

如果我去除血清但保留相同的野生型细胞系,我不应期望仅靠培养基调整来阻止衰老、漂移或附着损失。 在这篇文章中,我展示了在培养肉中无血清成功通常依赖于 系统的两方面:细胞外的定义培养基,以及细胞内的编辑帮助其持续分裂、保持附着并保留肌源性功能。
对于生物工艺工程师和细胞培养团队,核心要点很简单:
- 无血清培养基改变细胞行为, 不仅仅是成分列表。在无血清条件下,葡萄糖、谷氨酰胺、甘氨酸和胱氨酸的摄取可能会发生变化。
- 初级卫星细胞在细胞系限制中早期达到极限。 野生型猪细胞通常在 传代10. 左右失去肌源性特征。
- CDKN2A 基因敲除是文章中最明显的例子之一 :编辑的猪卫星细胞系扩增超过 15+ 代, 保持 >90% 的活力, ,在某些克隆中显示出 ~194 倍更高的 PAX7 在 第20代 比野生型对照组。
- 存在权衡。 更好的扩增并不保证晚期代次的分化;一些编辑的细胞系在 第30代. 仍然显示出较低的分化。
- 验证必须在生产设置中进行: 相同的无血清培养基、相同的培养模式,以及相同的生长、废物积累、谱系标记和融合的读数。
简而言之:如果您想要一个可以转移到生产中的无血清工艺,我会将培养基设计、基因编辑、克隆筛选和生物反应器适配视为一个连贯的工作流程,而不是四个独立的工作。
| 关注领域 | 我会首先检查的内容 | 重要性原因 |
|---|---|---|
| 细胞周期控制 | CDKN2A, 传代寿命,PAX7 | 帮助显示细胞系是否可以在不早衰的情况下扩展 |
| 生长信号 | IGF1R, EGFR, FGFR 响应 | 无血清系统具有较低的外部信号支持 |
| 应激存活 | 活力,凋亡标志物,剪切响应 | 血清撤除和传代可能导致细胞丢失 |
| 营养处理 | 葡萄糖使用,乳酸,氨,氨基酸摄取 | 更快的摄取也可能意味着更快的废物积累 |
| 身份保留 | PAX7, MYOD, MYOG, 融合指数 | 一条快速增长的细胞系如果不再形成目标组织 | 是没有用的。
然后,我会讲解无血清培养失败的地方,这些失败点对应的编辑,以及在工艺转移之前我将如何验证一个编辑过的细胞系。
