

表观遗传沉默正在改变我们对培养肉生产的方式。对于R&D专业人士来说,它提供了一种无需永久改变DNA即可控制基因表达的方法,解决了细胞增殖、分化和质量控制等关键挑战. 以下是您需要了解的内容:
- 定义: 通过DNA甲基化、组蛋白修饰或RNA干扰抑制基因活性——可逆且精确的方法,保持遗传序列完整。
- 重要性: 延长细胞寿命,增强肌肉细胞分化,提高可扩展性,同时避免永久基因编辑带来的致癌风险。
- 关键工具: CRISPR-dCas9系统(如KRAB或DNMT3A)和基于TALE的编辑器实现高效沉默,有些效果持续超过300天。
- 挑战: 在大规模生产中交付这些工具,尤其是在生物反应器中,并针对特定物种的途径进行定制仍然是难题。
对于生物工艺工程师和细胞培养科学家来说,重点是精确控制细胞行为以提高生产力和产品质量。表观遗传沉默可能是克服培养肉生产瓶颈.
家畜细胞中表观遗传沉默的核心机制
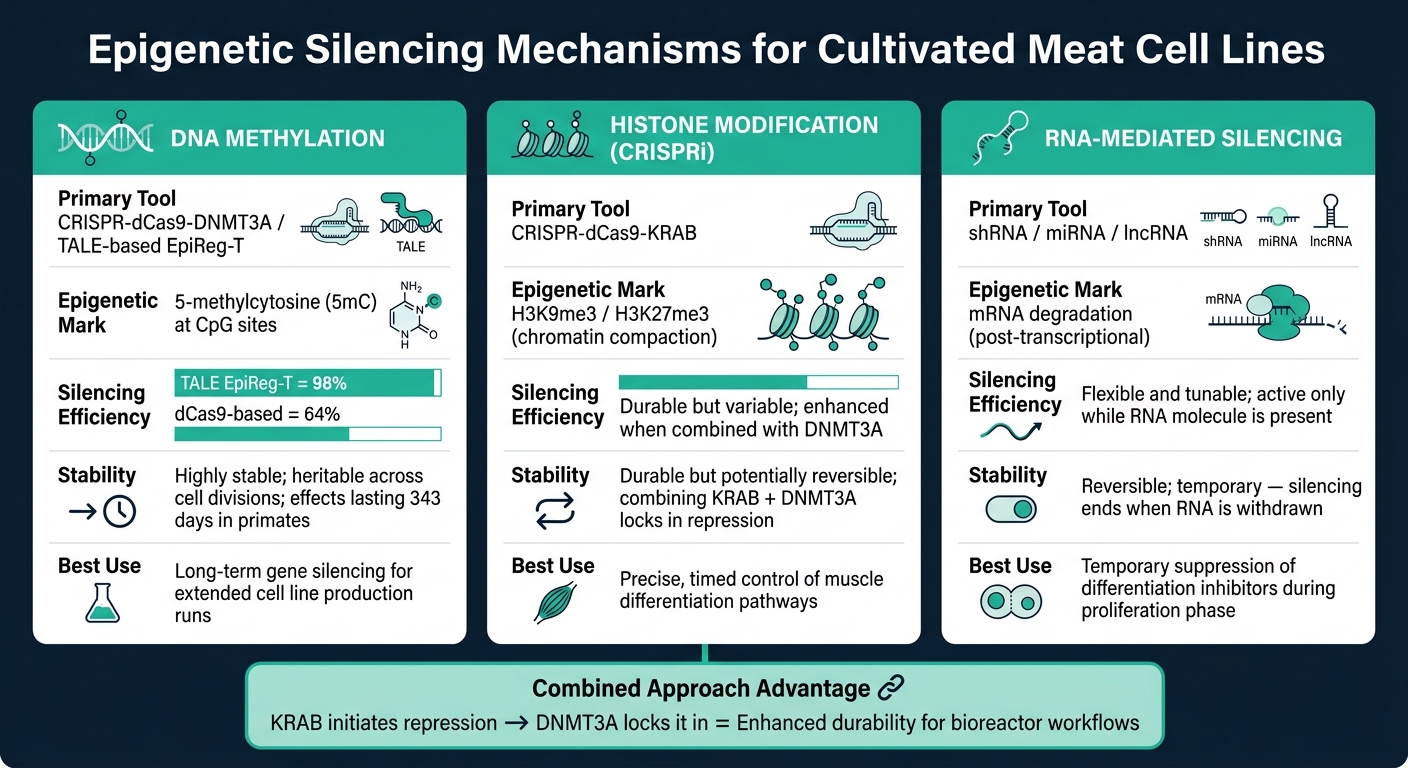
培养肉的表观遗传沉默工具:机制、效率&稳定性
提高培养肉细胞系的性能在很大程度上依赖于对表观遗传机制的精确控制。以下是家畜细胞中使用的主要方法概述。
基于DNA甲基化的沉默
DNA甲基化涉及将甲基基团添加到CpG位点,这一过程由DNA甲基转移酶(DNMTs)驱动。当这种情况发生在基因启动子区域时,它会阻止转录机制访问基因,从而有效地关闭基因[6]. 这种沉默是可遗传的,DNMT1在细胞分裂过程中维持甲基化模式[7].
