

如果您可以扩展细胞但无法在正确的时间将其切换到正确的命运,您的过程将在分化时停滞。 这是核心要点:合成基因电路为您提供 细胞内的承诺、时间、记忆和谱系混合控制,而单靠培养基变化往往会留下 异质性, 部分承诺的群体。
如果我在构建培养肉分化工作流程,我会立即从这篇文章中提取四个要点:
- 从原生网络开始,而不是构建体。 使用 snRNA-seq, 轨迹分析、GRN推断和miRNA分析来找出细胞停滞、漂移或分支到错误命运的地方。
-
将电路类型与过程问题相匹配。
一个拨动开关适合锁定, 前馈或带通设计适合时间控制, 逻辑门适合多信号门控,而 miSFITs适合分级输出。 - 从第一天起就设计低泄漏、低噪音和安全性。 正交部件、负自调节、iFFLs、cm 转基因和可诱导的杀伤或生长抑制模块是构建的一部分,而不是事后的想法。
- 在规模相关条件下尽早验证。 在二维中工作的电路可能会因为诱导剂梯度、氧气限制和剪切而在 三维、微载体或搅拌悬浮液中发生变化。
文章还提出了一个对工艺团队重要的实际观点:单系控制和比例控制是不同的工作. 一个Tet-On MyoD盒可能会推动肌源性进入,但整体切割产品需要控制 肌肉、脂肪和ECM比例, ,这通常意味着反馈、旁分泌信号和更严格的克隆筛选。
一些数据支持了这一信息:
- 标准的肌源性分化可能会停滞在融合指数大约为50-60%
- 在iPSCs中,工程化GRNs将目标谱系分化从52%提高到81%
- 在改造的MSCs中,合成电路将心脏分化驱动到76%
- 一些猪的Tet-On-PAX7系在40次传代后仍保持高肌源性潜力
- 大约20%的人类多能干细胞可能携带与癌症相关的突变,这就是为什么可诱导的安全模块很重要
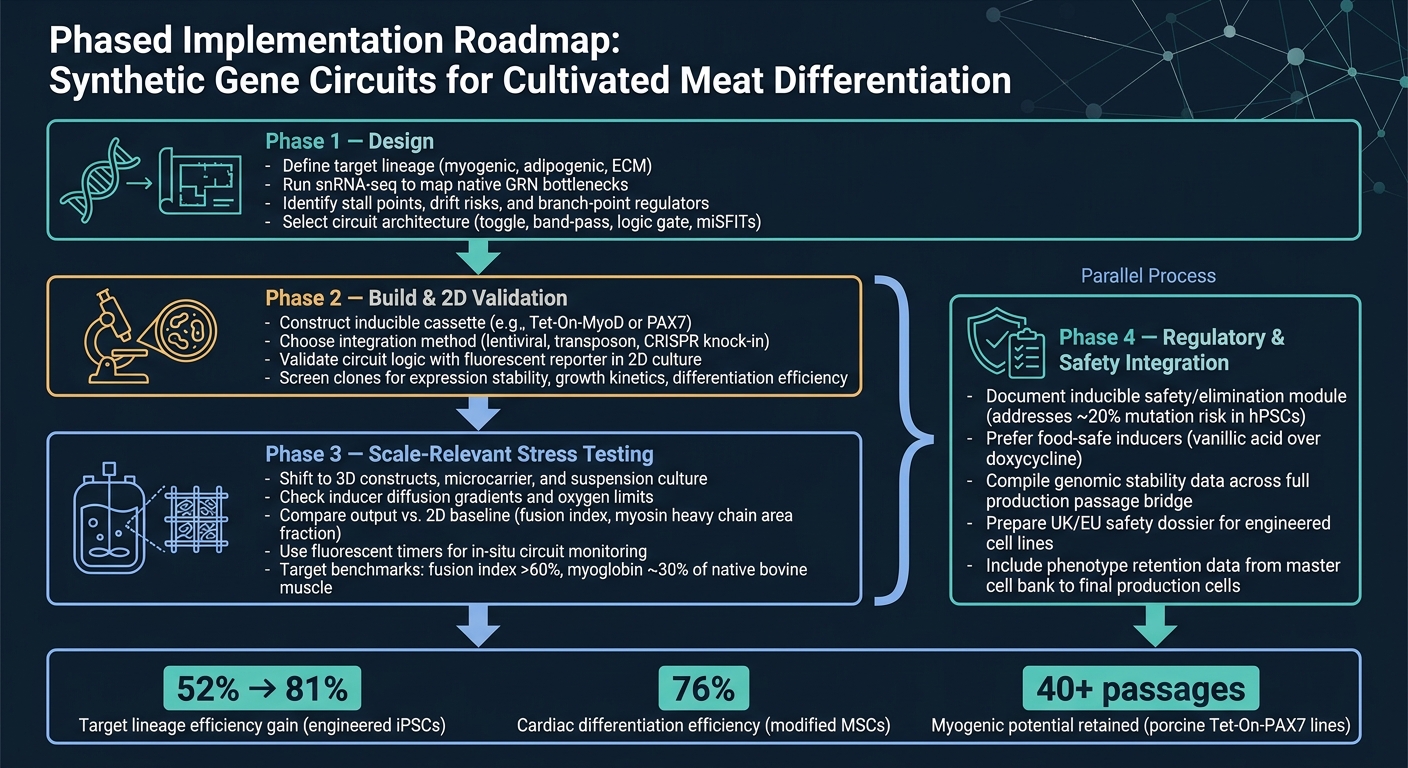
合成基因电路在培养肉分化中的工作流程
科学家故事:Michael Elowitz,活细胞中的遗传电路
快速比较
| 主题 | 文章用简单的术语说 |
|---|---|
| 主要瓶颈 | 差异化,而非扩展 |
| 为什么电路有帮助 | 它们在细胞内增加阈值控制和细胞状态记忆 |
| 最佳第一步 | 使用snRNA-seq和相关工具绘制本地命运控制图 |
| 主要电路选项 | 切换、前馈、带通、逻辑门、miSFITs、CRISPRa/i |
| 肌生成示例 | Tet-On-MyoD将生长与终末分化分开 |
