

หากคุณสามารถขยายเซลล์ได้แต่ไม่สามารถเปลี่ยนให้เป็นชะตากรรมที่ถูกต้องในเวลาที่เหมาะสม กระบวนการของคุณจะหยุดชะงักที่การแยกแยะ นี่คือจุดสำคัญ: วงจรยีนสังเคราะห์ให้คุณควบคุมภายในเซลล์ เกี่ยวกับการตัดสินใจ, เวลา, หน่วยความจำ และการผสมผสานของสายพันธุ์ ซึ่งการเปลี่ยนแปลงของสื่อเพียงอย่างเดียวมักจะทิ้งประชากรที่มีความหลากหลาย, และมีการตัดสินใจบางส่วน
หากฉันกำลังสร้างกระบวนการแยกแยะเนื้อสัตว์ที่เพาะเลี้ยง ฉันจะนำสี่จุดจากบทความนี้ไปใช้ทันที:
- เริ่มต้นด้วยเครือข่ายพื้นเมือง ไม่ใช่โครงสร้าง ใช้การวิเคราะห์เส้นทาง snRNA-seq, การอนุมาน GRN และการวิเคราะห์โปรไฟล์ miRNA เพื่อค้นหาว่าเซลล์หยุดชะงัก, ล่องลอย หรือแยกออกไปในชะตากรรมที่ผิดพลาดที่ใด
-
จับคู่ประเภทวงจรกับปัญหากระบวนการ
สวิตช์สลับเหมาะสำหรับการล็อคอิน, การออกแบบ ฟีดฟอร์เวิร์ดหรือแบนด์พาสเหมาะสำหรับการควบคุมเวลา, เกตลอจิกเหมาะสำหรับการเกตสัญญาณหลายสัญญาณ, และ miSFITsเหมาะสำหรับเอาต์พุตที่มีการไล่ระดับ. - ออกแบบเพื่อการรั่วไหลต่ำ, เสียงรบกวนต่ำ และความปลอดภัยตั้งแต่วันแรก. ชิ้นส่วนออร์โธโกนอล, การควบคุมอัตโนมัติเชิงลบ, iFFLs, cm transgenes และโมดูลการฆ่าหรือหยุดการเจริญเติบโตที่สามารถกระตุ้นได้เป็นส่วนหนึ่งของการสร้าง, ไม่ใช่การคิดภายหลัง.
- ตรวจสอบภายใต้เงื่อนไขที่เกี่ยวข้องกับขนาดตั้งแต่เนิ่นๆ. วงจรที่ทำงานใน 2D อาจเปลี่ยนแปลงใน 3D, ไมโครแคเรียร์ หรือการกวนแขวนลอย เนื่องจากการไล่ระดับของตัวกระตุ้น, ขีดจำกัดของออกซิเจน และแรงเฉือน.
บทความยังชี้ให้เห็นถึงประเด็นที่สำคัญสำหรับทีมกระบวนการ: การควบคุมสายพันธุ์เดียวและการควบคุมอัตราส่วนเป็นงานที่แตกต่างกัน. A Tet-On MyoD cassette อาจผลักดันการเข้าสู่กล้ามเนื้อ แต่ผลิตภัณฑ์แบบ whole-cut จำเป็นต้องควบคุม สัดส่วนของกล้ามเนื้อ ไขมัน และ ECM, ซึ่งมักหมายถึงการป้อนกลับ การส่งสัญญาณแบบพาราคริน และการคัดกรองโคลนที่เข้มข้นขึ้น
ตัวเลขบางส่วนที่ยึดข้อความนั้น:
- การแยกแยะกล้ามเนื้อมาตรฐานสามารถหยุดชะงักได้ที่ ดัชนีการหลอมรวมประมาณ 50–60%
- GRN ที่ออกแบบใน iPSCs เพิ่มการแยกแยะสายพันธุ์เป้าหมายจาก 52% เป็น 81%
- วงจรสังเคราะห์ใน MSCs ที่ดัดแปลงขับเคลื่อนการแยกแยะหัวใจไปที่ 76%
- บางสายพันธุ์ Tet-On-PAX7 ของหมูยังคงมีศักยภาพในการสร้างกล้ามเนื้อสูงเกิน 40 ครั้ง
- ประมาณ 20% ของเซลล์ต้นกำเนิดพลูริโพเทนท์ของมนุษย์อาจมีการกลายพันธุ์ที่เชื่อมโยงกับมะเร็ง ซึ่งเป็นเหตุผลที่โมดูลความปลอดภัยที่สามารถเหนี่ยวนำได้มีความสำคัญ
