

الصمت الجيني فوق الجيني يغير كيفية تعاملنا مع إنتاج اللحوم المزروعة. بالنسبة لمتخصصي R&D، فإنه يوفر طريقة للتحكم في التعبير الجيني دون تغيير دائم في الحمض النووي، مما يعالج التحديات الرئيسية مثل تكاثر الخلايا، التمايز، ومراقبة الجودة. إليك ما تحتاج إلى معرفته:
- ما هو: قمع نشاط الجينات عبر مثيلة الحمض النووي، تعديل الهيستون، أو تداخل الحمض النووي الريبي - طرق قابلة للعكس ودقيقة تترك التسلسل الجيني سليماً.
- لماذا هو مهم: يمدد عمر الخلايا، يعزز تمايز خلايا العضلات، ويحسن القابلية للتوسع مع تجنب المخاطر مثل التسرطن من تعديلات الجينات الدائمة.
- الأدوات الرئيسية: أنظمة CRISPR-dCas9 (مثل KRAB أو DNMT3A) والمحررات القائمة على TALE تحقق كفاءة عالية في الصمت، مع بعض التأثيرات التي تستمر لأكثر من 300 يوم.
- التحديات: تظل تقديم هذه الأدوات على نطاق واسع، خاصة في المفاعلات الحيوية، وتخصيص الأساليب لمسارات الأنواع المحددة عقبات.
بالنسبة لمهندسي العمليات الحيوية وعلماء زراعة الخلايا، يتركز الاهتمام على التحكم الدقيق في سلوك الخلايا لتحسين الإنتاجية وجودة المنتج. قد يكون كبت الجينات اللاجيني هو المفتاح للتغلب على عنق الزجاجة في إنتاج اللحوم المزروعة.
الآليات الأساسية لكبت الجينات اللاجيني في خلايا الماشية
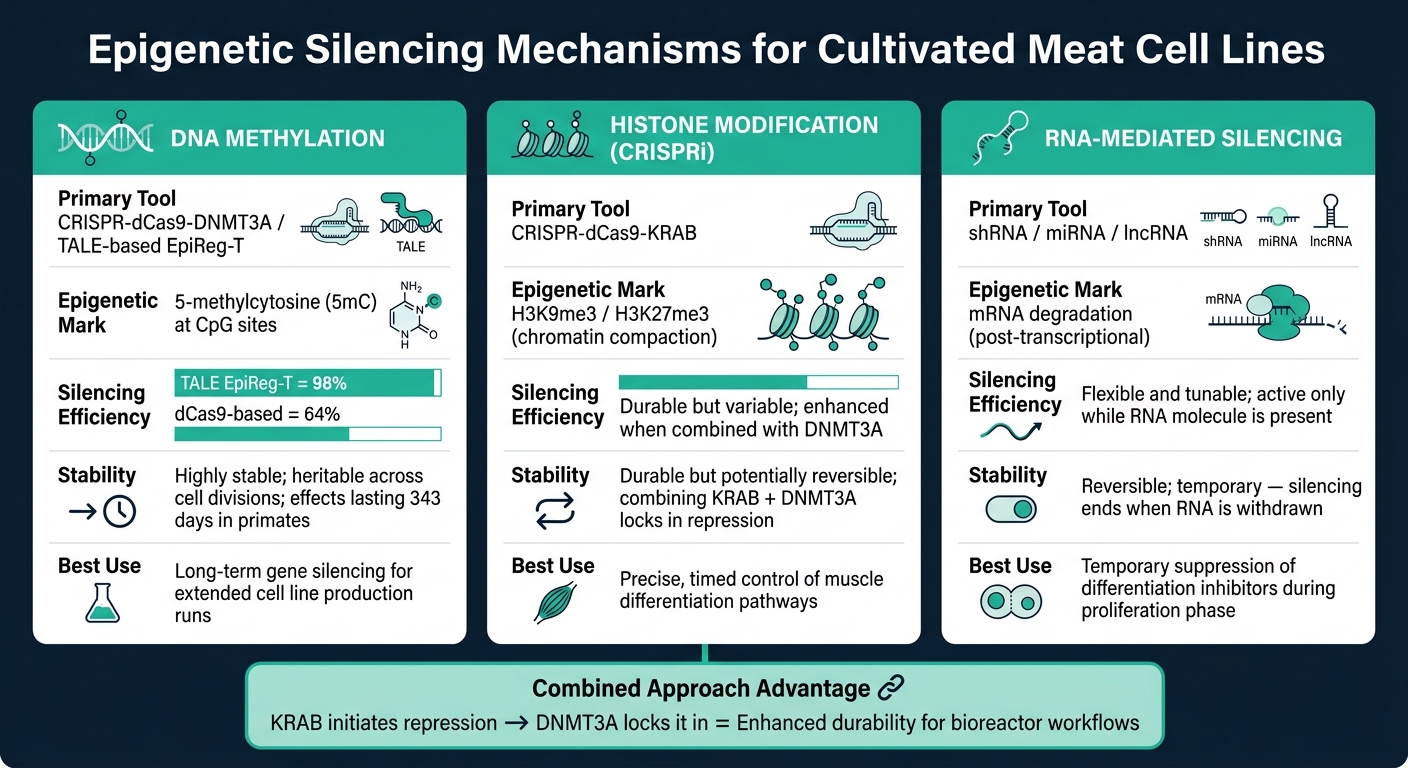
أدوات كبت الجينات اللاجيني للحوم المزروعة: الآليات، الكفاءة & الاستقرار
يعتمد تحسين أداء خطوط خلايا اللحوم المزروعة بشكل كبير على التحكم الدقيق في الآليات اللاجينية. فيما يلي نظرة عامة على الطرق الأساسية المستخدمة في خلايا الماشية.
إسكات قائم على مثيلة الحمض النووي
تشمل مثيلة الحمض النووي إضافة مجموعات الميثيل إلى مواقع CpG، وهي عملية يقودها إنزيمات مثيلة الحمض النووي (DNMTs). عندما يحدث هذا في مناطق محفزات الجينات، فإنه يمنع آلية النسخ من الوصول إلى الجين، مما يؤدي إلى إيقافه فعليًا [6]. هذا الإسكات وراثي، حيث يحافظ DNMT1 على نمط المثيلة عبر انقسامات الخلايا [7].