哺乳动物细胞系中的CRISPR-Cas基因组编辑 | 协议预览
无血清培养的主要生物学障碍
去除血清暴露了三个瓶颈:信号传导、附着和细胞身份。问题始于细胞内部, ,不仅仅是在培养基配方中。这很重要,因为它决定了团队花费时间的方向:培养基调整、基因编辑,或两者的结合。
| 特征 | 血清补充培养 | 无血清培养 |
|---|---|---|
| 增殖 | 强健;由多种生长因子支持 | 可变;易于复制性衰老和G1/S阻滞 |
| 附着 | 由血清中的ECM蛋白(纤维连接蛋白、玻连蛋白)支持 | 需要外源涂层或添加剂;脱离风险增加 |
| 营养运输 | 由载体蛋白如白蛋白和转铁蛋白促进 | 依赖于较少缓冲的摄取;需要优化的ITS-X和脂质浓度 |
| 凋亡风险 | 低;PI3K-AKT和MAPK-ERK通路强烈激活 | 更高的;对氧化应激和代谢废物的敏感性增加 |
| 身份稳定性 | 通常在早期到中期传代中保持稳定 | 表型漂移风险高;干性标志物通常迅速下降 |
生长和存活信号的丧失
一旦去除血清,生长因子水平急剧下降。细胞随后失去许多外部支持,这些支持有助于保持 PI3K-AKT 和 MAPK-ERK 活性高。在实践中,这意味着更多的凋亡和较弱的增殖,这对于扩大规模是一个直接的问题。
粘附、营养摄取和应激瓶颈
血清不仅仅是为细胞提供营养。它还提供支持附着和扩展的 ECM 蛋白。没有纤维连接蛋白、玻连蛋白和相关因子,初级卫星细胞更容易脱落并进入凋亡,尤其是在生物反应器条件下的剪切力下。使用 Y-27632 抑制 ROCK 可以在一定程度上有所帮助,但不能解决附着问题。
营养处理也变得更加困难。没有血清载体蛋白,葡萄糖、谷氨酰胺、甘氨酸和胱氨酸的摄取变得不那么缓冲 [1] . 同时,代谢废物如氨和乳酸可能会积累并抑制生长[3]. 因此,即使基础培养基在纸面上看起来不错,运输和废物平衡仍可能成为限制步骤。
无血清适应期间的表型漂移
无血清适应可以选择适应新条件的亚群,但不再符合产品规格。这就是陷阱:细胞可能扩展良好,但失去形成预期组织的能力。
在连续传代过程中,标记如PAX7, MYOD, 和MYOG可能会下降[2]. 在适应过程中跟踪谱系标记,以便漂移在早期显示出来,而不是在长时间的培养基优化周期之后。这些是基因编辑需要稳定的途径。