一种先进的工具,CRISPR-dCas9-DNMT3A, 结合了催化失活的dCas9蛋白和DNMT3A酶,将甲基化引导至特定的基因组位置。此方法在不切割DNA的情况下实现了高沉默效率。一种更精细的方法,基于TALE的表观遗传调节器(EpiReg-T), 在小鼠中显示出98%的沉默效率,相比之下,早期的dCas9系统为64%[5] . 在非人灵长类动物的研究中,单剂量的该系统维持基因沉默长达343天 [5] .
在DNA甲基化建立之后,组蛋白修饰提供了一个次要的、动态的基因调控层。
组蛋白修饰和CRISPRi
组蛋白修饰通过靶向组蛋白改变染色质结构,使基因更易或更难接近。像H3K9me3和H3K27me3 这样的标记使染色质紧密化,阻止转录因子接触DNA [6] .
CRISPR干扰(CRISPRi)利用与KRAB抑制域融合的dCas9。该复合物被引导至特定基因启动子,在那里招募抑制蛋白质,沉积抑制性组蛋白标记。在绵羊的研究中,H3K27me3 被强调为肌肉发育过程中的关键抑制信号,而活跃的增强子与促进优越生长性能的基因相关联[8] . 通过了解调节牲畜肌肉分化的组蛋白状态,科学家可以精确地微调细胞行为。
"表观遗传编辑是一种有前途的策略,可以在避免基因组编辑技术的永久性改变和潜在基因毒性的同时修改基因表达。" - Nature Biotechnology [5]
组蛋白修饰通常比DNA甲基化更具动态性,需要持续或定时干预以维持其效果。将KRAB与DNMT3A结合在一个构建体中可以增强持久性:组蛋白标记启动抑制,而甲基化将其固定[5].
除了这些基于DNA的方法外,RNA介导的沉默提供了一种灵活且临时的替代方案。
RNA介导的沉默
RNA介导的沉默直接关注于降低mRNA水平。微小RNA(miRNA)和短发夹RNA(shRNA)与互补的mRNA序列结合,导致其在翻译前降解 [6] . 同时,长非编码RNA(lncRNA)通过将染色质修饰复合物招募到特定基因组区域来更早地发挥作用[6] .
对于培养肉应用,RNA介导的沉默提供了一个主要优势:可逆性和灵活性. 沉默仅在RNA分子存在时保持活跃,使其成为临时干预的理想选择。例如,可以在增殖阶段抑制分化抑制剂,然后移除以允许正常的肌肉发育。然而,在将细胞系扩展到生物反应器培养时,保持RNA分子的连续递送可能会增加复杂性scaling cell lines for bioreactor cultivation.
下表总结了这些机制的关键特征:
| 机制 | 主要工具 | 表观遗传标记 | 稳定性 |
|---|---|---|---|
| DNA 甲基化 | CRISPR-dCas9-DNMT3A | 5-甲基胞嘧啶 (5mC) | 高度稳定;可遗传分裂[5][7] |
| 组蛋白抑制 (CRISPRi) | CRISPR-dCas9-KRAB | H3K9me3 / H3K27me3 | 持久但可能可逆[5][8] |
| RNA 干扰 | shRNA / miRNA | mRNA 降解 | 可逆且可调节[6] |
sbb-itb-ffee270
提高细胞系性能的目标基因和通路
在之前关于表观遗传机制的讨论基础上,选择合适的基因目标对于提高细胞系性能至关重要。这些干预措施的成功不仅取决于目标的识别,还取决于选择合适的沉默方法。研究已经确定了一组核心基因目标,当这些基因被抑制时,可以增强培养肉细胞系的关键方面,包括增殖、分化和代谢稳定性。
增殖和永生化
提高增殖能力通常涉及靶向基因如CDKN2A和TP53. CDKN2A 编码p16^INK4A和p14^ARF,这些蛋白质限制细胞周期并驱动衰老。沉默CDKN2A可防止G1/S停滞,从而实现强劲的细胞扩增。例如,猪细胞 通过CDKN2A 沉默在18-30代次中保持了其肌源性特性,而野生型细胞在第10代次时就失去了这些特性。此外,CDKN2A 的耗竭在第20代时导致PAX7 表达增加约194倍,这是肌肉干细胞身份的关键因素 [9].