أداة متقدمة واحدة، CRISPR-dCas9-DNMT3A, تجمع بين بروتين dCas9 غير النشط تحفيزيًا مع إنزيم DNMT3A لتوجيه المثيلة إلى مواقع جينومية محددة. تحقق هذه الطريقة كفاءة إسكات عالية دون قطع الحمض النووي. نهج أكثر دقة، منظمات الجينات القائمة على TALE (EpiReg-T), أظهرت كفاءة إسكات بنسبة 98% في الفئران، مقارنة بـ 64% في الأنظمة السابقة القائمة على dCas9 [5]. في الدراسات التي أجريت على الرئيسيات غير البشرية، حافظت جرعة واحدة من هذا النظام على إسكات الجينات لمدة تصل إلى 343 يومًا [5].
بعد تأسيس مثيلة الحمض النووي، توفر تعديلات الهيستون طبقة ثانوية وديناميكية لتنظيم الجينات.
تعديل الهيستون وCRISPRi
تعديلات الهيستون تغير بنية الكروماتين عن طريق استهداف بروتينات الهيستون، مما يجعل الجينات أكثر أو أقل وصولاً. العلامات مثل H3K9me3 و H3K27me3 تضغط الكروماتين، مما يمنع عوامل النسخ من الوصول إلى الحمض النووي [6].
تداخل CRISPR (CRISPRi) يستخدم dCas9 مدمجًا مع مجال مثبط KRAB. يتم توجيه هذا المركب إلى محفزات جينية محددة، حيث يجند بروتينات مثبطة تودع علامات هيستون مثبطة.أظهرت الأبحاث في الأغنام أن H3K27me3 يعد إشارة قمعية رئيسية خلال تطوير العضلات، بينما ترتبط المحفزات النشطة بالجينات التي تعزز الأداء النموذجي المتفوق [8]. من خلال فهم حالات الهيستون التي تنظم تمايز العضلات في الماشية، يمكن للعلماء ضبط سلوك الخلايا بدقة.
"يعتبر التحرير الجيني استراتيجية واعدة لتعديل التعبير الجيني مع تجنب التغييرات الدائمة والسمية الجينية المحتملة لتقنيات تحرير الجينوم." - Nature Biotechnology [5]
تعد تعديلات الهيستون أكثر ديناميكية من مثيلة الحمض النووي، مما يتطلب تدخلات مستمرة أو موقوتة للحفاظ على تأثيراتها. يمكن أن يعزز الجمع بين KRAB وDNMT3A في بنية واحدة من المتانة: حيث تبدأ علامات الهيستون في القمع، بينما تقوم المثيلة بتثبيته [5].