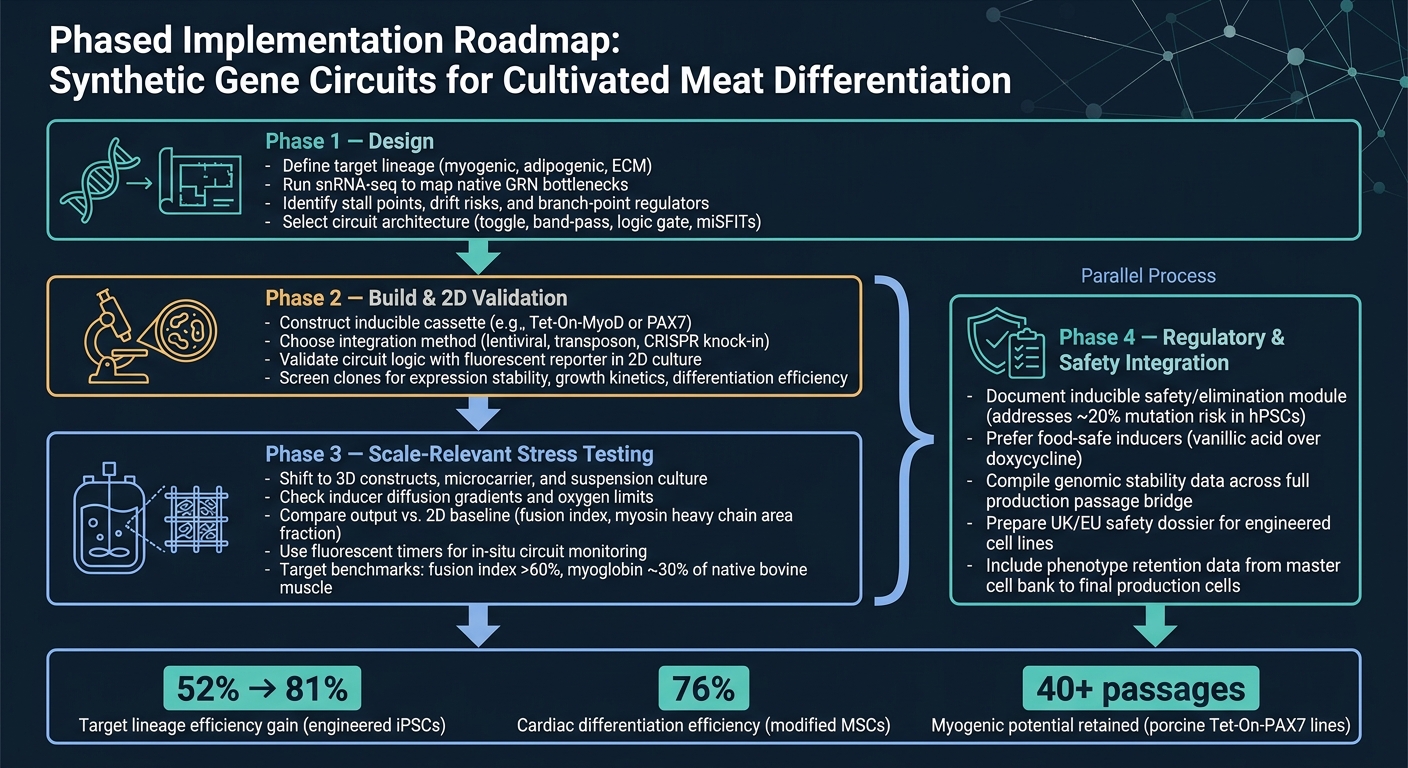
เวิร์กโฟลว์วงจรยีนสังเคราะห์สำหรับการแยกแยะเนื้อสัตว์ที่เพาะเลี้ยง
เรื่องราวของนักวิทยาศาสตร์: Michael Elowitz, วงจรพันธุกรรมในเซลล์ที่มีชีวิต
การเปรียบเทียบอย่างรวดเร็ว
| หัวข้อ | สิ่งที่บทความกล่าวในแง่ที่เข้าใจง่าย |
|---|---|
| คอขวดหลัก | การสร้างความแตกต่าง ไม่ใช่การขยายตัว |
| ทำไมวงจรถึงช่วยได้ | พวกมันเพิ่มการควบคุมเกณฑ์และหน่วยความจำสถานะเซลล์ภายในเซลล์ |
| ขั้นตอนแรกที่ดีที่สุด | ทำแผนที่การควบคุมชะตากรรมพื้นเมืองด้วย snRNA-seq และเครื่องมือที่เกี่ยวข้อง |
| ตัวเลือกวงจรหลัก | Toggle, feedforward, band-pass, logic gates, miSFITs, CRISPRa/i |
| ตัวอย่างการสร้างกล้ามเนื้อ | Tet-On-MyoD เพื่อแยกการเจริญเติบโตจากการสร้างความแตกต่างขั้นสุดท้าย |
| การสร้างเซลล์ไขมัน / การควบคุม ECM | miSFITs และการออกแบบแบบโบว์ไทสำหรับผลลัพธ์ประเภท PPARG/BMP4 แบบเกรด |
| ความท้าทายในการตัดทั้งหมด | การควบคุมอัตราส่วนระหว่างกล้ามเนื้อ ไขมัน และเนื้อเยื่อเกี่ยวพัน |
| ความเสี่ยงในการขยายขนาด | พฤติกรรม 2D อาจไม่เป็นจริงใน 3D หรือ bioreactors |
| ตัวเลือกการบูรณาการ | Lentivirus, transposons, CRISPR knock-in, episomal vectors |
| จุดควบคุมทางกฎระเบียบ | สายพันธุ์ที่ถูกออกแบบต้องมีแพ็คเกจความปลอดภัยที่กว้างขึ้น; ตัวกระตุ้นที่ปลอดภัยต่ออาหารเช่น vanillic acid เป็นที่ต้องการมากกว่า DOX เมื่อเป็นไปได้ |
ดังนั้นในคำง่ายๆ: นี่ไม่ใช่แค่บทความการออกแบบวงจร. ฉันจะอ่านมันเป็นคู่มือในการเชื่อมโยงสถาปัตยกรรมวงจร, ชีววิทยาสายพันธุ์, การเลือกโคลน, ประสิทธิภาพของเครื่องปฏิกรณ์ชีวภาพ และเอกสารความปลอดภัยของสหราชอาณาจักร/สหภาพยุโรปเข้าเป็นกลยุทธ์การสร้างความแตกต่างหนึ่งเดียว.
อ่านต่อหากคุณต้องการเส้นทางเต็มรูปแบบจากการทำแผนที่เครือข่ายพื้นเมืองไปจนถึงการเลือกโครงสร้าง, การตรวจสอบการขยายขนาด และความเหมาะสมด้านกฎระเบียบ.
2. หลักการออกแบบสำหรับวงจรควบคุมการสร้างความแตกต่าง
2.1 ทำแผนที่เครือข่ายชะตากรรมเซลล์พื้นเมืองก่อนออกแบบวงจร
ก่อนที่คุณจะออกแบบวงจร คุณจำเป็นต้องมีภาพที่ชัดเจนว่าเซลล์กำลังทำอะไรอยู่.
การจัดลำดับ RNA ของนิวเคลียสเดี่ยว (snRNA-seq) เป็นจุดเริ่มต้นที่ดี มันสามารถแสดงกลุ่มย่อยที่อยู่ในสภาวะสงบ รวมถึงเซลล์สำรองที่ถูกทำเครื่องหมายโดย NOTCH2 และ HEYL , และชี้ไปยังเป้าหมายของเส้นทางที่อาจปรับปรุงการสร้างความแตกต่าง [3].
