效率[1].
在实际应用中,集成合成、组装和翻译(iSAT)系统提供了一个有价值的工具。这些系统使用无细胞提取物和基于荧光的检测方法在体外原型化优化的mRNA,然后将其整合到稳定的细胞系中。这种迭代方法使研究人员能够快速比较密码子优化的变体,提高重要肌源性蛋白的产量,并增强培养肉生产的可扩展性[1].
权衡:生长、分化和产品质量
优化核糖体性能涉及在提高蛋白质合成和管理对细胞生长和分化的影响之间取得微妙的平衡,如前所述。
代谢负担和蛋白质稳态压力
工程化核糖体以增强蛋白质生产会带来更高的能量需求,因为它将ATP和氨基酸从其他重要的细胞功能中转移出来。核糖体合成已经是细胞内最耗能的过程之一,进一步的放大可能会加剧这些能量挑战。
这种强化活动也可能影响蛋白质质量。过度活跃的核糖体可能会使细胞伴侣蛋白不堪重负,导致蛋白质错误折叠并激活未折叠蛋白反应(UPR)。这种压力可能会抑制生长,甚至导致细胞死亡。对于像牛或羊这样的牲畜物种的成年干细胞,它们自然具有有限的增殖能力,这些额外的压力可能会显著减少在衰老开始之前可行的细胞分裂次数[5].
在培养肉生产中,由于营养扩散的限制,组织厚度很少超过200 μm,这可能导致较大组织聚集体核心的细胞死亡[5]. 增加能量消耗的策略有加速这些关键区域营养耗尽的风险,在这些区域中,持续的蛋白质合成是必不可少的。此外,增加的代谢压力可能干扰肌肉分化所需的精细信号通路。
对肌肉分化和蛋白质组成的影响
核糖体工程引入的压力可能超越代谢,潜在地扰乱肌肉发育。肌肉形成过程,即肌生成,依赖于一系列严格调控的转录因子:Pax7确保干细胞保持静止,Myf5促进肌母细胞的增殖,而MyoD 触发分化[5] . 改变蛋白质合成可能会扰乱这一序列,阻碍分化或产生非典型的肌纤维组成。这可能导致肌内脂肪沉积减少,而肌内脂肪沉积是实现培养肉理想质地和风味的关键 [5].
因此,通过监测肌生成标志物的表达来保持严格的质量控制,对于确保肌肉的正常发育和产品质量至关重要。
sbb-itb-ffee270
研究空白和未来方向
核糖体工程的进展显示出希望,但其在商业化培养肉生产中的应用仍面临重大障碍。为了弥合这些差距,研究人员需要专注于先进的分子分析技术和可扩展的生物工艺策略,以承受长期生产的需求。
多组学和长期稳定性研究
一个主要挑战在于缺乏工程化细胞系的长期稳定性数据。随着时间的推移,这些细胞可能会积累自发突变,可能改变其表型。来自诺维萨德大学的伊万娜·帕金强调了这一问题:永生化细胞“由于长期培养期间可能发生的自发突变,并不总是代表原代培养物”[13]. 对于核糖体工程化的细胞系,风险更高——核糖体成分的突变可能会在未被立即检测到的情况下削弱翻译效率。
多组学方法提供了一种解决这些问题的方法。通过整合转录组学、蛋白质组学和代谢组学,研究人员可以监测关键的肌源性标志物,如Pax7, MyoD, 和Myogenin, 以及MyHC同工型的变化。然后,基因组规模的代谢模型可以将这些见解转化为可操作的培养基成分变化,以满足工程化核糖体的独特需求[5][11]. 对于培养肉,确保在延长周期内持续的蛋白质生产是至关重要的。没有这样的纵向监测,很难将可持续的改进与短暂的效果区分开来。
除了遗传和代谢稳定性之外,将这些创新扩展到工业水平也带来了自身的一系列挑战。
生物工艺集成与放大
将核糖体工程细胞从小烧瓶扩展到工业生物反应器绝非易事。在一个5,000升的搅拌罐生物反应器中生产仅1公斤的蛋白质需要大约八万亿个肌肉细胞[5]. 在这些密度下,营养梯度成为一个关键问题。氧气和其他营养物质的200微米扩散极限意味着在3D组织结构核心的细胞可能面临饥饿,特别是在由于高蛋白质合成而对资源需求达到峰值时。
生物反应器搅拌产生的剪切应力增加了复杂性。虽然未修改的细胞可能能够忍受这种湍流,但具有修改翻译机制的工程细胞可能更容易受到影响。压力不仅可能破坏细胞通路,还可能对已经处于代谢压力下的细胞造成物理损伤 [13]. 解决这些问题需要将实时数据与数字生物制造模型相结合,包括计算流体动力学模拟,以更好地理解和预测大型容器内的多样化微环境 [10]. 下游过程如收获也需要关注 - 涉及胰蛋白酶的酶法可能会改变工程细胞的表面蛋白质组 [14], 可能抵消核糖体工程的好处。