بالإضافة إلى هذه الأساليب القائمة على الحمض النووي، يوفر إسكات الحمض النووي الريبي بديلاً مرناً ومؤقتاً.
الإسكات بوساطة الحمض النووي الريبي
يركز الإسكات بوساطة الحمض النووي الريبي على تقليل مستويات mRNA مباشرة. الميكرو RNA (miRNAs) و الشعرية القصيرة RNA (shRNAs) ترتبط بتسلسلات mRNA المكملة، مما يؤدي إلى تحللها قبل الترجمة [6]. وفي الوقت نفسه، الحمض النووي الريبي الطويل غير المشفر (lncRNAs) يعمل في وقت مبكر عن طريق تجنيد مجمعات تعديل الكروماتين إلى مناطق جينومية محددة [6].
بالنسبة لتطبيقات اللحوم المزروعة، يوفر الإسكات بوساطة الحمض النووي الريبي ميزة كبيرة: القابلية للعكس والمرونة. يبقى الإسكات نشطاً فقط بينما تكون جزيئات الحمض النووي الريبي موجودة، مما يجعله مثالياً للتدخلات المؤقتة. على سبيل المثال، يمكن قمع مثبطات التمايز خلال مرحلة التكاثر، ثم إزالتها للسماح بتطور العضلات الطبيعي.ومع ذلك، فإن الحفاظ على التسليم المستمر لجزيئات RNA يمكن أن يضيف تعقيدًا عند توسيع خطوط الخلايا لزراعة المفاعلات الحيوية.
يلخص الجدول أدناه الميزات الرئيسية لهذه الآليات:
| الآلية | الأداة الرئيسية | العلامة الجينية التخلقية | الاستقرار |
|---|---|---|---|
| مثيلة الحمض النووي | CRISPR-dCas9-DNMT3A | 5-ميثيل سيتوزين (5mC) | مستقر للغاية؛ وراثي عبر الانقسامات[5][7] |
| قمع الهستون (CRISPRi) | CRISPR-dCas9-KRAB | H3K9me3 / H3K27me3 | دائم ولكن يمكن عكسه[5][8] |
| تداخل الحمض النووي الريبي | shRNA / miRNA | تحلل mRNA | قابل للعكس وقابل للضبط[6] |
sbb-itb-ffee270
الأهداف الجينية والمسارات لتحسين أداء خطوط الخلايا
بناءً على المناقشات السابقة حول الآليات الجينية اللاجينية، فإن اختيار الأهداف الجينية الصحيحة أمر حاسم لتحسين أداء خطوط الخلايا.يعتمد نجاح هذه التدخلات ليس فقط على تحديد الأهداف ولكن أيضًا على اختيار طرق الإسكات المناسبة. حددت الأبحاث مجموعة أساسية من الأهداف الجينية التي، عند قمعها، تعزز الجوانب الرئيسية لخطوط خلايا اللحوم المزروعة، بما في ذلك التكاثر، التمايز، والاستقرار الأيضي.
التكاثر والخلود
غالبًا ما يتضمن تحسين القدرة التكاثرية استهداف جينات مثل CDKN2A و TP53 . CDKN2A يرمز إلى p16^INK4A و p14^ARF، وهما بروتينات تحد من دورة الخلية وتدفع إلى الشيخوخة. يمنع إسكات CDKN2A توقف G1/S، مما يتيح توسع الخلايا بشكل قوي. على سبيل المثال، حافظت الخلايا الخنزيرية مع إسكات CDKN2A على خصائصها العضلية عبر 18-30 تمريرة، بينما فقدت الخلايا البرية هذه الخصائص بحلول التمريرة 10.علاوة على ذلك، تسبب استنفاد CDKN2A في زيادة تعبير PAX7 بحوالي 194 ضعفًا في الممر 20، وهو عامل حاسم لهوية الخلايا الجذعية العضلية [9] .