จากนั้น การวิเคราะห์วิถีและการอนุมานเครือข่ายการควบคุมยีน (GRN) ช่วยในการทำแผนที่ลำดับการเปิดใช้งานของตัวควบคุมและเน้นจุดที่เซลล์มีแนวโน้มที่จะหยุดชะงัก ในการสร้างกล้ามเนื้อ กระบวนการหลักจะผ่าน MYOD1 และ MYOG. ในกระบวนการสร้างเซลล์ไขมัน โหนดหลักคือ PPARG และ CEBPA, โดยมีชะตากรรมของเซลล์ต้นกำเนิดไฟโบร-อะดิโพเจนิก (FAP) เป็นจุดเสี่ยงหลัก ตารางด้านล่างสรุปตัวควบคุมหลักและคอขวด
| ลำดับวงศ์ตระกูล | ตัวควบคุมหลักที่สำคัญ | เส้นทางการส่งสัญญาณที่สำคัญ | คอขวดที่ระบุได้ |
|---|---|---|---|
| กล้ามเนื้อ | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | การสร้างเซลล์สำรอง (การพักตัว) |
| การสร้างไขมัน | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | ชะตากรรมของเซลล์ต้นกำเนิดไฟโบร-อะดิโพเจนิก (FAP) |
| เซลล์ต้นกำเนิดที่มีความสามารถหลายด้าน | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | การแยกตัวเองออกจากกัน / ความหลากหลาย |
อีกชั้นที่มีประโยชน์คือ การแสดงออกของโปรไฟล์ miRNA. miRNA ภายในเช่น miR-302a, ที่เชื่อมโยงกับความสามารถหลายด้าน และ miR-375, ที่เชื่อมโยงกับการแยกตัว สามารถทำหน้าที่เป็นตัวจำแนกภายในในการออกแบบการรับรู้และตอบสนอง ที่ช่วยให้วงจรอ่านสถานะจริงของเซลล์แทนที่จะพึ่งพาเพียงตัวกระตุ้นภายนอก [5].
คอขวดเหล่านั้นควรกำหนดการเลือกวงจร หากปัญหาหลักคือการลอย คุณอาจต้องการล็อคอิน หากปัญหาคือเวลา พัลส์อาจเหมาะสมกว่า หากการควบคุมชะตากรรมขึ้นอยู่กับมากกว่าหนึ่งสัญญาณ ลอจิกหลายอินพุตมักจะมีเหตุผลมากกว่า
2.2 เลือกสถาปัตยกรรมวงจรที่เหมาะสม
นี่คือจุดที่การแลกเปลี่ยนปรากฏให้เห็นชัดเจน สถาปัตยกรรมที่เหมาะสมขึ้นอยู่กับสามประเด็นปฏิบัติ: ความถาวรของการผูกพันต้องเป็นอย่างไร การควบคุมเวลาต้องแน่นแค่ไหน และเซลล์สามารถบรรทุกพันธุกรรมได้มากเพียงใดโดยไม่มีปัญหา
สวิตช์สลับสองสถานะ เหมาะกับกรณีที่การผูกพันตามสายพันธุ์ต้องคงที่ ปัญหาหลักคือการพลิกสถานะเองที่เกิดจากเสียงรบกวนในการถอดรหัส
ฟิลเตอร์แบนด์พาส เหมาะสำหรับกรณีที่ต้องการปัจจัยการถอดรหัสเฉพาะในช่วงเวลาพัฒนาที่กำหนด ปัญหาคือระดับของตัวกระตุ้นต้องการการควบคุมอย่างเข้มงวด มิฉะนั้นเวลาจะคลาดเคลื่อน
เกตลอจิก เพิ่มความเฉพาะเจาะจงโดยต้องการมากกว่าหนึ่งอินพุตในเวลาเดียวกัน ตัวอย่างเช่น การแยกแยะสามารถอนุญาตได้เฉพาะเมื่อมีตัวกระตุ้นภายนอก และ เซลล์แสดงโปรไฟล์ miRNA ภายในที่ถูกต้อง ซึ่งช่วยลดความเสี่ยงของการผูกมัดที่ไม่ตรงเป้าหมาย
ตารางด้านล่างสรุปสถาปัตยกรรมหลักและการแลกเปลี่ยนที่มาพร้อมกับพวกมัน
| สถาปัตยกรรม | การย้อนกลับได้ | ความแม่นยำเชิงเวลา | ความซับซ้อนในการบูรณาการ | กรณีการใช้งานหลัก | ความเสี่ยงหลัก |
|---|---|---|---|---|---|
| สวิตช์สองสถานะ | ต่ำ (เมื่อถูกล็อคแล้ว) | ปานกลาง | ปานกลาง | ความมุ่งมั่นในสายพันธุ์ถาวร | การพลิกกลับเองเนื่องจากเสียงรบกวน |
| ตัวกรองแถบผ่าน | สูง (ขึ้นอยู่กับความเข้มข้น) | สูง | สูง | ขั้นตอนการพัฒนาชั่วคราว | ต้องการการควบคุมตัวกระตุ้นอย่างเข้มงวด |
| เกตลอจิก (AND/OR/NOT) | แปรผัน | ปานกลาง | ปานกลาง–สูง | การกระตุ้นเฉพาะชนิดของเซลล์ | การรั่วไหลในสถานะปิด |
| โบว์ไท / อินพุตหลายรายการ | สูง | ปานกลาง | ปานกลาง | การรวมสัญญาณหลายรายการ | พึ่งพาเสถียรภาพของ miRNA ภายใน |
| miSFITs | สูง | ปานกลาง | ต่ำ–ปานกลาง | การปรับจูนเอาต์พุตแบบเกรด | ช่วงไดนามิกแคบถ้าปรับจูนไม่ดี |
"โดยการลดจำนวนชั้นการคำนวณในขณะที่ยังคงรักษาฟังก์ชันการทำงาน กลยุทธ์นี้แก้ไขอุปสรรคด้านการขยายตัวในวิศวกรรมวงจรยีน" - Nature Communications [9]
แต่ละชั้นการควบคุมที่เพิ่มขึ้นจะเพิ่มภาระและใช้ทรัพยากรของเซลล์ ในทางปฏิบัติ หากการออกแบบสองแบบทำงานเหมือนกัน แบบที่ง่ายกว่ามักจะเป็นตัวเลือกที่ดีกว่าเมื่อขนาดมีความสำคัญ
เมื่อกำหนดสถาปัตยกรรมแล้ว งานต่อไปคือทำให้มันคงทนภายใต้การรั่วไหลต่ำ การระงับเสียงรบกวน และการควบคุมที่ปลอดภัย
2.3 สร้างเพื่อความน่าเชื่อถือ การรั่วไหลต่ำ และความปลอดภัย
วงจรต้องคงที่ในระยะเวลาการเพาะเลี้ยงที่ยาวนาน การทำงานที่ดีในช่วงสั้น ๆ ไม่เพียงพอสำหรับการใช้งานในการผลิต
ชิ้นส่วนออร์โธโกนัล เป็นการป้องกันแรก โปรโมเตอร์ ปัจจัยการถอดรหัส และองค์ประกอบการควบคุมที่ไม่ทำปฏิกิริยาข้ามกับเครื่องจักรพื้นเมืองช่วยจำกัดผลกระทบที่ไม่ตรงเป้าหมายและลดโอกาสที่สัญญาณภายในจะเปิดหรือปิดวงจรโปรโมเตอร์ที่มีการปรับเปลี่ยนให้มีความแน่นสูง เช่น PCREm ถูกใช้เพื่อลดการรั่วไหลพื้นฐานในระบบสัตว์เลี้ยงลูกด้วยนมที่สามารถเหนี่ยวนำได้ [6] .