| 放大因子 | 关键瓶颈 | 与核糖体工程的相关性 |
|---|---|---|
| 营养扩散 | 200 μm 渗透极限 [5] | 可能导致在3D组织中具有高蛋白合成需求的细胞饥饿 |
| 遗传稳定性 | 自发突变 [13] | 可能随着时间的推移损害工程化的翻译效率 |
| 剪切应力 | 搅拌罐湍流 [13] | 有风险破坏工程化的细胞通路 |
| 收获方法 | 胰蛋白酶的蛋白水解损伤 [14] | 可能改变蛋白质组并掩盖蛋白质质量的改进 |
解决这些规模化挑战对于将核糖体工程从实验室转化为商业生产至关重要。每个策略都必须经过严格测试,以确保在工业条件下获得可靠的蛋白质产量、稳定性和安全性。
结论:培育肉中核糖体工程的理由
在5,000升生物反应器中生产1公斤蛋白质需要惊人的8万亿肌肉细胞[5]. 这突显了扩大培育肉生产规模的巨大挑战。核糖体工程通过提高单个细胞的蛋白质产量,而不是简单地增加细胞数量,提供了解决方案。
在应用核糖体工程时,时机至关重要。在错误阶段增强翻译可能会扰乱肌生成,可能影响关键收缩蛋白如MyHC的生产[5]. 在翻译和肌生成之间取得正确的平衡与工程本身同样重要。
“为了实现高质量的CBM及其高产量生产,分子方面需要进行彻底检查,以实现商业生产的良好实验室实践。” - Asim Azhar 等人,食品科学与技术前沿 [5]
已经有几种技术在增加重组蛋白产量方面显示出前景,例如过表达翻译起始因子(eIF3i 和 eIF3c)、密码子优化和靶向 mRNA 修饰 [15]. 然而,这些方法必须谨慎应用,以避免代谢负担、蛋白质稳态压力和长期遗传不稳定等问题。虽然分子优化是必不可少的,但它不能完全解决营养扩散限制、剪切应力敏感性和收获期间蛋白质组破坏等挑战。这些障碍需要在生物工艺设计方面同时取得进展。
培养肉的潜在环境效益是巨大的。与传统畜牧业相比,它可以减少78%–96%的温室气体排放,减少99%的土地使用,并降低82%–96%的水资源使用[12]. 在大规模实现这些效益取决于弥合当前细胞培养生产力与经济可行性之间的差距。核糖体工程是帮助缩小这一差距的强大工具,但它必须是包括分子生物学、生物工艺创新和综合多组学监测在内的更广泛、综合方法的一部分。只有通过结合这些努力,才能实现培养肉的全部潜力。
如何Cellbase 支持培养肉研究
从分子优化到培养肉的大规模生产需要在每个阶段使用精确的工具和材料。
对于从事细胞系优化的团队,
在扩大生产规模时,
常见问题
哪种核糖体工程方法对培养肉细胞系最有前景?
培养肉的核糖体工程研究旨在增强蛋白质生物合成并影响细胞命运决策。一种有前景的方法是核糖体池工程, ,它通过修改核糖体RNA操纵子来提高翻译效率。像iSAT和RISE这样的工具提供了体外核糖体进化的平台,使得开发具有改进功能的核糖体成为可能。此外,像
如何在不引起蛋白质错误折叠或细胞压力的情况下提高翻译速率?
为了在不触发蛋白质错误折叠或细胞压力的情况下提高翻译速率,研究人员专注于微调翻译过程,而不是全面加速。一些关键方法包括:
- 使用慢翻译密码子 : 这些有助于将翻译速度与蛋白质折叠的自然过程对齐,确保正确的结构形成。
- 减少5'编码区的自由折叠能量: 这种调整可以提高蛋白质生产效率,同时保持细胞健康。
其他技术包括低诱导方案, 温度下降, 和先进的合成工具,如SINEUP RNAs. 这些策略可以在不增加细胞负担的情况下实现更高的蛋白质产量。
对于使用特殊材料的人来说,像
生物反应器需要进行哪些更改以支持超过200 µm的核糖体工程肌肉组织?
为了生长厚度超过200 µm的肌肉组织,生物反应器必须克服与营养、氧气和pH扩散相关的挑战——这些因素对于三维结构中的细胞存活至关重要。搅拌罐生物反应器需要精确调整以维持均匀条件,同时减少可能损害细胞的剪切应力。在许多情况下,灌流系统在创建稳定环境方面发挥关键作用,尤其是在密集组织中。对于使用特殊生物反应器和材料的人来说,