"استهداف موقع جين CDKN2A ضروري لمنع الشيخوخة أو تحفيز خلود الخلايا." - أبحاث مواد الغذاء [9]
TP53 هو هدف رئيسي آخر. حددت شاشة CRISPR لـ 600 جين في الخلايا الجذعية الميزنشيمية البقرية TP53 كأكثر الأهداف فعالية لتعزيز التكاثر. أدى تعطيل TP53 إلى زيادة وفرة الخلايا بمقدار 1,000 ضعف خلال 30 يومًا، مع أداء متسق في التوسع طويل الأمد [1] . بالإضافة إلى ذلك، فإن إسكات PTEN, وهو منظم سلبي لمسار PI3K/AKT/mTOR، يعزز معدلات مضاعفة الخلايا ونشاط mTOR.ومع ذلك، يتطلب هذا النهج مراقبة دقيقة، حيث قد يقلل من كفاءة التمايز [1].
تضع هذه التطورات في التكاثر الأساس لتحسين التمايز، وهو الخطوة الحاسمة التالية.
التحكم في التمايز
موازنة توسع الخلايا مع تكوين الأنسجة هو تحدٍ معقد في إنتاج اللحوم المزروعة. أحد الأهداف المدروسة جيدًا هو الميوستاتين (MSTN), وهو منظم سلبي لتكوين العضلات. يؤدي إسكات MSTN إلى تعزيز تكوين ألياف العضلات، مشابهًا لصفة "العضلات المزدوجة" في بعض سلالات الماشية [4] . عند الجمع بين MYOD1 التنشيط والتقنيات المتقدمة مثل معالجة الضوء الرقمي (DLP) الطباعة الحيوية ثلاثية الأبعاد على الهلاميات المائية ذات الأنماط المحززة، يتم تحسين محاذاة الخلايا العضلية وتمايزها بشكل كبير من خلال التوظيف السطحي [4] .
جانب حاسم آخر هو إدارة منظمات التعددية مثل SOX2 و OCT4 . الصمت العكسي لـ SOX2 باستخدام منصات CRISPR/dCas9-KRAB يحقق قمعًا يصل إلى 85% خلال 72 ساعة، مع استعادة التعبير الأساسي إلى حوالي 90% بعد سحب البناء التحريري [3] . تسمح هذه القابلية للعكس بالقمع المتحكم فيه أثناء توسع الخلايا والإفراج في الوقت المناسب لدعم التطور السليم للأنسجة.
مسارات الإجهاد والتمثيل الغذائي
يتضمن الحفاظ على جودة الخلايا على مدى دورات الإنتاج الممتدة معالجة تحديات الإجهاد والتمثيل الغذائي. TP53 يلعب دورًا مزدوجًا كمثبط للأورام وكحساس للإجهاد. تحت ظروف الزراعة، يمكن أن يحفز الشيخوخة المبكرة، حتى بدون تلف جيني كبير [1] . من خلال إسكات TP53, تحتفظ الخلايا بملفات التعبير الجيني للخلايا في المراحل المبكرة، مما يحافظ على الوظائف الحيوية مثل تخليق البروتين وتكرار الحمض النووي [1].
يلخص الجدول أدناه الأهداف الجينية الأساسية وأدوارها الوظيفية:
| الجين المستهدف | المسار | تأثير الإسكات | سياق الأنواع |
|---|---|---|---|
| CDKN2A | قمع دورة الخلية | يمنع الشيخوخة؛ ~194× PAX7 زيادة في التعبير عند المرور 20 [9] | الخنزير |
| TP53 | استجابة الإجهاد / مثبط الورم | زيادة 1,000× في وفرة الخلايا على مدى 30 يومًا؛ توسع طويل الأمد متسق [1] | البقري |
| PTEN | PI3K/AKT/mTOR | يزيد من معدل التضاعف؛ يعزز نشاط mTOR [1] | البقري |
| MSTN | تنظيم تكوين العضلات | يعزز تكوين ألياف العضلات وكفاءة التمايز [4] | بقري |
| SOX2 | الحفاظ على التعددية القدرات | يدير الانتقال من الجذعية إلى التمايز؛ 85% قمع في 72 ساعة [3] | متعدد |
نهج واعد يكتسب زخماً هو الاستهداف المتعدد، والذي يتضمن إسكات جينات متعددة في وقت واحد. على سبيل المثال، الجمع بين قمع CDKN2A وتفعيل GATA4 أظهر تأثيرات تآزرية تتفوق على التدخلات الفردية [9] [10]. تسلط هذه الاستراتيجية على مستوى الأنظمة الضوء على أهمية المنصات المتخصصة مثل
أدوات الإبيجينيتك وطرق التوصيل
للاستفادة من أهداف الجينات المحددة، يعتمد الباحثون على أدوات إبيجينيتك متخصصة وأنظمة توصيل فعالة.
