

组蛋白修饰是对蛋白质的化学改变,它们在不改变DNA的情况下影响基因活性。这些修饰对于培养肉生产中使用的细胞系的开发至关重要,帮助细胞生长、保持其身份并分化为肌肉组织。文章探讨了特定组蛋白标记如H3K4me3(基因激活)、H3K27ac(增强子活性)和H3K27me3(基因抑制)如何调节细胞行为。
涵盖的关键点:
- H3K4me3 支持活跃基因和快速分化。
- H3K27ac 在生长阶段控制基因表达的增强子。
- H3K27me3 确保不需要的基因程序保持不活跃。
- 由这些标记塑造的染色质状态在不同物种和细胞类型之间有所不同,影响生产质量。
该文章还强调了最近的研究,包括猪细胞中的位置基因表达如何影响肉质,以及如何通过靶向表观遗传编辑来提高细胞系性能。未来的方向包括完善表观遗传工具和研究染色质状态,以优化生产效率和规模.
组蛋白修饰解释 | 乙酰化、甲基化 & 基因调控
sbb-itb-ffee270
组蛋白修饰的类型及其功能
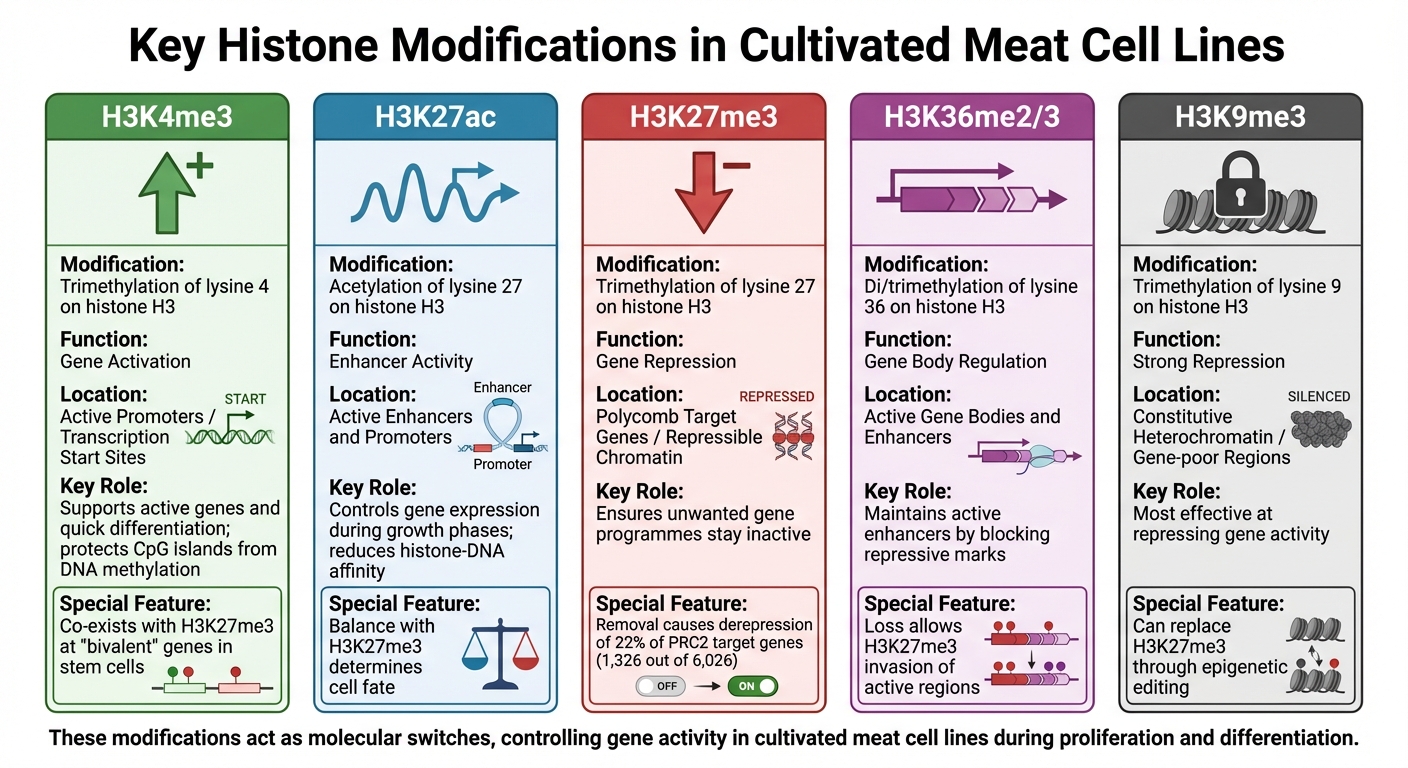
培养肉细胞系中的关键组蛋白修饰:功能和基因组背景
组蛋白修饰在调节基因活性中起着至关重要的作用,像分子开关一样控制培养肉细胞系中基因的开启或关闭。这些化学标签 - 主要是甲基化和乙酰化 - 附着在组蛋白的特定残基上,形成独特的基因组模式。每种修饰都有特定的功能,通过了解这些角色,研究人员可以更好地预测和影响生产过程中的细胞行为。这些知识对于优化培养肉生物加工.
过程至关重要。以下是影响培养肉细胞系基因调控的主要组蛋白修饰的分解。