การควบคุมตนเองเชิงลบ ก็ควรเพิ่มเข้าไปเมื่อเป็นไปได้ มันเป็นหนึ่งในรูปแบบที่รู้จักกันดีในการลดเสียงรบกวนของการถอดรหัสและสร้างการตอบสนองที่เป็นเส้นตรงมากขึ้นต่อความเข้มข้นของสารเหนี่ยวนำ [6]. วงจรป้อนกลับไปข้างหน้าแบบไม่สอดคล้อง (iFFLs) สามารถเพิ่มชั้นของการควบคุมอีกชั้นหนึ่งโดยการกรองความผันผวนแบบสุ่ม เพื่อให้เซลล์ตอบสนองต่อสัญญาณที่ยั่งยืนแทนที่จะเป็นเสียงรบกวนสั้น ๆ
เวอร์ชันที่มีการปรับเปลี่ยนโคดอน (cm) ของปัจจัยการถอดรหัสสังเคราะห์ทำให้การจำแนกง่ายขึ้นเช่นกัน พวกเขาให้คุณแยกการแสดงออกที่ขับเคลื่อนด้วยวงจรออกจากการแสดงออกของจีโนมภายใน ( g) ระหว่างการตรวจสอบ [1]. นั่นอาจฟังดูเป็นรายละเอียดเล็กน้อย แต่ช่วยประหยัดเวลาเมื่อคุณพยายามหาว่าการอ่านค่ามาจากวงจรหรือจีโนมของโฮสต์
จำเป็นต้องมีโมดูลความปลอดภัย ประมาณ 20% ของเซลล์ต้นกำเนิดพลูริโพเทนต์ของมนุษย์มีการกลายพันธุ์ที่เกี่ยวข้องกับมะเร็ง [7]. ดังนั้น หากวงจรกำลังเข้าสู่สายเซลล์ที่ได้จากเซลล์ต้นกำเนิด ควรรวมโมดูลการหยุดการเจริญเติบโตหรือการกำจัดที่สามารถเหนี่ยวนำได้ กรดวานิลลิก เป็นตัวเหนี่ยวนำที่มีประโยชน์ที่ควรให้ความสำคัญที่นี่เพราะเป็นสารเติมแต่งอาหารที่ได้รับอนุญาต ซึ่งช่วยสนับสนุนการใช้เป็นตัวกระตุ้นวงจรในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง [1].
"ชีววิทยาสังเคราะห์ให้วิศวกรมีวิธีการในการปรับใช้วงจรเพื่อปรับการแสดงออกของยีนหลายตัวได้อย่างง่ายดายและแม่นยำเพื่อ... กำจัดผลข้างเคียงที่ไม่พึงประสงค์ที่อาจเกิดขึ้นนอกเป้าหมาย" - npj Systems Biology and Applications [6]
ตัวเลือกเหล่านี้ตั้งค่าระบบวงจรเฉพาะสายพันธุ์ในส่วนที่ 3.
sbb-itb-ffee270
3. กลยุทธ์วงจรสำหรับการแยกแยะ myogenic, adipogenic และการควบคุมอัตราส่วน
3.1 วงจร myogenic ที่แยกการขยายตัวออกจากการแยกแยะขั้นสุดท้าย
เมื่อสถาปัตยกรรมวงจรถูกตั้งค่าแล้ว งานต่อไปคือการใช้งานเฉพาะสายพันธุ์ สำหรับ myogenesis ปัญหาหลักคือการระบุได้ง่ายแต่ยากที่จะดำเนินการ: เซลล์จำเป็นต้องขยายตัวก่อน จากนั้นจึงเปลี่ยนไปสู่การแยกแยะเมื่อถูกขอ โดยไม่ลอยไปในทิศทางใดทิศทางหนึ่งเร็วเกินไป.
A Tet-On-MyoD cassette เป็นหนึ่งในวิธีที่ตรงที่สุดในการทำเช่นนี้ ในการตั้งค่านี้ เซลล์จะขยายตัวภายใต้สภาวะมาตรฐานเมื่อไม่มี doxycycline (DOX) เพิ่ม DOX และวงจรจะขับเคลื่อนการมุ่งมั่นของ myogenic.นักวิจัยจากหลายสถาบันในจีนใช้วิธีนี้ในเซลล์ไฟโบรบลาสต์ของตัวอ่อนไก่และรายงานการสร้างไมโอทูบที่มีประสิทธิภาพหลังจากการเหนี่ยวนำ [4].
สวิตช์สลับสองสถานะ ให้การควบคุมสถานะที่แน่นหนาขึ้น ระบบที่สร้างจากตัวยับยั้งที่ยับยั้งซึ่งกันและกัน เช่น E-KRAB และ Pip-KRAB สามารถเปลี่ยนพัลส์ DOX สั้น ๆ ให้เป็นโปรแกรมไมโอเจนิกที่เสถียร [6]. ในทางปฏิบัติ หมายความว่าสถานะแยกแยะยังคงล็อคอยู่หลังจากการเหนี่ยวนำแทนที่จะจางหายไปเมื่อสัญญาณถูกลบออก การเพิ่มการควบคุมอัตโนมัติเชิงลบยังช่วยลดเสียงรบกวนแบบสุ่มและจำกัดการแยกแยะที่รั่วไหลในช่วงการขยายตัว [6] .
สิ่งนี้มีความสำคัญเนื่องจากการแยกแยะไมโอเจนิกในหลอดทดลองมาตรฐานมักจะหยุดชะงักกลางคัน ดัชนีการหลอมรวมมักจะอยู่ที่ประมาณ 50–60% ซึ่งทำให้มีประชากรสำรองที่ไม่หลอมรวมจำนวนมาก [3]. การกระตุ้น MyoD ที่ขับเคลื่อนด้วยวงจรสามารถปรับปรุงความมุ่งมั่นได้ แต่ความมุ่งมั่นเพียงอย่างเดียวไม่รับประกันการหลอมรวมที่สม่ำเสมอ เมื่อวงจร MyoD รวมกับการปรับ MEK, NOTCH และ RXR การหลอมรวมเกือบสมบูรณ์ในวัฒนธรรม 2D จะเป็นไปได้ [3]. วงจรควบคุมเวลาของการเข้าสู่สายพันธุ์; โมเลกุลขนาดเล็กผลักดันการออกจากวงจรเซลล์ที่สะอาดและสม่ำเสมอยิ่งขึ้น.