منصات الإبيجينيتك الاصطناعية
تحديد أهداف الجينات الصحيحة هو جزء فقط من المعادلة - الأدوات المستخدمة لإسكات هذه الجينات هي بنفس الأهمية. يبرز نظامان قابلان للبرمجة لأهميتهما في أبحاث اللحوم المزروعة: CRISPRoff و منظمات الإبيجينيتك القائمة على TALE (EpiReg-T).
CRISPRoff يستخدم هيكل dCas9 مدمجًا مع مجالات KRAB وDNMT3A/3L لإنشاء علامات قمعية موروثة، مثل مثيلة الحمض النووي وH3K9me3، دون إدخال كسور في الحمض النووي. يضمن هذا النهج إسكات الجينات بشكل مستمر، مما يجعله مفيدًا بشكل خاص للحفاظ على خطوط الخلايا لفترات طويلة - وهو عامل رئيسي في معالجة تحديات القابلية للتوسع والاتساق في إنتاج اللحوم المزروعة. في المقابل، أظهر EpiReg-T القائم على TALE كفاءة إسكات فائقة، حيث حقق 98% مقارنة بـ 64% التي شوهدت مع أنظمة مماثلة تعتمد على dCas9 [5].
أبرزت دراسة محورية نُشرت في Nature Biotechnology في أكتوبر 2025 إمكانات المحررين القائمين على TALE.باحثون، بما في ذلك من Epigenic Therapeutics و الأكاديمية الصينية للعلوم , أظهروا أن جرعة واحدة من EpiReg-T تم توصيلها عبر الجسيمات النانوية الدهنية (LNPs) قامت بإسكات جين PCSK9 في قرود المكاك بكفاءة تزيد عن 90% لمدة 343 يومًا. تم تحقيق ذلك مع تأثيرات جانبية طفيفة، كما تم تأكيده من خلال تحليلات متعددة الأوميكس [5]. تضع هذه النتائج الأنظمة القائمة على TALE في مكانة متميزة عندما تكون المتانة والفعالية أمرًا حاسمًا.
تحديات التوصيل
يبقى توصيل هذه الأدوات بفعالية إلى خلايا الماشية - خاصة على نطاق واسع - تحديًا تقنيًا كبيرًا. بينما يتجنب المحررون الجينيون مخاطر انكسار الحمض النووي المزدوج، إلا أنهم لا يزالون بحاجة إلى آلية توصيل موثوقة. الجسيمات النانوية الدهنية (LNPs) برزت كخيار غير فيروسي رائد.إنهم يقدمون بشكل عابر mRNA الذي يشفر المحرر الجيني، مما يمكن من اتباع نهج "الضرب والهرب" الذي يؤسس لإسكات الجينات بشكل دائم دون دمج الحمض النووي [5]. هذه الطبيعة العابرة مهمة بشكل خاص للحوم المزروعة، حيث تظل المخاوف التنظيمية حول التعديلات الجينية قضية رئيسية.
ومع ذلك، يمكن أن تختلف كفاءة LNP بشكل كبير اعتمادًا على نوع الخلية. لا يزال تحسين التركيبات لخلايا البقرية الأولية أو خلايا الأقمار الصناعية العضلية للخنازير، خاصة في إعدادات مفاعل حيوي على نطاق واسع، مجالًا نشطًا للبحث. غالبًا ما تفشل طرق التوصيل التي تعمل بشكل جيد في التجارب الصغيرة في الأداء بشكل متسق في المفاعلات الحيوية ذات الخزانات المقلوبة. حل هذه التحديات في التوصيل ضروري لتقدم البحث وتوسيع الإنتاج، وهي عملية مدعومة بشكل متزايد من قبل منصات متخصصة.
كيف Cellbase يدعم التعديلات الجينية &R
تتطلب خطوط الخلايا المعدلة جينياً أدوات دقيقة التحقق. يحتاج الباحثون إلى الوصول إلى خطوط خلايا موصوفة جيداً ومتوافقة مع التعديلات الجينية، وصيغ وسائط محددة تحافظ على الاستقرار الجيني، وأدوات تحليلية قادرة على تأكيد إسكات الجينات على مستوى الكروماتين. غالباً ما تفتقر موردي المختبرات العامة إلى الخبرة لضمان التوافق مع تطبيقات اللحوم المزروعة.