H3K4me3和基因激活
H3K4me3(组蛋白H3上赖氨酸4的三甲基化)与活跃的基因启动子相关,并促进基因起始位点的转录,特别是涉及细胞生长和代谢的基因。这种修饰还保护CpG岛启动子免受新的DNA甲基化,确保必需基因保持可转录状态[4].
在用于培养肉的原代或永生化细胞系中,H3K4me3通常与H3K27me3等抑制性标记共存于“双价”基因。这些基因保持激活状态,能够在需要时快速分化为肌肉组织[4] .
有趣的是,H3K4me3与其他修饰相互作用。例如,H3K36me3的沉积可以抑制H3K4甲基转移酶,降低启动子处的H3K4me3水平并改变基因表达模式[4].
H3K27ac和增强子活性
H3K27ac(组蛋白H3上赖氨酸27的乙酰化)是活性增强子和启动子的标志。通过降低组蛋白与DNA之间的亲和力,H3K27ac创造了促进转录的环境[5]. 在培养肉细胞系中,不同生长阶段H3K27ac水平的变化决定了细胞从增殖到分化过程中哪些基因被表达。
H3K27ac与H3K27me3等抑制性修饰之间的平衡是决定细胞命运的关键。例如,支持增强子活性的H3K36me2的丧失可能允许H3K27me3侵入先前活跃的区域,降低H3K27ac水平并沉默目标基因[5].
H3K27me3和基因抑制
H3K27me3(组蛋白H3上赖氨酸27的三甲基化)是一种抑制性标记,促进闭合染色质结构,有效地沉默基因。这种修饰由Polycomb Repressive Complex 2(PRC2)催化,对于维持数千个发育基因的抑制至关重要[4] .