“靶向CDKN2A基因位点对于防止衰老或诱导细胞永生化至关重要。” - 食品材料研究[9]
TP53是另一个关键目标。在牛间充质干细胞中对600个基因进行的CRISPR筛选中,TP53被确定为增强增殖的最有效目标。敲除TP53 在30天内导致细胞丰度增加1000倍,并在长期扩展中表现一致 [1]. 此外,沉默 PTEN, PI3K/AKT/mTOR通路的负调节因子,能提高细胞倍增率和mTOR活性。然而,这种方法需要仔细监控,因为它可能会降低分化效率[1].
这些增殖方面的进展为优化分化奠定了基础,这是下一个关键步骤。
控制分化
在培养肉生产中,平衡细胞扩增与组织形成是一个复杂的挑战。一个研究较多的目标是肌生成抑制素 (MSTN), 肌生成的负调节因子。沉默MSTN可以增强肌纤维的形成,类似于某些牛品种中的“双肌”特性[4]. 当结合 MYOD1激活和先进技术如数字光处理 (DLP) 3D 生物打印在沟槽图案水凝胶上时,通过表面功能化,肌肉细胞的排列和分化显著改善 [4].
另一个关键方面是管理多能性调节因子,如SOX2和OCT4. 使用CRISPR/dCas9-KRAB平台对SOX2 进行可逆沉默,在72小时内实现高达85%的抑制,编辑构建体撤回后基线表达恢复到约90%[3]. 这种可逆性允许在细胞扩增期间进行受控抑制,并在适当的时间释放以支持正常的组织发育。
压力和代谢途径
在延长生产周期中保持细胞质量涉及解决压力和代谢挑战。TP53在肿瘤抑制和压力传感中起双重作用。在培养条件下,即使没有显著的基因组损伤,它也可能过早触发衰老[1]. 通过沉默 TP53, 细胞保留早期传代细胞的基因表达谱,保留蛋白质合成和DNA复制等关键功能[1].
下表总结了主要基因靶点及其功能角色:
| 靶基因 | 通路 | 沉默效应 | 物种背景 |
|---|---|---|---|
| CDKN2A | 细胞周期抑制 | 防止衰老;~194× PAX7 在第20代时上调 [9] | 猪 |
| TP53 | 应激反应/肿瘤抑制因子 | 30天内细胞数量增加1,000倍;长期扩增一致 [1] | 牛 |
| PTEN | PI3K/AKT/mTOR | 增加倍增率;增强mTOR活性 [1] | 牛 |
| MSTN | 肌生成调节 | 增强肌纤维形成和分化效率[4] | 牛 |
| SOX2 | 多能性维持 | 管理干性到分化的过渡;72小时内抑制85%[3] | 多重 |
一种有前景的方法正在获得关注,即多重靶向技术,涉及同时沉默多个基因。例如,结合CDKN2A抑制与GATA4激活显示出协同效应,优于单独干预措施[9] [10] . 这种系统级策略强调了像
表观遗传工具和传递方法
为了利用特定的基因目标,研究人员依赖于专业的表观遗传工具和高效的传递系统。
合成表观遗传平台
确定正确的基因目标只是方程的一部分——用于沉默这些基因的工具同样关键。两个可编程系统因其与培养肉研究的相关性而脱颖而出:CRISPRoff和基于TALE的表观遗传调节器(EpiReg-T).
CRISPRoff 利用 dCas9 支架结合 KRAB 和 DNMT3A/3L 结构域来建立可遗传的抑制标记,例如 DNA 甲基化和 H3K9me3,而不引入 DNA 断裂。这种方法确保了持久的基因沉默,使其特别适用于长期维持细胞系——这是解决培养肉生产中可扩展性和一致性挑战的关键因素。相比之下,基于 TALE 的 EpiReg-T 显示出更高的沉默效率,达到 98%,而类似的 dCas9 系统仅为 64% [5] .