ما الذي يعنيه إسكات الجينات اللاجينية لمعالجة اللحوم المزروعة
تحسينات قابلة للقياس في خطوط الخلايا
يقدم إسكات الجينات اللاجينية مزايا عملية أصبحت واضحة بشكل متزايد، خاصة في تمديد العمر الإنتاجي لخطوط الخلايا. من خلال استخدام استراتيجية مؤقتة "اضرب واهرب"، يمكن للباحثين قمع الجينات المسؤولة عن الشيخوخة مؤقتًا دون تعديل دائم للجينوم [2]. لقد أظهرت هذه الطريقة نجاحًا في الخلايا العضلية للأبقار و الخلايا العضلية للخنازير, مما يتيح مضاعفات خلوية أكبر بكثير ومعالجة الاختناقات الشائعة في معالجة العمليات الحيوية. الأهم من ذلك، أن هذه الطريقة قابلة للعكس - بمجرد سحب التركيب، تعود تعبيرات الجينات تقريبًا إلى مستوياتها الأساسية [3] . هذا التحكم القابل للعكس مثالي لعمليات سير العمل في المفاعلات الحيوية، حيث يضمن استمرار تكاثر الخلايا خلال مرحلة التوسع ويسمح بتحفيز التمايز في الوقت المناسب. يؤدي التوسع المحسن للخلايا مباشرة إلى تمايز الأنسجة بشكل أكثر كفاءة وتحسين جودة المنتج.
تكوين الأنسجة وجودة المنتج
تشكل الزيادات في تكاثر الخلايا الأساس لتكوين أنسجة أفضل. التمايز المتحكم فيه هو المكان الذي يؤثر فيه الإسكات الجيني بشكل مباشر على جودة المنتج النهائي. على سبيل المثال، في إعادة برمجة خلايا الأبقار، يسهل إسكات علامات تعدد القدرات مثل OCT4, SOX2, و NANOG الانتقال إلى السلالة العضلية.تؤدي هذه العملية إلى تكوين أنابيب عضلية طويلة ومتعددة النوى بحلول اليوم 30 من بروتوكول التمايز [11] .
"إسكات mOSKM وعلامات تعدد القدرات... أمر حاسم للانتقال من تعدد القدرات إلى السلالة العضلية." - Frontiers in Nutrition [11]
إلى جانب تطوير ألياف العضلات، يلعب التحكم الدقيق في المسارات التمايزية للخلايا الدهنية دورًا حاسمًا في تحقيق الترخيم. الترخيم هو عامل رئيسي يؤثر على كل من النكهة والإحساس في الفم، ويمكن تحقيق هذه التحسينات دون إجراء تغييرات دائمة على الجينوم.
الاعتبارات التنظيمية والمستهلكين
تجلب التطورات في تكاثر الخلايا وتكوين الأنسجة أيضًا وجهات النظر التنظيمية والمستهلكين إلى التركيز.تدعم الهيئات التنظيمية عمومًا إسكات الجينات اللاجينية بسبب تأثيره غير الدائم على الجينوم. أدوات مثل dCas9-KRAB وEpiReg-T المستندة إلى TALE تتجنب المخاطر المرتبطة بانكسارات الحمض النووي مزدوجة السلسلة، مما يجعلها مناسبة لخطوط الخلايا الغذائية التي يجب أن تُظهر استقرارًا جينيًا طوال الإنتاج [5].
ومع ذلك، يظل الحفاظ على حالة خالية من الجينات المنقولة تحديًا. دراسة نُشرت في مايو 2025 من قبل باحثين من جامعة ساو باولو وجامعة كوبنهاغن, بما في ذلك كايانا ريكيا وكريستين فرويد، استكشفت هذه القضية. قاموا بإعادة برمجة الخلايا الليفية الجنينية البقرية باستخدام نواقل إبزومية غير مدمجة، ووجدوا أنه بينما ظلت المستعمرات مستقرة لأكثر من 33 تمريرة، كانت البلازميدات الإبزومية لا تزال قابلة للكشف في التمريرات 12 و17 [11] .
من جانب المستهلك، الشفافية حول الأساليب المستخدمة أمر بالغ الأهمية.التواصل الواضح بأن إسكات الجينات اللاجينية لا يغير الحمض النووي بشكل دائم سيكون مفتاحًا لبناء الثقة العامة مع اقتراب منتجات اللحوم المزروعة من التسويق التجاري.