| 脂肪生成/ECM控制 | miSFITs和蝴蝶结设计用于分级PPARG/BMP4型输出 |
| 整体切割挑战 | 肌肉、脂肪和结缔组织之间的比例控制 |
| 放大风险 | 二维行为可能不适用于三维或生物反应器 |
| 集成选择 | 慢病毒、转座子、CRISPR 敲入、非整合载体 |
| 监管点 | 工程化细胞系需要更广泛的安全包;食品安全诱导剂如香草酸优于DOX,尽可能使用 |
所以,简单来说:这不仅仅是一篇电路设计文章. 我会将其视为将电路架构、谱系生物学、克隆选择、生物反应器性能和英国/欧盟安全文件链接到一个差异化策略的指南。
如果您想了解从本地网络映射到构建选择、放大检查和法规适配的完整路径,请继续阅读。
2. 差异化控制电路的设计原则
2.1 在设计电路之前映射本地细胞命运网络
在设计电路之前,您需要清楚了解细胞当前的活动。
单核RNA测序 (snRNA-seq) 是一个不错的起点。它可以显示静止的亚群,包括由NOTCH2 和 HEYL, 标记的储备细胞,并指出可能改善分化的途径目标 [3].
从那里,轨迹分析和基因调控网络(GRN)推断有助于绘制调节器激活的顺序,并突出显示细胞最有可能停滞的地方。在肌生成中,主要级联通过MYOD1和MYOG。在脂肪生成中,主要节点是 PPARG和CEBPA,纤维脂肪生成前体(FAP)命运作为主要分支点风险。下表总结了主要调节器和瓶颈。
| 谱系 | 关键主调控因子 | 关键信号通路 | 识别的瓶颈 |
|---|---|---|---|
| 成肌 | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | 储备细胞形成(静止) |
| 成脂 | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | 纤维-脂肪生成祖细胞(FAP)命运 |
| 多能性 | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | 自发分化/异质性 |
另一个有用的层面是miRNA表达谱分析. 内源性miRNAs如miR-302a, 与多能性相关,以及miR-375, 与分化相关,可以在感知和响应设计中充当内部分类器。 这使得电路可以读取细胞的实际状态,而不仅仅依赖于外部诱导器 [5].