基因编辑方法提高无血清性能
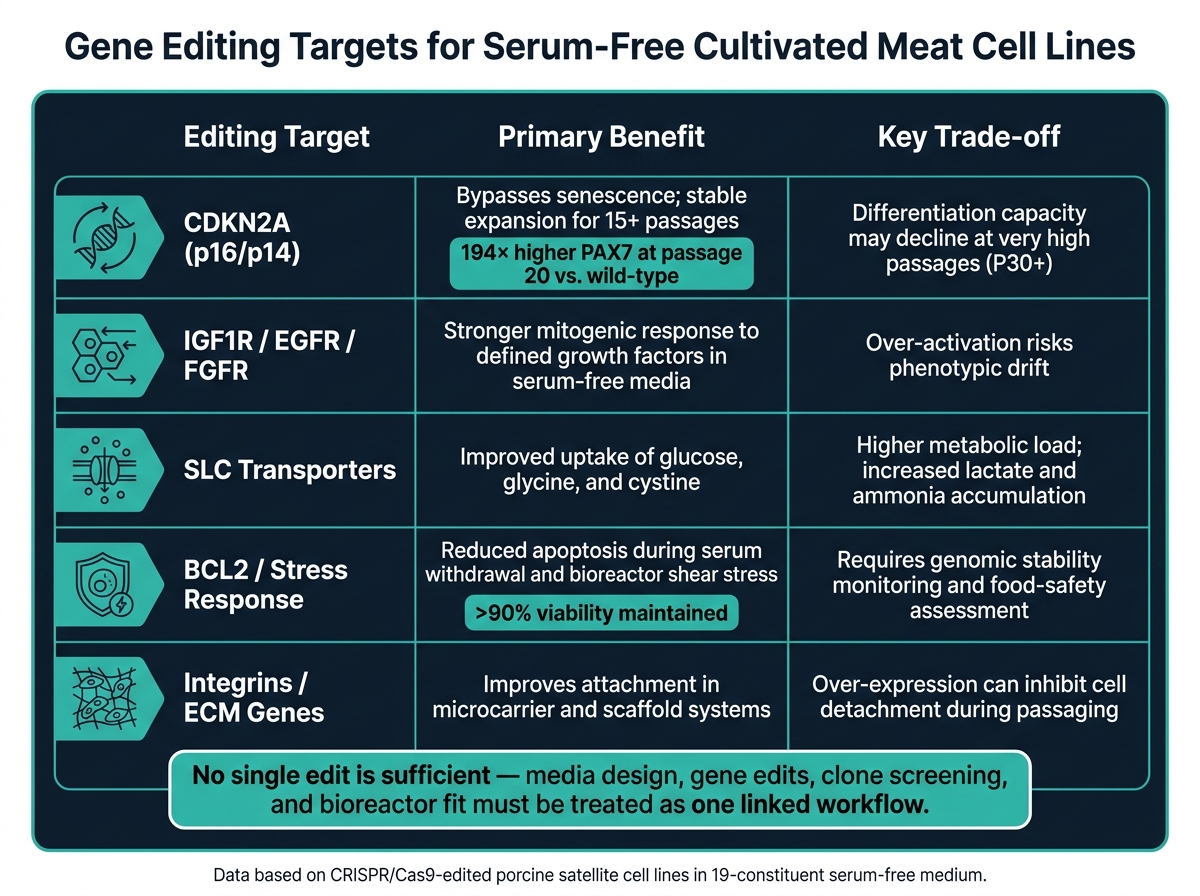
无血清细胞培养的基因编辑目标:收益与权衡
这些障碍分为三类编辑:信号传导、生存和分化。
编辑生长因子信号传导和营养利用
一种直接途径是上调或敏化IGF1R, EGFR, 和FGFR,使细胞对IGF-1、EGF和bFGF反应更强烈[2]. 这在无血清培养基中很重要,因为生长因子水平通常设计得较低。如果信号传导改善,细胞周期控制往往成为下一个瓶颈。
细胞周期调节因子CDKN2A在这里尤为突出。CRISPR/Cas9 敲除外显子2生成了 CDKN2A−/− 猪卫星细胞系,在19种成分的无血清培养基中强烈扩增超过15代。在特定克隆中, PAX7 表达在第20代时比野生型对照上调约194倍 [2].
对溶质载体 (SLC) 转运蛋白的编辑可以帮助避免在葡萄糖、谷氨酰胺、甘氨酸和胱氨酸放大过程中摄取的限制 [1]. 但有一个问题。更高的摄取也会导致乳酸和氨的快速积累,因此转运蛋白的编辑需要从第一天起就与培养基交换和废物控制一起计划。单靠摄取编辑是不够的。
提高细胞存活率和对无血清应激的抵抗力
血清撤除、常规传代和生物反应器剪切都会将细胞推入更高凋亡条件。编辑BCL2通路——通过上调促生存成员或抑制促凋亡成员——可以减少这些过渡期间的细胞损失。这在微载体系统中变得更加重要,因为细胞需要应对附着应力和机械应力。
任何改善生存或延长增殖的编辑都需要在整个制造传代范围内进行基因组稳定性检查。CDKN2A−/−猪卫星细胞在连续无血清增殖期间保持了90%以上的活细胞率 [2]. 即便如此,团队应在设定的传代间隔检查染色体完整性,而不是假设稳定性会持续。
平衡粘附、增殖和分化能力
最困难的部分是管理扩增和分化之间的拉扯。CDKN2A 基因敲除在第10代时保持了肌生成潜力,而在无血清条件下的野生型细胞几乎完全失去了肌生成特性。在编辑的细胞系中,融合指数报告为16.3%到56.3% [2]. 然而,到第30代时,即使是编辑过的细胞也可能表现出分化能力下降 [2].