الاتجاهات المستقبلية والفجوات البحثية
التحديات الخاصة بالأنواع
أحد أكبر العقبات في هذا المجال هو نقص الفهم التفصيلي لمسارات التكوين العضلي في أنواع الماشية. بينما تم توثيق مسارات مثل IGF-1 وMAPK/Erk وWnt/β-catenin بشكل جيد في البشر والفئران، فإن أدوارها في الأبقار والخنازير مفهومة جزئيًا فقط [11]. بدون خريطة كاملة، يصبح تحديد أهداف الجينات المحددة لإسكات الجينات اللاجينية تحديًا كبيرًا.
يضيف تكوين ألياف العضلات طبقة أخرى من التعقيد. على سبيل المثال، تحتوي عضلة Longissimus في الخنازير على حوالي 55% من الألياف السريعة الانقباض من النوع IIb، لكن هذه الألياف غائبة في أنواع مثل الأغنام والخيول.عند دمج هذا مع تعبير الجينات HOX الخاص بالمنطقة، يصبح من الواضح أن استراتيجيات الإسكات تحتاج إلى أن تكون مخصصة لكل نوع [13] . الخلايا الساتلية، التي تحتفظ بتعبير الجينات HOX الموضعي (e.g. ، HOXA11 و HOXA13 في عضلات الأطراف الخلفية)، تزيد من تعقيد الأمور. يمكن أن تؤثر هذه الأنماط على ما إذا كانت الخلايا تميل أكثر نحو التكاثر السريع أو التمايز القوي [14] .
"لأن الخلايا الساتلية يمكن أن تحتفظ بهذه التوقيعات الموضعية، قد تختلف قدراتها التكاثرية والتمييزية حسب العضلة الأصلية." - npj Science of Food [14]
من الناحية العملية، يعني هذا أن الباحثين يجب أن يفحصوا خطوط الخلايا لتعبير الجينات HOX قبل تطبيق الإسكات الجيني.يمكن أن تعمل هذه التواقيع الجينية كرموز شريطية بيولوجية، مما يساعد على التحقق من الهوية الإقليمية للخلايا ومواءمتها مع الخصائص المرغوبة للمنتج النهائي.
تسلط هذه التحديات الخاصة بالأنواع الضوء على أهمية النظر في مصادر الخلايا البديلة، مثل الخلايا الجذعية المحفزة (iPSCs)، في تطوير استراتيجيات بنوك الخلايا.
روابط لتطوير الخلايا الجذعية المحفزة وبنوك الخلايا
الخلايا الجذعية المحفزة (iPSCs) تقدم بديلاً واعداً للخلايا الساتلية، التي تكون عرضة للشيخوخة وتتطلب خزعات متكررة. في مايو 2025، نجح الباحثون من جامعة ساو باولو وجامعة كوبنهاغن - بما في ذلك كايانا ريكيا وكريستين فرويد - في تطوير خطوط خلايا iPSC البقرية باستخدام نواقل إبزومية غير مدمجة. حافظت هذه الخلايا على الاستقرار لأكثر من 33 مروراً وتمايزت إلى أنابيب عضلية متعددة النوى بحلول اليوم 30 [11]. ومع ذلك، فإن تأكيد خلوها من الجينات المحورة من خلال تفاعل البوليميراز المتسلسل الجينومي الصارم يبقى خطوة حاسمة.
قضية ذات صلة هي الذاكرة الجينية اللاجينية. غالبًا ما تحتفظ الخلايا الجذعية المحفزة متعددة القدرات (iPSCs) بآثار من نسيجها الجسدي الأصلي، مما يمكن أن يحرف التمايز بعيدًا عن السلالة المقصودة [12]. بالنسبة لبنوك الخلايا، من الضروري اختيار أنسجة المتبرعين ذات الملفات الجينية اللاجينية الموجهة بالفعل نحو تكوين العضلات أو الدهون. بالإضافة إلى ذلك، فإن ضمان إسكات فعال لمؤشرات التعددية المتبقية أمر حيوي لإنشاء بنوك خلايا موثوقة وطويلة الأمد.
كما يؤكد تطوير بروتوكولات iPSC القوية على الحاجة إلى اختبارات موحدة وممارسات مشاركة بيانات متسقة عبر الجهود البحثية.