3.2 การควบคุมการสร้างไขมันและไฟโบรเจนิกสำหรับองค์ประกอบและโครงสร้าง
กล้ามเนื้อเพียงอย่างเดียวไม่เพียงพอ เมื่อการควบคุม myogenic อยู่ในสถานที่ ปัญหาต่อไปคือองค์ประกอบ: ไขมันจะก่อตัวมากแค่ไหน, ECM จะถูกสะสมมากแค่ไหน, และเมื่อโปรแกรมเหล่านั้นเปิดใช้งาน ที่นี่ การควบคุมแบบเปิด/ปิดมักจะหยาบเกินไป สิ่งที่ทีมมักต้องการคือผลลัพธ์ที่เป็นขั้นตอน โดยเฉพาะอย่างยิ่งรอบจุดแยก adipogenic-fibrogenic.
miSFITs ให้วิธีการที่ใช้งานได้จริงในการปรับการแสดงออกเป็นขั้นตอน.โดยการวางตำแหน่งไซต์เป้าหมาย miRNA ที่กลายพันธุ์ - เช่น ไซต์สำหรับ miR-17 - ใน 3′UTR ของยีนที่ส่งออกเช่น PPARG หรือ BMP4 นักวิจัยสามารถเลือกระดับการแสดงออกจากห้องสมุดตัวแปร [5]. ซึ่งทำให้การเหนี่ยวนำเซลล์ไขมันเหมือนสวิตช์หรี่ไฟมากกว่าสวิตช์ไฟ แทนที่จะผลักดันเซลล์ให้ตอบสนองแบบทั้งหมดหรือไม่มีเลย ทีมสามารถปรับการสร้างเซลล์ไขมันได้อย่างระมัดระวังมากขึ้น [5].
ไฟโบรบลาสต์ก็ไม่ใช่แค่ผู้สังเกตการณ์ที่นี่ พวกเขาจัดหาโปรตีน ECM ที่กำหนดเนื้อสัมผัส [10]. ซึ่งทำให้การควบคุมการสร้างเส้นใยเป็นส่วนหนึ่งของการออกแบบผลิตภัณฑ์ ไม่ใช่แค่ประเด็นข้างเคียง วงจรสามารถช่วยจัดการการเปลี่ยนแปลงระหว่างสถานะการสร้างเส้นใยและการสร้างเซลล์ไขมัน และในสัตว์ปีกที่เพาะเลี้ยง การกระตุ้น PPARG โดยตรงในไฟโบรบลาสต์อาจจำเป็นเพื่อสร้างการสะสมไขมันที่มีความหมาย [10].
สถาปัตยกรรมแบบโบว์ไทเหมาะสมกับปัญหานี้เพราะช่วยให้การรับรู้และการส่งออกแยกออกจากกัน ชั้นการรับรู้จะอ่านสถานะปัจจุบันของเซลล์ ในขณะที่ชั้นการส่งออกจะปรับ PPARG, CEBPA หรือผู้ควบคุมสายพันธุ์อื่นๆ การแยกนี้ช่วยหยุดโปรแกรมการสร้างไขมันหรือเนื้อเยื่อเกี่ยวพันจากการเปิดใช้งานก่อนที่เซลล์จะถึงขั้นพัฒนาที่ถูกต้อง
3.3 การควบคุมอัตราส่วนหลายสายพันธุ์และการรับรู้การตอบกลับ
วงจรควบคุมอัตราส่วนจัดการกับปัญหาที่แตกต่าง: ไม่ใช่ว่าการแยกแยะเกิดขึ้นหรือไม่ แต่เป็นว่าการผสมผสานประชากรสุดท้ายยังคงอยู่ในที่ที่ควรจะเป็นหรือไม่ สำหรับผลิตภัณฑ์ที่เป็นชิ้นเดียว การได้รับกล้ามเนื้อ ไขมัน และ ECM ในสัดส่วนที่ถูกต้องมีความสำคัญพอๆ กับการทำให้สายพันธุ์ใดสายพันธุ์หนึ่งแยกแยะออกมา
ระบบเหล่านี้สร้าง การควบคุมการตอบกลับ เข้าไปในเซลล์เอง โปรโมเตอร์เฉพาะสถานะจำกัดการแสดงออกของโปรตีนสัญญาณไปยังเซลล์ที่ได้มุ่งมั่นไปยังสายพันธุ์ที่กำหนดแล้วโมดูลพาราครายน์สังเคราะห์ทำให้เซลล์กล้ามเนื้อที่มุ่งมั่นปล่อยสัญญาณยับยั้งที่กดการมุ่งมั่นของเซลล์ไขมันในเซลล์ใกล้เคียง ตรรกะนี้คล้ายกับการยับยั้งด้านข้างในระบบพัฒนาการ Delta-Notch [1][6]. เมื่อการแตกแขนงซับซ้อนมากขึ้น ประตูหลายอินพุตสามารถรวมสัญญาณภายนอกกับสัญญาณสถานะภายใน [9].
miSFITs ยังทำงานในขั้นตอนนี้ โดยการปรับความแรงของการส่งออกของ BMP4 หรือมอร์โฟเจนอื่น ๆ ทีมสามารถเปลี่ยนสมดุลสายพันธุ์โดยไม่ต้องเขียนตรรกะการตัดสินใจต้นน้ำใหม่ ในเครือข่ายการควบคุมยีนที่ออกแบบ การควบคุมประเภทนี้เพิ่มประสิทธิภาพการแยกสายพันธุ์เป้าหมายจาก 52% ในการควบคุมเป็น 81% ใน iPSCs ที่ออกแบบ [2]. ในเซลล์ต้นกำเนิดมีเซนไคม์ที่ดัดแปลง วงจรสังเคราะห์ขับเคลื่อนประสิทธิภาพการแยกสายพันธุ์หัวใจถึง 76% [2].