这些瓶颈应该驱动电路的选择。如果主要问题是漂移,您可能需要一个锁定。如果是时间问题,脉冲可能更合适。如果命运控制依赖于多个线索,多输入逻辑通常更有意义。
2.2 选择合适的电路架构
这是权衡取舍显而易见的地方。合适的架构取决于三个实际点:承诺的永久性要求、时间控制的紧密程度以及细胞可以无问题携带的遗传负载量。
双稳态切换开关 适用于需要保持谱系承诺锁定的情况。主要问题是由转录噪声引起的自发状态翻转。
带通滤波器 适用于仅在特定发育窗口内需要转录因子的情况。问题在于诱导剂水平需要严格控制,否则时间会出错。
逻辑门 通过要求同时存在多个输入来增加特异性。例如,只有在存在外源诱导剂并且细胞显示正确的内源miRNA谱时,才允许分化。这有助于降低非目标承诺的风险。
下表概述了主要架构及其权衡。
| 架构 | 可逆性 | 时间精度 | 集成复杂性 | 主要用例 | 关键风险 |
|---|---|---|---|---|---|
| 双稳态开关 | 低(锁定后) | 中等 | 中等 | 永久谱系承诺 | 由于噪声导致的自发翻转 |
| 带通滤波器 | 高(浓度依赖) | 高 | 高 | 瞬时发育阶段 | 需要严格的诱导剂控制 |
| 逻辑门(与/或/非) | 可变 | 中等 | 中等–高 | 细胞类型特异性激活 | 关闭状态下的泄漏性 |
| 蝴蝶结/多输入 | 高 | 中等 | 中等 | 多信号整合 | 依赖于内源性miRNA稳定性 |
| miSFITs | 高 | 中等 | 低–中等 | 分级输出调节 | 如果调节不当,动态范围窄 |
每增加一层调控层都会增加负载并消耗细胞资源。在实际操作中,如果两个设计完成相同的工作,当规模重要时,通常更简单的设计是更好的选择。
一旦架构确定,接下来的任务就是确保其在低泄漏性、噪声抑制和故障安全控制下保持稳定。
2.3 为可靠性、低泄漏性和安全性而构建
电路必须在长时间培养中保持稳定。短暂的良好性能不足以用于生产。
正交部件是第一道防线。不与本地机器交叉反应的启动子、转录因子和调控元件有助于限制非目标效应,并减少内源信号开启或关闭电路的可能性。改良的高紧密性启动子如PCREm已被用于减少可诱导哺乳动物系统中的基础泄漏性[6] .
负反馈自调节在可能的情况下也值得添加。这是减少转录噪声并对诱导剂浓度产生更线性响应的较知名的模式之一[6]. 不一致前馈环路(iFFLs) 可以通过过滤随机波动增加另一层控制,使细胞对持续信号而非短暂噪声尖峰作出反应。
密码子修饰的(cm)合成转录因子的版本也使表征更容易。它们让您在验证过程中将电路驱动的表达与内源基因组( g)表达分开[1]. 这可能听起来像是一个小细节,但当你试图确定读数是来自电路还是宿主基因组时,它可以节省时间。
需要安全模块。 大约20%的人类多能干细胞携带与癌症相关的突变 [7]. 因此,如果电路要进入干细胞衍生的细胞系,它应包括一个可诱导的生长抑制或消除模块。香草酸 是一个值得优先考虑的诱导剂,因为它是一种许可的食品添加剂,这有助于将其用作培养肉细胞系电路触发器的理由 [1].
“合成生物学为工程师提供了手段,可以部署电路以轻松且精确地调节多个基因的表达,以...消除潜在的不良脱靶副作用。" - npj Systems Biology and Applications [6]
这些选择在第3节中建立了谱系特定的电路。
sbb-itb-ffee270
3. 肌源性、脂源性和比例控制分化的电路策略
3.1 将扩展与终末分化分开的肌源性电路
一旦电路架构设置完成,接下来的任务就是谱系特定的部署。对于肌生成,主要问题很简单但执行起来很难:细胞需要先增殖,然后在需要时切换到分化,而不会过早地偏向任何一个方向。
A Tet-On-MyoD cassette 是实现这一目标的最直接方法之一。在这种设置中,当没有多西环素(DOX)时,细胞在标准条件下增殖。添加DOX,电路驱动肌源性承诺。多个中国机构的研究人员在鸡胚胎成纤维细胞中使用了这种方法,并报告称诱导后肌管形成效率很高[4].