在培养肉细胞系中,H3K27me3确保在特定生长阶段不需要的基因程序保持不活跃,从而保持细胞的预期身份。
"H3K27me3与H2AK119ub1一起,对于维持数千个Polycomb靶基因的转录抑制至关重要。" - Nature Communications [4]
研究表明,去除小鼠胚胎干细胞中的H3K27me3会导致约22%(6,026个PRC2靶基因中的1,326个)的基因去抑制 [4]. 对于培养肉,控制这种修饰可以帮助抑制替代细胞命运,例如脂肪或结缔组织的形成,同时专注于肌肉发育。
| 组蛋白修饰 | 调控功能 | 基因组背景 |
|---|---|---|
| H3K4me3 | 基因激活 | 活跃启动子 / 转录起始位点 |
| H3K27ac | 增强子活性 | 活跃增强子和启动子 |
| H3K27me3 | 基因抑制 | Polycomb 目标基因 / 可抑制染色质 |
| H3K36me2/3 | 基因体调控 | 活跃基因体和增强子 |
| H3K9me3 | 强抑制 | 组成性异染色质 / 基因贫乏区域 |
培养肉细胞系中的染色质状态
组蛋白修饰不是单独起作用的 - 它们结合形成染色质状态, ,这些是控制基因可及性的独特基因组环境。这些状态在扩增和分化过程中对培养肉细胞系的行为起着至关重要的作用,使其成为优化生物加工的关键。
通过组蛋白标记识别染色质状态
研究人员通过研究组蛋白标记的组合来绘制染色质状态图,例如H3K4me3、H3K27ac和H3K27me3。例如,在猪胎儿成纤维细胞(PFF)和滋养外胚层(PTr2)细胞中,已识别出10种不同的染色质状态,包括活跃的转录起始位点、双效启动子和假定增强子[6]. 这些状态有助于预测基因活性。
增强子状态主要由H3K27ac在基因间和内含子区域标记,通常与染色质重塑蛋白BRG1共同富集 [6].
一个特别显著的特点是存在宽广的H3K4me3区域, ,这些区域跨越4 kb或更大的区域。这些区域仅占1.7%到1.8%的所有预测的转录起始位点在猪细胞系中,但对于标记发育和组织特异性基因至关重要[6]. 有趣的是,在猪胎儿成纤维细胞中,52%的基因 由这些宽广区域标记为组织特异性,而在PTr2细胞中仅为25% [6].
“这些发现增强了我们对早期猪发育中表观遗传景观的理解,并提供了关于染色质状态的变化如何与细胞身份相关的见解。" - BMC Epigenetics & 染色质 [6]
这些染色质状态概况不仅在单一物种内有所不同,而且在用于培养肉生产的各种动物细胞系中也有所变化。
动物细胞系间的染色质差异
染色质状态模式根据用于培养肉生产的物种和细胞类型的不同而显著变化。例如,在鸡细胞系中,H3K4me3在基因启动子上的基因组存在占比为 30% 到 55% [7]. 然而,在鸡的原始生殖细胞(PGCs)中,H3K4me3水平相比多能细胞显著下降。这种减少支持了在生殖系规范化过程中双价状态向抑制状态的转变 [7].
猪滋养层(PTr2)细胞在启动子区域显示出更高的H3K27ac水平(57.36%),而胎儿成纤维细胞为(41.58%),同时PTr2细胞中的H3K27me3富集较低(7.77%),而PFF细胞为(22%)[6]. 这些变化反映了每个发育阶段不同的表观遗传需求,并影响这些细胞对培养条件的响应方式。
在牛卫星细胞 , 分化为“储备细胞”命运(Pax7+/Ki-67-)是由静止染色质状态通过NOTCH和MAPK/ERK信号调节驱动的。然而,这一过程会降低蛋白质产量[3]. 这种变异性强调了染色质状态如何直接影响生产效率。深入了解这些差异对于在培养肉生产中微调细胞系性能至关重要。
利用组蛋白修饰改进细胞系
基于我们对染色质状态的了解,让我们深入探讨如何通过靶向组蛋白修饰直接提高培养肉细胞系的性能。
促进增殖和适应悬浮生长
调整组蛋白标记可以显著增加细胞增殖,并帮助细胞从贴壁生长过渡到悬浮生长。这一转变对于培养肉的生物反应器系统. 至关重要。例如,减少H3K36甲基化使成纤维细胞对TGFβ的反应性降低,从而导致更灵活的细胞状态[1].
2022年12月,Believer Meats的研究人员在鸡成纤维细胞(HUN-CF-2和HUN-CF-4)上取得了突破。他们在 无血清悬浮培养, 中展示了自发的永生化,达到每毫升1亿个细胞(10⁸个细胞/毫升),并实现了36% w/v. 的产量。由Yaakov Nahmias领导的团队使用卵磷脂——一种食品安全的小分子——激活PPARγ通路并促进脂肪形成,而不依赖于基因改造。他们培养的鸡肉原型获得了4.5分(满分5.0) 的感官评分[2].