| 编辑目标 | 无血清培养中的主要优势 | 关键权衡 |
|---|---|---|
| CDKN2A (p16/p14) | 绕过衰老;稳定扩增超过15代[2] | 在非常高的传代数(P30+)时分化能力可能下降[2] |
| IGF1R / EGFR / FGFR | 对特定生长因子的促有丝分裂反应更强[2] | 过度激活有表型漂移风险 |
| SLC 转运蛋白 | 改善葡萄糖、甘氨酸和胱氨酸的摄取[1] | 更高的代谢负荷;乳酸和氨的积累增加[1] |
| BCL2 / 应激反应 | 减少撤退和剪切应力期间的凋亡 [2] | 需要基因组稳定性监测和食品安全评估 [2] |
| 整合素 / ECM 基因 | 改善微载体和支架系统中的附着力 [2] | 过表达可以抑制传代过程中的细胞脱落 [2] |
粘附编辑在微载体或支架设置中最有用。它们更适合作为特定格式的工具,而不是每个无血清过程的解决方案。
可诱导的CRISPR系统为团队提供了一种实用的方法来处理扩增与分化的权衡。这个想法很简单:使用可诱导的编辑将扩增阶段与分化分开。
如果表型在预期的无血清培养基中不成立,这些编辑都无关紧要。
sbb-itb-ffee270
构建和验证用于无血清培养的编辑细胞系
找到合适的编辑只是工作的一部分。更困难的部分是将该编辑转化为能够处理无血清生产的稳定细胞系。这需要一个紧密的工作流程,将编辑、克隆选择和验证链接在一个管道中。而且该管道应直接测试已识别的信号、生存和附着限制。
选择编辑工具和交付方法
对于目标如CDKN2A, CRISPR/Cas9敲除是去除细胞周期抑制因子并支持长期扩展的实际第一步[2]. 在初级牲畜细胞中,常见的交付途径包括非病毒转染系统,如Lipofectamine,以及病毒系统,如lentiCRISPR v2 [2][4]. 在进行克隆工作之前,确认交付效率。
有一点比有时得到的重视更重要:在计划生产的确切培养基和培养模式中筛选每个克隆. 如果制造过程使用定义的无血清培养基、静态贴壁培养、微载体或其他设置,那就是细胞在筛选期间应面对的条件。
在生产无血清配方中筛选编辑过的细胞
一种常见的方法是通过有限稀释分离克隆,然后通过Sanger测序在目标位点确认编辑[2]. 一旦编辑得到验证,筛选应继续在用于生产的相同无血清配方和培养模式中进行[2][1].
在此阶段,测量基本指标,以判断克隆体是否能适应该过程,而不仅仅是存活于编辑中:
- 生长
- 存活率
- 葡萄糖消耗
- 乳酸生成
- 氨积累
也有必要在早期添加 PAX7 RT-qPCR,因为干性丧失可能在细胞系以更明显的方式失败之前出现 [1][2].
在工艺转移前表征编辑细胞
在工艺转移之前,验证应涵盖四个相关领域:基因组编辑、途径响应、传代稳定性和功能。每个领域解决不同的问题。基因组检查处理表型漂移风险。耗尽培养基分析指向营养摄取和废物积累的限制。融合指数告诉您肌源性分化是否仍然存在 [2][1].
| 检测类型 | 测量内容 | 对无血清培养肉系的重要性 |
|---|---|---|
| T7内切酶I / Sanger测序 | 编辑效率和精确的基因组序列 | 在扩大规模前确认基因敲除或敲入的成功 [2] |
| RT-qPCR (PAX7, MYOD, MYOG, BAX, CCND1) | 干性、分化和凋亡标志物的转录水平 | 监测细胞健康和长期传代中的分化潜力 [2][4] |
| 免疫荧光 (MyHC / CK18) | 谱系特异性蛋白表达 | 确保细胞在编辑和适应后保持肌肉或上皮特性 [2][4] |
| 消耗培养基分析 | 葡萄糖、氨基酸、乳酸和氨的分析 | 确定营养需求并指导生物反应器的供料策略 [1] |
| 融合指数 | 多核肌管中核的整合百分比 | 确认在无血清条件下保持肌源性分化能力 [2] |
| 质地剖面分析 (TPA) | 3D结构的硬度、弹性和咀嚼性 | 验证编辑后的细胞能生产出具有肉类物理特性的最终产品 [2] |
基因组验证 依赖于 T7 核酸内切酶 I 测试加上单个克隆的桑格测序 [2]. 通路确认 使用RT-qPCR或Western blot来显示计划的转录本或蛋白质变化确实发生,包括标记如PAX7, MYOD , MYOG 和 MyHC [2][4].