一个双稳态开关提供了更严格的状态控制。由相互抑制的抑制因子如E-KRAB和Pip-KRAB构建的系统可以将短暂的DOX脉冲转化为稳定的肌源性程序[6]. 实际上,这意味着分化状态在诱导后保持锁定,而不是在信号移除后消退。添加负自调节也有助于减少随机噪声并限制扩展阶段的泄漏分化 [6].
这很重要,因为标准的体外肌源性分化通常会在中途停滞。融合指数通常只有50-60%左右,这留下了大量未融合的储备群体[3]. 电路驱动的MyoD激活可以提高承诺,但仅有承诺并不能保证均匀融合。当MyoD电路与MEK、NOTCH和RXR调节结合时,在二维培养中几乎可以实现完全融合[3]. 电路控制谱系进入的时机;小分子推动更清洁和更均匀的细胞周期退出。
3.2 脂肪生成和纤维生成的控制用于组成和结构
仅有肌肉是不够的。一旦肌生成控制到位,下一个问题就是组成:形成多少脂肪,沉积多少ECM,以及这些程序何时启动。在这里,二进制的开/关控制通常过于生硬。团队通常需要的是分级输出,特别是在脂肪生成-纤维生成分支点附近。
miSFITs提供了一种实用的方法来逐步调整表达。通过在输出基因(如PPARG或BMP4)的3′UTR中放置突变的miRNA靶位点——例如miR-17的位点,研究人员可以从变异库中选择表达水平[5]. 这使得脂肪细胞诱导更像是一个调光开关而不是一个灯开关。团队可以更仔细地滴定脂肪生成,而不是将细胞推向全有或全无的反应[5].
成纤维细胞在这里也不仅仅是旁观者。它们提供塑造纹理的ECM蛋白[10]. 这使得纤维生成控制成为产品设计的一部分,而不仅仅是一个次要问题。电路可以帮助管理纤维生成和脂肪生成状态之间的转换,在培养的家禽中,可能需要在成纤维细胞中直接激活PPARG以产生有意义的脂肪沉积[10].
蝴蝶结架构非常适合这个问题,因为它将感知和输出分开。感知层读取细胞的当前状态,而输出层调整PPARG、CEBPA或其他谱系调节因子。这种分离有助于防止脂肪生成或结缔组织程序在细胞达到正确发育阶段之前启动。
3.3 多谱系比例控制和反馈感知
比例控制电路处理的是不同的问题:不是分化是否发生,而是最终的群体混合是否保持在应有的位置。对于整切产品,将肌肉、脂肪和ECM按正确比例组合同样重要,就像让任何一个谱系分化一样。
这些系统将反馈控制构建到细胞本身。状态特异性启动子将信号蛋白的表达限制在已经承诺给定谱系的细胞中。合成旁分模块然后让已承诺的肌源性细胞释放抑制信号,抑制附近细胞的脂肪生成承诺。其逻辑类似于Delta-Notch发育系统中的侧向抑制[1][6] . 当分支变得更加复杂时,多输入门可以将细胞外线索与内部状态信号结合[9].
miSFITs也在这个阶段起作用。通过调整BMP4或其他形态发生素的输出强度,团队可以在不重写上游决策逻辑的情况下改变谱系平衡。在工程基因调控网络中,这种类型的控制将目标谱系分化效率从对照组的52%提高到工程化iPSCs的81%[2]. 在改造的间充质干细胞中,合成电路将心脏分化效率提高到76%[2].