ตารางด้านล่างเปรียบเทียบวิธีการแบบสายพันธุ์เดียวและการควบคุมอัตราส่วนโดยใช้จุดที่สำคัญที่สุดในการผลิต
| คุณสมบัติ | วงจรสายพันธุ์เดียว | วงจรควบคุมอัตราส่วน |
|---|---|---|
| ความซับซ้อน | ต่ำ; โดยทั่วไปมีโปรโมเตอร์และตัวควบคุมที่สามารถเหนี่ยวนำได้เพียงตัวเดียว [4] | สูง; ต้องการเกตลอจิกและการส่งสัญญาณพาราคริน [6][9] |
| ภาระการตรวจสอบ | ต่ำ; โดยปกติติดตามผู้รายงานเพียงตัวเดียว [4] | สูง; ต้องการการติดตามตัวบ่งชี้สายพันธุ์หลายตัว [5] |
| ความทนทาน | ปานกลาง; มีแนวโน้มที่จะเกิดความไม่สม่ำเสมอและการสร้างเซลล์สำรอง [3] | สูง; ใช้ข้อเสนอแนะและการยับยั้งด้านข้างเพื่อปรับสมดุลประชากร [1] |
| มูลค่าการผลิต | สูงสำหรับชีวมวล; จำกัดสำหรับโครงสร้างเนื้อเยื่อที่ซับซ้อน [10] | จำเป็นสำหรับผลิตภัณฑ์ที่ต้องการกล้ามเนื้อ ไขมัน และ ECM [4] |
วงจรควบคุมอัตราส่วนเพิ่มภาระการตรวจสอบที่หนักขึ้นแต่การตอบรับในตัวของพวกเขานั้นยากที่จะจับคู่กับการควบคุมกระบวนการเพียงอย่างเดียว ซึ่งทำให้เกิดแรงกดดันมากขึ้นในการเลือกโคลนและการทดสอบกระบวนการ
4. จากการสร้างสู่กระบวนการ: การตรวจสอบความถูกต้อง, การขยายขนาด และความเหมาะสมด้านกฎระเบียบ
4.1 กลยุทธ์การบูรณาการและการเลือกโคลนเพื่อประสิทธิภาพที่เสถียร
หลังจากการออกแบบวงจร ส่วนที่ยากเริ่มต้น: การนำวงจรนั้นเข้าสู่เซลล์ในลักษณะที่คงเสถียรภาพผ่านการส่งต่อในระดับการผลิต
การส่งผ่าน Lentiviral มักมีประสิทธิภาพและให้การรวมตัวที่เสถียรได้อย่างรวดเร็ว แต่การรวมตัวนั้นเป็นแบบสุ่ม ซึ่งหมายถึงความสนใจด้านกฎระเบียบที่มากขึ้น รวมถึงความเสี่ยงที่การแสดงออกจะลดลงเมื่อเวลาผ่านไปเนื่องจากการปิดเสียง ระบบ Transposon เช่น PiggyBac และ Sleeping Beauty อยู่ในระดับกลาง พวกเขาสามารถรักษาประสิทธิภาพผ่านการส่งต่อหลายครั้ง แต่คุณยังคงต้องคัดกรองจำนวนสำเนาและตำแหน่งการแทรก Sleeping Beauty , ตัวอย่างเช่น ได้ถูกใช้ในการทำให้เซลล์ดาวเทียมของวัวคงอยู่ตลอดไปโดยการแสดงออกมากเกินไปของ TERT และ CDK4 โดยที่สายเซลล์ยังคงมีศักยภาพในการสร้างกล้ามเนื้อเกินกว่า 40 ครั้ง [10]. CRISPR knock-in ให้การควบคุมที่แน่นหนาที่สุดเกี่ยวกับตำแหน่งที่โครงสร้างลงจอดและการตั้งค่าทางพันธุกรรมที่แม่นยำที่สุด แม้ว่าการเลือกโคลนจะช้ากว่าและผลผลิตจะต่ำกว่า
| วิธีการบูรณาการ | การควบคุมการแทรก | ความเสถียร | ความสามารถในการขยายตัว | ข้อพิจารณาด้านกฎระเบียบ |
|---|---|---|---|---|
| การส่งผ่าน Lentiviral | ต่ำ (การบูรณาการแบบสุ่ม) | สูง แต่มีแนวโน้มที่จะถูกปิดเสียง | สูง | การตรวจสอบด้านกฎระเบียบที่เข้มงวดขึ้นเนื่องจากการแทรกแบบสุ่มและเศษไวรัส |
| Transposons (PiggyBac/SB) | ปานกลาง | สูงในหลายๆ การส่งผ่าน | สูง | ต้องการการคัดกรองจำนวนสำเนาและตำแหน่งการแทรก |
| CRISPR Knock-in | สูง (เฉพาะจุด) | สูงมาก | ปานกลาง | เป็นที่น่าพอใจ; ลดความเสี่ยงในการรบกวนยีนภายใน |
| เวกเตอร์ Episomal | ไม่มี (นอกโครโมโซม) | ต่ำ; อาจสูญหายระหว่างการแบ่งเซลล์ | ต่ำ | อุปสรรคการรวมต่ำกว่า แต่ไม่เหมาะสำหรับการขยายระยะยาว |
การคัดกรองโคลนจำเป็นต้องทำมากกว่าการยืนยันว่ามีโครงสร้างอยู่มันควรติดตามการเปลี่ยนแปลงของการแสดงออก, โปรไฟล์การแทรก, จลนศาสตร์การเจริญเติบโต, ประสิทธิภาพการแยกแยะ, และการคงอยู่ของลักษณะทางพันธุกรรมที่จำนวนการผ่านสูง snRNA-seq มีประโยชน์ที่นี่เพราะสามารถตัดออกโคลนที่มีเซลล์สำรอง Pax7⁺/Ki-67⁻ ที่ออกจากวงจรเซลล์โดยไม่แยกแยะ ก่อนการขยายขนาด [3]. Porcine EPSCs with a Tet-On-PAX7 circuit retained high muscle differentiation in 3D microcarrier and suspension culture over 40 passages [8].