一项关键研究发表在 Nature Biotechnology 于 2025 年 10 月,强调了基于 TALE 的编辑器的潜力。研究人员,包括来自Epigenic Therapeutics和中国科学院, 的研究人员,展示了一剂通过脂质纳米颗粒(LNPs)递送的EpiReg-T在猕猴中以超过90%的效率沉默了PCSK9基因,持续时间为 343天. 通过多组学分析确认,这一成果是在最小化脱靶效应的情况下实现的 [5]. 这样的结果在耐久性和效力至关重要时,使TALE系统脱颖而出。
递送挑战
将这些工具有效地递送到牲畜细胞中——尤其是在大规模情况下——仍然是一个主要的技术挑战。虽然表观遗传编辑器避免了双链DNA断裂的风险,但它们仍然需要可靠的递送机制。脂质纳米颗粒(LNPs)已成为领先的非病毒选择。他们暂时性地传递编码表观遗传编辑器的mRNA,实现了一种“打击后撤”的方法,在不进行DNA整合的情况下建立持久的基因沉默[5]. 这种暂时性对于培养肉尤其重要,因为围绕基因改造的监管问题仍然是一个关键问题。
然而,LNP的效率可能会因细胞类型而显著变化。针对初级牛 或猪肌卫星细胞的配方优化,特别是在生物反应器规模的环境中,仍然是一个活跃的研究领域。在小规模实验中效果良好的传递方法往往在搅拌罐式生物反应器. 中无法一致地表现。解决这些传递挑战对于推进研究和扩大生产规模至关重要,这一过程越来越多地得到专业平台的支持。
如何 Cellbase 支持表观遗传学R&D
表观遗传修饰的细胞系需要经过精确验证的试剂。研究人员需要获得与表观遗传修饰兼容的良好表征细胞系、保持表观遗传稳定性的定义培养基配方,以及能够在染色质水平确认基因沉默的分析工具。一般实验室供应商通常缺乏确保与培养肉应用兼容的专业知识。
表观遗传沉默对培养肉类生物加工的意义
可测量的细胞系改进
表观遗传沉默提供了实际优势,这些优势越来越明显,特别是在延长细胞系的生产寿命方面。通过采用瞬时的“打击-逃跑”策略,研究人员可以暂时抑制负责衰老的基因,而不永久修改基因组 [2]. 这种方法在牛和猪肌卫星细胞, 中显示出成功,能够显著增加细胞倍增次数并解决常见的生物加工瓶颈。 重要的是,这种方法是可逆的——一旦构建体被撤回,基因表达几乎恢复到基线水平[3]. 这种可逆控制非常适合生物反应器工作流程,因为它确保细胞在扩增阶段继续增殖,并允许在适当的时间触发分化。增强的细胞扩增直接转化为更高效的组织分化和改进的产品质量。
组织形成和产品质量
细胞增殖的提高为更好的组织形成奠定了基础。受控分化是表观遗传沉默直接影响最终产品质量的地方。例如,在牛细胞重编程中,沉默多能性标记如OCT4, SOX2, 和NANOG促进了向肌源性谱系的转变。该过程导致在分化协议的第30天形成细长的、多核的肌管。[11].
"mOSKM和多能性标志物的沉默...对于从多能性向肌源性谱系的过渡至关重要。" - Frontiers in Nutrition [11]
除了肌纤维发育之外,对脂肪细胞分化途径的精确表观遗传控制在实现大理石花纹方面起着关键作用。大理石花纹是影响风味和口感的关键因素,这些改进可以在不对基因组进行永久性改变的情况下实现。
监管和消费者考量
细胞增殖和组织形成的进步也使监管和消费者的观点成为关注的焦点。监管机构普遍支持表观遗传沉默,因为它对基因组的影响是非永久性的。像dCas9-KRAB和基于TALE的EpiReg-T这样的工具避免了与双链DNA断裂相关的风险,使其适用于必须在整个生产过程中展示遗传稳定性的食品级细胞系[5].
然而,保持无转基因状态仍然是一个挑战。2025年5月,来自圣保罗大学和哥本哈根大学, 的研究人员,包括Kaiana Recchia和Kristine Freude,发表了一项研究,探讨了这个问题。他们使用非整合的环状载体对牛胎儿成纤维细胞进行了重编程,发现虽然菌落在超过33次传代后仍然稳定,但在第12和第17次传代时仍能检测到环状质粒[11].
从消费者的角度来看,关于所用方法的透明度至关重要。 清晰地传达表观遗传沉默不会永久改变DNA,这对于在培育肉产品接近商业化时建立公众信任至关重要。
未来方向和研究空白
物种特定的挑战
该领域最大的障碍之一是对牲畜物种肌生成途径缺乏详细了解。虽然像IGF-1、MAPK/Erk和Wnt/β-catenin这样的途径在人类和小鼠中有详细记录,但它们在牛和猪中的作用仅部分被理解[11]. 在没有完整图谱的情况下,定位表观遗传沉默的特定基因目标成为一个重大挑战。
肌肉纤维组成增加了复杂性。例如,猪Longissimus肌肉包含约55%的IIb型快缩纤维,但这些纤维在绵羊和马等物种中不存在。当你将其与特定区域的HOX基因表达结合起来时,很明显,沉默策略需要针对每个物种进行定制[13]. 卫星细胞保留位置性HOX基因表达( e.g. ,HOXA11 和HOXA13在后肢肌肉中),进一步复杂化了问题。这些模式可以影响细胞是更倾向于快速增殖还是强健分化[14] .