下表比较了单系和比率控制方法在生产中最重要的点。
| 特性 | 单系电路 | 比例控制电路 |
|---|---|---|
| 复杂性 | 低;通常是一个可诱导的启动子和调节器[4] | 高;需要逻辑门和旁分泌信号[6][9] |
| 监测负担 | 低;通常跟随一个单一的报告基因[4] | 高;需要跟踪多个系标记[5] |
| 稳健性 | 中等;易于异质性和储备细胞形成[3] | 高;使用反馈和侧抑制来平衡群体 [1] |
| 生产价值 | 生物质高;复杂组织结构有限 [10] | 对于需要肌肉、脂肪和ECM的整切产品是必需的 [4] |
比例控制电路增加了更重的验证负担。但是,他们内置的反馈很难仅通过过程控制来匹配,这给克隆选择和过程测试带来了更大的压力。
4. 从构建到过程:验证、放大和法规适应性
4.1 稳定性能的整合策略和克隆选择
电路设计之后,困难的部分开始了:将电路以一种在生产规模传代中保持稳定的方式引入细胞。
慢病毒递送通常效率高,并能快速提供稳定的整合体。但整合是随机的。这意味着需要更多的法规关注,并且由于沉默的原因,表达可能会随着时间的推移而下降。转座子系统如PiggyBac和Sleeping Beauty处于中间位置。它们可以在多次传代中保持性能,但仍需要筛选拷贝数和插入位点。 睡美人, 例如,通过过表达TERT和CDK4,已被用于稳定地使牛卫星细胞永生化,细胞系在超过40次传代后仍保留肌原性潜力 [10] . CRISPR敲入 提供了对构建体着陆位置的最严格控制和最精确的基因组设置,尽管克隆挑选速度较慢且吞吐量较低。
| 整合方法 | 插入控制 | 稳定性 | 可扩展性 | 监管考虑 |
|---|---|---|---|---|
| 慢病毒递送 | 低(随机整合) | 高,但易于沉默 | 高 | 由于随机插入和病毒残留,监管审查更严格 |
| 转座子(PiggyBac/SB) | 中等 | 在多次传代中保持高水平 | 高 | 需要筛查拷贝数和插入位点 |
| CRISPR 敲入 | 高(位点特异性) | 非常高 | 中等 | 有利;降低破坏内源基因的风险 |
| 外源性载体 | 无(染色体外) | 低;在分裂过程中可能丢失 | 低 | 整合障碍较低,但不适合长期扩展 |
克隆筛选需要做的不仅仅是确认构建体的存在。它应该跟踪表达漂移、插入特征、增长动力学、分化效率和高传代数的表型保留。snRNA-seq在这里很有用,因为它可以排除富集Pax7⁺/Ki-67⁻储备细胞的克隆——这些细胞在不分化的情况下离开细胞周期——在扩大规模之前。[3]. 含有Tet-On-PAX7电路的猪EPSCs在3D微载体和悬浮培养中超过40代保持了高肌肉分化能力[8].
4.2 电路行为在3D、微载体和生物反应器培养中的变化
一旦你有了一个克隆,接下来的测试是它在2D之外是否表现相同。在许多情况下,它并不如此。2D中的性能很少能干净地转移到悬浮、微载体或支架培养中,因为扩散梯度、氧气限制和剪切力都会改变电路输出。
首先的检查之一是诱导剂扩散。在搅拌罐式生物反应器中,小分子诱导剂需要均匀地到达细胞。在实际操作中,尤其是在密集的微载体培养和聚集体或支架核心内部,可能会形成梯度。悬浮培养通常更适合大规模培养肉类,因为它支持更高的细胞密度并提供更严格的过程控制。
随着系统规模的扩大,监测细胞状态也变得更加困难。在二维显微镜下易于读取的荧光报告信号在不透明的三维结构中可能难以解析。荧光计时器——随着蛋白质成熟从蓝色转为红色的探针——可以提供实时电路激活数据原位 [1]. 合理的路线是分阶段验证:首先在二维,然后在三维结构中,然后在最终的生物反应器条件下 [3][8] .
4.3 表征、食品安全文件和英国/欧盟的考虑
在工艺测试之后,表征必须显示电路功能、表型和安全性仍然有效。核心数据堆栈应包括流式细胞术、带有 cm 序列的 qPCR、RNA-seq 时间过程和功能读数,如肌球蛋白重链面积分数和肌红蛋白表达 [1]. 优化的无血清分化培养基已被证明可将肌红蛋白表达提高到约 30% 的原生牛肌肉水平 [3]. 这为团队提供了一个明确的基准,而不是一个模糊的目标。
您还需要记录蛋白质、氨基酸和脂肪的特征,以及感官特性[10][3].