4.2 การเปลี่ยนแปลงพฤติกรรมของวงจรใน 3D, วัฒนธรรมไมโครแคเรียร์ และไบโอรีแอคเตอร์
เมื่อคุณมีโคลนแล้ว การทดสอบถัดไปคือว่ามันมีพฤติกรรมเหมือนเดิมนอก 2D หรือไม่ ในหลายกรณี มันไม่เป็นเช่นนั้น ประสิทธิภาพใน 2D มักไม่สามารถถ่ายโอนไปยังการแขวนลอย, ไมโครแคเรียร์, หรือวัฒนธรรมโครงสร้างได้อย่างสะอาด เพราะความลาดชันของการแพร่, ขีดจำกัดของออกซิเจน, และแรงเฉือนทั้งหมดเปลี่ยนผลลัพธ์ของวงจร
หนึ่งในการตรวจสอบแรกคือการแพร่กระจายของตัวเหนี่ยวนำ ในเครื่องปฏิกรณ์ชีวภาพแบบถังคน ตัวเหนี่ยวนำโมเลกุลขนาดเล็กจำเป็นต้องเข้าถึงเซลล์อย่างสม่ำเสมอ ในทางปฏิบัติ อาจเกิดการไล่ระดับ โดยเฉพาะในวัฒนธรรมไมโครแคเรียร์ที่หนาแน่นและภายในกลุ่มหรือแกนโครงสร้าง การเพาะเลี้ยงแบบแขวนลอยมักจะเหมาะสมกว่าสำหรับการผลิตเนื้อสัตว์ขนาดใหญ่เนื่องจากรองรับความหนาแน่นของเซลล์ที่สูงขึ้นและให้การควบคุมกระบวนการที่แน่นหนาขึ้น
การตรวจสอบสถานะเซลล์ก็ยากขึ้นเมื่อระบบขยายขนาด สัญญาณรายงานฟลูออเรสเซนต์ที่อ่านง่ายด้วยกล้องจุลทรรศน์ใน 2D อาจยากที่จะตีความในโครงสร้าง 3D ที่ทึบแสง ตัวจับเวลาเรืองแสง - โพรบที่เปลี่ยนการปล่อยจากสีน้ำเงินเป็นสีแดงเมื่อโปรตีนสุก - สามารถให้ข้อมูลการเปิดใช้งานวงจรแบบเรียลไทม์ ในสถานที่ [1]. เส้นทางที่สมเหตุสมผลคือการตรวจสอบแบบเป็นขั้นตอน: เริ่มจากใน 2D, จากนั้นในโครงสร้าง 3D, และสุดท้ายภายใต้สภาวะของไบโอรีแอคเตอร์ [3][8].
4.3 การจำแนกประเภท, เอกสารความปลอดภัยของอาหาร และการพิจารณาของสหราชอาณาจักร/สหภาพยุโรป
หลังจากการทดสอบกระบวนการ การจำแนกประเภทต้องแสดงให้เห็นว่าฟังก์ชันวงจร, ฟีโนไทป์, และความปลอดภัยยังคงอยู่ ข้อมูลหลักควรรวมถึงโฟลไซโตเมทรี, qPCR กับลำดับ cm, RNA-seq time courses, และการอ่านผลการทำงานเช่นพื้นที่เศษส่วนของโซ่หนักไมโอซินและการแสดงออกของไมโอโกลบิน [1]. สื่อการแยกเซลล์ที่ปรับให้เหมาะสมโดยไม่ใช้เซรั่มได้แสดงให้เห็นว่าสามารถนำการแสดงออกของไมโอโกลบินไปสู่ประมาณ 30% ของระดับที่พบในกล้ามเนื้อโคพื้นเมือง [3]. นั่นให้ทีมมีเกณฑ์มาตรฐานที่ชัดเจนแทนที่จะเป็นเป้าหมายที่คลุมเครือ
คุณยังจำเป็นต้องบันทึกโปรไฟล์ของโปรตีน กรดอะมิโน และไขมัน พร้อมกับลักษณะทางประสาทสัมผัส [10][3].
จากมุมมองด้านกฎระเบียบ สหราชอาณาจักรและสหภาพยุโรปได้กำหนดเส้นแบ่งที่ชัดเจนระหว่างสายเซลล์ที่เป็นอมตะโดยธรรมชาติ (ไม่ใช่ GMO) และสายเซลล์ที่ถูกดัดแปลงพันธุกรรม ซึ่งสายเซลล์ที่ถูกดัดแปลงพันธุกรรมต้องการเอกสารความปลอดภัยที่กว้างขวางกว่า [10][3]. แพ็คเกจความเสถียรควรแสดงการคงลักษณะทางฟีโนไทป์และความเสถียรของจีโนมตลอดสะพานการผลิตทั้งหมด - จากธนาคารเซลล์หลักไปจนถึงเซลล์การผลิตสุดท้าย - และบันทึกการติดตามต้องคำนึงถึงทุกขั้นตอนระหว่างนั้น [10]. หากวงจรขึ้นอยู่กับสารกระตุ้นทางเคมี ควรเลือกใช้สารเติมแต่งที่ปลอดภัยต่ออาหารหรือได้รับอนุญาต เช่น กรดวานิลลิก แทนที่จะใช้ด็อกซีไซคลิน [1].
การเฝ้าระวังทางพันธุกรรมอย่างสม่ำเสมอเป็นสิ่งจำเป็น และควรมีการบันทึกสวิตช์การทำลายตัวเองหรือการกำจัดที่สามารถกระตุ้นได้เป็นมาตรการควบคุมความเสี่ยงหลัก [7]. ฟังก์ชันของมันควรรวมอยู่ในเอกสารความปลอดภัยด้วย โดยเฉพาะอย่างยิ่งเมื่อกฎระเบียบของสหราชอาณาจักรและสหภาพยุโรปสำหรับเนื้อสัตว์ที่เพาะเลี้ยงยังคงอยู่ในระหว่างการพัฒนา
5. แผนงานที่ปฏิบัติได้จริงและบทสรุป
5.1 แผนงานการดำเนินการเป็นขั้นตอนสำหรับทีมเนื้อสัตว์ที่เพาะเลี้ยง
เส้นทางที่สะอาดที่สุดจากแนวคิดสู่การผลิตคือการทำงานเป็นขั้นตอน
ขั้นตอนที่ 1 คือการออกแบบ เริ่มต้นด้วยการกำหนดสายพันธุ์เป้าหมาย จากนั้นใช้ snRNA-seq เพื่อยืนยันคอขวดหลักก่อนที่จะเลือกสถาปัตยกรรมวงจร ขั้นตอนนี้สำคัญเพราะวงจรสามารถแก้ไขข้อจำกัดที่คุณได้ระบุไว้เท่านั้น
ขั้นตอนที่ 2 คือการสร้างและการตรวจสอบ 2D สร้างโครงสร้างและตรวจสอบว่าวงจรทำงานตามที่ตั้งใจไว้ใน 2D โดยใช้การอ่านค่ารายงานที่ง่ายในขั้นตอนนี้ เป้าหมายคือการยืนยันว่าโลจิกทำงานได้ก่อนที่จะย้ายไปยังโมเดลที่ยากและมีค่าใช้จ่ายสูงกว่า