对于长期稳定性 , 基准是15-30次传代,重复检查生长、活力和标记表达。CDKN2A 敲除的猪卫星细胞在无血清条件下保持了超过90%的活细胞率超过15次传代,但分化能力在第30次传代时开始下降 [2].
功能测试 然后提出最简单的问题:这些仍然是您需要的细胞吗?在肌源性细胞系中,融合指数显示编辑后的细胞是否仍能在无血清条件下形成多核肌管 [2] . 纹理剖面分析 (TPA) 然后检查 3D 构造是否显示出类似肉的硬度、弹性和咀嚼性 [2].
使用这些数据来设置克隆的无血清制造转移条件。
从编辑细胞系到无血清制造
将编辑细胞与培养基和生物反应器设计相匹配
一旦克隆通过验证,工作就会改变。此时,成功取决于细胞系与工艺的契合度。更多的筛选无法弥补工艺匹配不佳的问题。
消耗培养基分析应推动葡萄糖补充、氨基酸补充和生长因子剂量,包括定义的输入如 bFGF 和 IGF-1 [2]. 在贴附系统中,支架接种密度和粘附窗口——在生物反应器转移前约2小时——应根据编辑线的附着行为设置,而不是基于含血清的协议[2]. 这些数据应直接用于决定饲料时间、接种密度和转移时间。
编辑线可以支持更长的扩展、更高的细胞密度和更稳定的标记表达。这意味着放大必须遵循编辑线的测量行为,而不是野生型假设。
在实践中,线选择成为采购和放大决策,而不仅仅是生物学决策。
研发、生产和采购团队的关键要点
无血清适应不仅仅是一个培养基配方问题。它始于细胞系,单靠培养基优化无法解决。靶向基因编辑,特别是对细胞周期抑制因子如CDKN2A的敲除,, 解决了导致原代卫星细胞在无血清条件下失效的基础生物学问题。CDKN2A−/−猪卫星细胞在第20代时保持PAX7表达比野生型对照高约194倍,并在第10代时达到高达56.3%的融合指数——此阶段未编辑的细胞已基本丧失成肌功能[2].
对于开发和制造团队来说,分工相当明确:
- 研发& 团队应建立一个验证流程,从一开始就在实际生产条件下测试编辑克隆。这包括生长、营养消耗、谱系稳定性和3D分化能力。
- 生产团队 应使用编辑行的营养成分来设置饲料设计和生物反应器参数,因为从含血清协议中复制的假设不太可能成立 [1].
- 采购团队 需要符合编辑行特定要求的采购计划,包括定义的生长因子、脂质、抗氧化剂以及适合该行粘附特性的支架或微载体。
常见问题解答
为什么单靠培养基优化不够?
单靠培养基优化是不够的。在许多情况下,动物细胞根本不具备大规模生产所需的特性,例如 抗剪切应力, 代谢效率 , 和 在高密度悬浮液中的存活率.
无血清培养基很重要,但它们无法解决细胞的内在限制。这些限制包括有限的增殖寿命, 对生物反应器压力的敏感性, 以及不同物种和发育阶段的营养需求.
在无血清培养中,哪些基因编辑最重要?
在培养肉生产中,最重要的编辑是那些减少对添加生长因子依赖的编辑。一个例子是CDKN2A删除,这可以在无血清条件下改善猪卫星细胞的增殖和分化。另一种方法是工程化肌肉干细胞以诱导过表达FGF2和突变的RasG12V. 这种设置支持自分泌信号传导,并消除了培养基中重组FGF2的需求。
编辑的细胞系应如何验证以用于生产?
编辑的细胞系应进行基因组、蛋白质组和功能测试,以确认其生产性能和分化潜力。
实际上,这意味着要检查编辑是否达到了预期效果而不在其他地方产生问题。研究人员应验证基因修改不会影响向目标组织的分化,并且预期特性(如抗压能力或无血清生长)如预期那样表达。