从监管的角度来看,英国和欧盟在自发永生化(非转基因)细胞系和基因工程细胞系之间划出了一条明确的界限。后者需要更广泛的安全档案[10][3]. 稳定性方案应显示从主细胞库到最终生产细胞的整个生产桥梁的表型保留和基因组稳定性,并且可追溯性记录需要说明每次传代[10]. 如果电路依赖于化学诱导剂,食品安全或许可的添加剂如香草酸比多西环素更可取[1].
常规基因组监测是必须的,并且应将可诱导的自杀或消除开关记录为核心风险控制措施[7]. 其功能也应包含在安全档案中,特别是在英国和欧盟对培养肉的法规不断成形之际。
5. 实用路线图和结论
5.1 培养肉团队的分阶段实施路线图
从概念到生产的最简洁路线是分阶段工作流程。
阶段 1 是设计。首先定义目标谱系,然后使用 snRNA-seq 确认主要瓶颈,再选择电路架构。这个步骤很重要,因为电路只能解决您实际识别出的约束。
阶段 2 是构建和二维验证。构建构造体并使用简单的报告器读数检查电路在二维中是否按预期运行。在这个阶段,目标很简单:确认逻辑有效,然后再进入更复杂和更昂贵的模型。
阶段3是与规模相关的压力测试。转向3D系统和生物反应器相关条件,然后将输出与2D基线进行比较。这是许多设计开始显示其弱点的地方,尤其是在质量传递、剪切和基质效应发挥作用时。
阶段4是法规和安全集成,它应与阶段3并行进行。安全和法规工作不应等到最后。与规模化同时进行,包括任何可诱导安全模块的文档。
5.2 通过Cellbase 采购支持工具和材料
一旦工作流程确定,采购通常成为限制步骤。
- 细胞系
- 无血清和化学定义的培养基
- 支架
- 生物反应器组件
- 传感器
- 分析设备
在每个阶段可靠地获取兼容材料直接影响到在规模相关条件下电路行为的表征速度。
5.3 关键要点
合成基因电路为培养肉团队提供了可编程的控制,能够超越仅靠培养基协议的时间、阈值和谱系平衡。架构选择影响可逆性、泄漏性和安全性。诱导系统通常更受欢迎,因为它们提供条件控制和较低的代谢负担[6].
“合成生物学工具包可用于建立具有可调基因表达的细胞系,当与PAT和计算建模结合时,可以实现闭环控制系统,以提供最佳产品产量和质量。” - npj Systems Biology and Applications [6]
成功的部署不仅仅是生物学问题。它依赖于电路工程、生物工艺设计、法规文件和采购之间的紧密结合。
常见问题
合成基因电路如何改善分化一致性?
合成基因电路可以使分化更加一致,因为它们为您提供了对细胞行为和谱系承诺的编程控制。实际上,这意味着使用模块化逻辑操作来调整基因和转录因子的表达,并具有严格的时间控制。
这种时间控制很重要。它帮助细胞按照正确的顺序经历定义的状态变化,而不是漂移到混合或不需要的命运中。它还减少了非目标分化并降低了整个培养过程中的噪音。
回报是显而易见的:更均匀、稳定和成熟的细胞群体用于培养肉生产。
哪种电路设计适合肌源性或脂肪源性控制?
在培养肉研究中,相同的鸡成纤维细胞可以被推动到任一谱系。肌生成遵循一套诱导协议,而脂肪生成可以通过将细胞暴露于鸡血清或脂肪酸等输入来开启。
从那里,这些细胞命运可以在3D水凝胶支架内逐步控制,以构建具有特定脂肪和胶原比例的肉结构。
为什么基因电路在3D培养中经常表现不同?
在3D培养 , 中,基因电路经常表现不同,因为细胞正在处理在2D单层中根本不存在的物理和结构输入。这些输入包括机械张力、剪切应力、基质刚度和局部细胞密度。
这些信号可以改变Notch等信号通路。它们还可以改变合成电路如何检测力并协调下游反应,包括细胞-细胞粘附和组织形态发生。