“不依赖基因改造的永生化和高产量制造对于培养肉的市场实现至关重要。” - Yaakov Nahmias,Believer Meats首席科学官 [2]
这些发现突显了精确表观遗传工具在进一步优化细胞系开发中的潜力。
表观遗传编辑的精确性
为了补充这些细胞变化,精确的表观遗传编辑方法允许对组蛋白标记进行有针对性的操控。2025年一项关于小鼠胚胎干细胞的研究表明,融合了SUV39H2或SETD2催化域的嵌合招募器(S12N)可以在数千个基因上用H3K9me3或H3K36me3替代H3K27me3。其中,H3K9me3在抑制基因活性方面更为有效[8].
然而,这些修饰的成功在很大程度上依赖于现有的染色质环境。例如,基因启动子上的残留H3K4me3可以阻碍DNA甲基化机制,使得实现预期的基因沉默更加困难[8]. 这表明,优化细胞性能通常需要同时调整多个组蛋白标记,而不是专注于单一修饰。
结论和未来方向
关键要点
组蛋白修饰作为分子开关, 在培养肉细胞系中控制基因活性方面起着关键作用。具体来说,H3K36me2和H3K36me3通过阻止H3K27me2/3等抑制性标记渗入基因体来帮助维持活性增强子 [9][10]. 当H3K36甲基化丢失时,染色质结构被破坏,允许H3K9me3等抑制性标记侵入活性区域[9].
"H3K36甲基化[是]染色质状态和基因组结构的关键调节器。" - Nature Communications[9]
组蛋白标记之间的相互作用对于提高细胞系性能至关重要。研究表明,同时针对多种组蛋白修饰通常比仅关注一种能获得更好的结果[4].
考虑到这些发现,未来的研究必须利用精准的表观遗传工具,以确保在培养肉细胞系性能上的持续改进。
未来研究机会
提升细胞系性能需要创新的方法,例如单核RNA测序, 以绘制不同细胞亚群内的表观遗传图谱。这对于识别和理解抗拒分化的静止“储备细胞”尤为重要。这些细胞表达PAX7和NOTCH2等标记,而不是致力于肌源性融合,在培养肉生产中构成了重大挑战[3].
另一个有前景的途径是开发嵌合表观遗传复合物,以实现精确的非遗传控制。例如,在2025年,研究人员展示了将SUZ12的N端与SUV39H2或SETD2的催化结构域结合,可以有效地用H3K9me3或H3K36me3替代许多基因上的H3K27me3 [4] . 此外,监测增强子上的H3K36me2可以作为质量控制标记,以确保细胞系的稳定性 [9].
未来的努力应集中于维持细胞代际间的H3K36甲基化。这将有助于防止表观遗传漂移,使研究人员和像
常见问题
组蛋白标记如何影响培养肉细胞系的肌肉分化?
组蛋白标记是肌肉分化的关键因素,特别是对于培养肉细胞系。例如,分化过程中H3K27me3 的减少会触发肌源性转录程序,从而激活肌肉发育所需的基因。微调像H3K27me3 这样的组蛋白修饰支持细胞系从增殖向具有特定特征的肌肉组织形成的过渡。这些表观遗传调整对于推进培养肉的生产至关重要。
哪些组蛋白修饰最能预测生物反应器中稳定、高产的细胞生长?
H3K36甲基化作为生物反应器中稳定、高产细胞生长的可靠标记脱颖而出。这种修饰在保持细胞身份和管理谱系程序中起着关键作用——这两者对于确保细胞持续增殖至关重要,特别是在培养肉的生产中。
表观遗传编辑能否在不改变DNA序列的情况下改善细胞系?
表观遗传编辑提供了一种在不改变DNA序列的情况下改善细胞系的方法。通过调整组蛋白标记和染色质结构,它控制基因表达。关于组蛋白修饰的研究强调了这些改变如何影响细胞身份和功能。这种方法在改进培养肉细胞系方面具有特别的前景。