“因为SCs可以保留这些位置特征,它们的增殖和分化能力可能因肌肉来源而异。” - npj Science of Food [14]
在实际操作中,这意味着研究人员在应用表观遗传沉默之前应筛选细胞系的HOX基因表达。这些基因特征可以作为生物条形码,帮助验证细胞的区域身份,并使其与最终产品的期望特性对齐。
这种特定物种的挑战突显了在细胞库策略开发中考虑替代细胞来源(如iPSCs)的重要性。
iPSC开发和细胞库的链接
诱导多能干细胞(iPSCs) 是卫星细胞的有前途的替代品,卫星细胞容易衰老并需要反复活检。2025年5月,圣保罗大学和哥本哈根大学的研究人员——包括Kaiana Recchia和Kristine Freude——成功使用非整合的环状载体开发了牛iPSC系。这些细胞在超过33次传代中保持稳定,并在第30天分化为多核肌管[11]. 然而,通过严格的基因组PCR确认其无转基因状态仍然是一个关键步骤。
一个相关的问题是表观遗传记忆. 诱导多能干细胞(iPSCs)通常保留其原始体细胞组织的痕迹,这可能导致分化偏离预期的谱系 [12]. 对于细胞库,选择表观遗传特征已经倾向于肌肉或脂肪形成的供体组织至关重要。此外,确保有效沉默残余的多能性标记对于创建可靠的长期细胞库至关重要。
强大的iPSC协议的发展也强调了在研究工作中需要标准化的检测和一致的数据共享实践。
标准化和数据缺失
为了充分利用表观遗传干预在培养肉中的潜力,必须解决标准化问题。目前,还没有一个通用的框架来监测工业规模生产所需的大量细胞倍增过程中的表观遗传稳定性[12]. 由于缺乏标准化的方法,跨实验室比较结果变得困难,关于扩大生产规模的决策往往依赖于不完整的数据。
一些实际步骤可以帮助解决这一差距。例如,采用一致的FACS纯化协议——针对CD31⁻/CD45⁻/CD29⁺/CD56⁺等标记——将使卫星细胞群体在不同物种和解剖来源之间更具可比性[14]. 从基于血清的培养基切换到化学定义的培养基也可以减少批次之间的变异性,创造更一致的表观遗传环境[12].
展望未来,整合AI驱动的计算机模拟可能会彻底改变表观遗传协议的优化。然而,要使这些模型有效,协调培养肉研究社区的数据是至关重要的。标准化的数据共享实践将使研究人员能够更准确地预测表观遗传操作的结果,从而加速该领域的进展。
常见问题
表观遗传沉默与培养肉细胞中的永久基因编辑有何不同?
表观遗传沉默调节基因活性而不对DNA序列进行永久性改变,这与基因编辑不同,后者涉及物理改变基因组。由于表观遗传方法不涉及打断或修改DNA,因此通常被视为培养肉生产中更安全的选择。诸如CRISPR工具等技术提供了灵活的基因调控优势,并且在某些情况下是可逆的。对于使用这些方法的研究人员,
应该首先沉默哪些基因以在不损害分化的情况下促进增殖?
为了在保持细胞分化能力的同时促进细胞增殖,关键是要沉默那些阻碍细胞周期或导致不良细胞命运的基因。例如,抑制 CDKN2A 已被证明可以显著增加猪卫星细胞的增殖,而不影响其分化潜力。同样,靶向肿瘤抑制基因如 TP53 和 PTEN 可以增强生长,尽管这些干预措施需要仔细监督。
如何在生物反应器规模上可靠地递送表观遗传编辑器?
在大规模培养肉生产中递送表观遗传编辑器是一个重大挑战。这主要是由于CRISPR工具的体积庞大以及传统递送方法如电穿孔或病毒载体的限制。然而,一些有前景的策略正在出现。例如,使用脂质纳米颗粒或工程化病毒样颗粒的瞬时递送系统显示出潜力。这些方法可以封装大型CRISPR货物,允许其高效进入细胞而不引起基因组整合。为了支持这些先进的计划,