เฟส 3 คือการทดสอบความเครียดที่เกี่ยวข้องกับการขยายขนาด เปลี่ยนไปใช้ระบบ 3D และสภาวะที่เกี่ยวข้องกับไบโอรีแอคเตอร์ จากนั้นเปรียบเทียบผลลัพธ์กับฐานข้อมูล 2D นี่คือจุดที่การออกแบบหลายอย่างเริ่มแสดงจุดอ่อน โดยเฉพาะเมื่อมีการถ่ายโอนมวล แรงเฉือน และผลกระทบของเมทริกซ์เข้ามาเกี่ยวข้อง
เฟส 4 คือการบูรณาการด้านกฎระเบียบและความปลอดภัย และควรดำเนินการควบคู่ไปกับเฟส 3 งานด้านความปลอดภัยและกฎระเบียบไม่ควรรอจนถึงตอนท้าย ควรดำเนินการควบคู่ไปกับการขยายขนาด รวมถึงการจัดทำเอกสารสำหรับโมดูลความปลอดภัยที่สามารถเหนี่ยวนำได้
5.2 การจัดหาวัสดุและเครื่องมือที่จำเป็นผ่าน Cellbase
เมื่อกำหนดเวิร์กโฟลว์แล้ว การจัดหามักจะกลายเป็นขั้นตอนที่จำกัดอัตรา
- สายเซลล์
- สื่อที่ปราศจากเซรั่มและกำหนดทางเคมี
- โครงสร้างรองรับ
- ส่วนประกอบของเครื่องปฏิกรณ์ชีวภาพ
- เซ็นเซอร์
- อุปกรณ์วิเคราะห์
การเข้าถึงวัสดุที่เข้ากันได้อย่างเชื่อถือได้ในแต่ละขั้นตอนมีผลโดยตรงต่อความเร็วในการวิเคราะห์พฤติกรรมของวงจรภายใต้สภาวะที่เกี่ยวข้องกับขนาด
5.3 ประเด็นสำคัญ
วงจรยีนสังเคราะห์ให้ทีมเนื้อสัตว์เพาะเลี้ยงควบคุมการตั้งโปรแกรมได้เหนือเวลา เกณฑ์ และสมดุลของสายพันธุ์ที่โปรโตคอลที่ใช้สื่อเพียงอย่างเดียวไม่สามารถเทียบได้ การเลือกสถาปัตยกรรมมีผลต่อการกลับคืนสภาพ การรั่วไหล และความปลอดภัยระบบที่สามารถเหนี่ยวนำได้มักจะได้รับความนิยมเพราะให้การควบคุมตามเงื่อนไขและมีภาระทางเมตาบอลิซึมที่ต่ำกว่า [6].
"เครื่องมือสังเคราะห์ชีววิทยาสามารถใช้ในการสร้างสายเซลล์ที่มีการแสดงออกของยีนที่ปรับแต่งได้ ซึ่งเมื่อรวมกับ PAT และการสร้างแบบจำลองเชิงคำนวณ สามารถทำให้ระบบควบคุมแบบวงปิดสามารถส่งมอบผลผลิตและคุณภาพของผลิตภัณฑ์ที่เหมาะสมที่สุด" - npj Systems Biology and Applications [6]
การใช้งานที่ประสบความสำเร็จไม่ใช่ปัญหาทางชีววิทยาเพียงอย่างเดียว มันขึ้นอยู่กับการเชื่อมโยงอย่างแน่นหนาระหว่างวิศวกรรมวงจร การออกแบบกระบวนการชีวภาพ เอกสารกำกับดูแล และการจัดซื้อจัดจ้าง
คำถามที่พบบ่อย
วงจรยีนสังเคราะห์ช่วยปรับปรุงความสม่ำเสมอของการแยกแยะได้อย่างไร?
วงจรยีนสังเคราะห์สามารถทำให้การแยกแยะมีความสม่ำเสมอมากขึ้นเพราะพวกเขาให้คุณ การควบคุมที่ตั้งโปรแกรมได้ เกี่ยวกับพฤติกรรมของเซลล์และการมอบหมายสายพันธุ์ในการปฏิบัติ นั่นหมายถึงการใช้การดำเนินการเชิงตรรกะแบบโมดูลาร์เพื่อปรับการแสดงออกของยีนและปัจจัยการถอดรหัสด้วยการจับเวลาที่แน่นหนา
การจับเวลานั้นมีความสำคัญ มันช่วยให้เซลล์เคลื่อนผ่านการเปลี่ยนแปลงสถานะที่กำหนดในลำดับที่ถูกต้อง แทนที่จะล่องลอยไปสู่ชะตากรรมที่ผสมหรือไม่ต้องการ นอกจากนี้ยังลดการแยกแยะที่ไม่ตรงเป้าหมายและลดเสียงรบกวนทั่วทั้งวัฒนธรรม
ผลตอบแทนเป็นเรื่องง่าย: ประชากรเซลล์ที่มีความสม่ำเสมอ มั่นคง และเติบโตเต็มที่มากขึ้น สำหรับการผลิตเนื้อสัตว์ที่เพาะเลี้ยง
การออกแบบวงจรใดที่เหมาะกับการควบคุมกล้ามเนื้อหรือไขมัน?
ในการวิจัยเนื้อสัตว์ที่เพาะเลี้ยง ไฟโบรบลาสต์ของไก่ชนิดเดียวกันสามารถถูกผลักเข้าสู่สายใดสายหนึ่ง การสร้างกล้ามเนื้อ ตามโปรโตคอลการเหนี่ยวนำชุดหนึ่ง ในขณะที่ การสร้างไขมัน สามารถเปิดใช้งานได้โดยการเปิดเผยเซลล์ต่ออินพุตเช่นเซรั่มไก่หรือกรดไขมัน
จากนั้น ชะตากรรมของเซลล์เหล่านี้สามารถควบคุมได้ทีละขั้นตอนภายในโครงสร้างไฮโดรเจล 3 มิติ เพื่อสร้างโครงสร้างเนื้อสัตว์ที่มีอัตราส่วนไขมันและคอลลาเจนที่กำหนดไว้
ทำไมวงจรยีนจึงมักทำงานแตกต่างกันในวัฒนธรรม 3 มิติ?
ใน วัฒนธรรม 3 มิติ , วงจรยีนมักทำงานแตกต่างกันเพราะเซลล์กำลังจัดการกับข้อมูลทางกายภาพและโครงสร้างที่ไม่มีอยู่ในชั้นเดียว 2 มิติ ข้อมูลเหล่านั้นรวมถึงแรงดึงเชิงกล ความเครียดเฉือน ความแข็งของเมทริกซ์ และความหนาแน่นของเซลล์ในท้องถิ่น
สัญญาณเหล่านี้สามารถเปลี่ยนเส้นทางการส่งสัญญาณ เช่น Notch และยังสามารถเปลี่ยนวิธีที่วงจรสังเคราะห์ตรวจจับแรงและประสานการตอบสนองที่ตามมา รวมถึงการยึดเกาะระหว่างเซลล์และการสร้างเนื้อเยื่อ