التوحيد والبيانات المفقودة
للاستفادة الكاملة من إمكانات التدخلات اللاجينية في اللحوم المزروعة، يجب معالجة قضايا التوحيد.حاليًا، لا يوجد إطار عمل عالمي لمراقبة الاستقرار الجيني خلال عمليات الانقسام الخلوي المكثفة المطلوبة للإنتاج على نطاق صناعي [12]. بدون طرق موحدة، يصبح من الصعب مقارنة النتائج عبر المختبرات، وغالبًا ما تعتمد القرارات المتعلقة بتوسيع الإنتاج على بيانات غير مكتملة.
يمكن أن تساعد خطوات عملية في معالجة هذه الفجوة. على سبيل المثال، اعتماد بروتوكولات تنقية FACS متسقة - تستهدف علامات مثل CD31⁻/CD45⁻/CD29⁺/CD56⁺ - سيجعل مجموعات الخلايا الساتلية أكثر قابلية للمقارنة عبر الأنواع والمصادر التشريحية [14]. التحول من الوسائط القائمة على المصل إلى وسائط محددة كيميائيًا يمكن أن يقلل أيضًا من التباين بين الدفعات، مما يخلق بيئات جينية أكثر اتساقًا [12].
بالنظر إلى المستقبل، يمكن أن يغير دمج النمذجة الحاسوبية المدفوعة بالذكاء الاصطناعي قواعد اللعبة في تحسين بروتوكولات الجينات.ومع ذلك، لكي تكون هذه النماذج فعالة، فإن توحيد البيانات عبر مجتمع أبحاث اللحوم المزروعة أمر ضروري. ستُمكّن ممارسات مشاركة البيانات الموحدة الباحثين من التنبؤ بنتائج التلاعبات الجينية بشكل أكثر دقة، مما يسرع التقدم في هذا المجال.
الأسئلة الشائعة
كيف يختلف إسكات الجينات اللاجيني عن تحرير الجينات الدائم في خلايا اللحوم المزروعة؟
ينظم إسكات الجينات اللاجيني نشاط الجينات دون إجراء تغييرات دائمة على تسلسل الحمض النووي، على عكس تحرير الجينات الذي يتضمن تعديل الجينوم فعليًا. نظرًا لأن الأساليب اللاجينية لا تتضمن كسر أو تعديل الحمض النووي، فإنها تُعتبر غالبًا خيارات أكثر أمانًا للاستخدام في إنتاج اللحوم المزروعة. توفر تقنيات مثل الأدوات القائمة على CRISPR ميزة تنظيم الجينات المرن وفي بعض الحالات القابل للعكس.بالنسبة للباحثين الذين يعملون بهذه الأساليب،
ما هي الجينات التي يجب إسكاتها أولاً لتعزيز التكاثر دون الإضرار بالتمايز؟
لتشجيع تكاثر الخلايا مع الحفاظ على قدرتها على التمايز، من الضروري إسكات الجينات التي إما تعيق دورة الخلية أو تؤدي إلى مصائر خلوية غير مرغوب فيها. على سبيل المثال، تم إثبات أن قمع CDKN2A يزيد بشكل ملحوظ من التكاثر في خلايا الأقمار الصناعية الخنزيرية دون التأثير على قدرتها على التمايز. وبالمثل، يمكن أن يؤدي استهداف جينات مثبطة الأورام مثل TP53 وPTEN إلى تعزيز النمو، على الرغم من أن هذه التدخلات تتطلب إشرافًا دقيقًا.
كيف يمكن توصيل المحررين الجينيين بشكل موثوق على نطاق مفاعل حيوي؟
تقديم المحررين الجينيين على نطاق واسع لإنتاج اللحوم المزروعة يمثل تحديًا كبيرًا. يعود ذلك بشكل كبير إلى الحجم الكبير لأدوات CRISPR والقيود المفروضة على طرق التوصيل التقليدية مثل الكهربة أو النواقل الفيروسية. ومع ذلك، تظهر بعض الاستراتيجيات الواعدة. على سبيل المثال، أنظمة التوصيل المؤقتة باستخدام الجسيمات النانوية الدهنية أو الجسيمات الشبيهة بالفيروسات المهندسة تظهر إمكانيات. يمكن لهذه الطرق تغليف شحنات CRISPR الكبيرة، مما يسمح بدخول فعال إلى الخلايا دون التسبب في تكامل الجينوم. لدعم مثل هذه المبادرات المتقدمة، يوفر









