

إذا كان بإمكانك توسيع الخلايا ولكن لا يمكنك تحويلها إلى المصير الصحيح في الوقت المناسب، فسيتوقف عمليتك عند التمايز. هذه هي النقطة الأساسية هنا: الدوائر الجينية الاصطناعية تمنحك داخل الخلية التحكم في الالتزام، التوقيت، الذاكرة ومزيج السلالة، حيث أن التغييرات في الوسائط وحدها غالبًا ما تترك مجموعات غير متجانسة, جزئيًا ملتزمة.
إذا كنت أبني سير عمل لتمايز اللحوم المزروعة، فسأخذ أربع نقاط من هذه المقالة على الفور:
- ابدأ بالشبكة الأصلية، وليس بالهيكل. استخدم تحليل مسار snRNA-seq, استنتاج GRN وتحليل miRNA لمعرفة أين تتوقف الخلايا، تنحرف أو تتفرع إلى المصير الخاطئ.
-
طابق نوع الدائرة مع مشكلة العملية.
مفتاح التبديل يناسب القفل، تصميم التغذية الأمامية أو تمرير النطاق يناسب التحكم في التوقيت، بوابة المنطق تناسب التحكم في الإشارات المتعددة، و miSFITs تناسب المخرجات المتدرجة. - التصميم لتقليل التسرب، وتقليل الضوضاء وضمان السلامة من اليوم الأول. الأجزاء المتعامدة، التنظيم الذاتي السلبي، iFFLs، الجينات المحورة cm ووحدة القتل أو إيقاف النمو القابلة للتحفيز هي جزء من البناء، وليست فكرة لاحقة.
- التحقق من الصحة في ظل ظروف ذات صلة بالحجم مبكرًا. قد يتغير الدائرة التي تعمل في 2D في 3D، أو حامل دقيق أو تعليق محرك بسبب تدرجات المحفز، حدود الأكسجين والقص.
توضح المقالة أيضًا نقطة عملية تهم فرق العمليات: التحكم في السلالة الواحدة والتحكم في النسبة هما وظائف مختلفة. قد يدفع شريط Tet-On MyoD الدخول العضلي، ولكن المنتجات المقطوعة بالكامل تحتاج إلى التحكم في نسب العضلات، الدهون و ECM, والتي تعني عادةً التغذية الراجعة، الإشارات الباركرينية وفحص النسخ الأثقل.
بعض الأرقام تدعم هذه الرسالة:
- يمكن أن تتوقف التمايز العضلي القياسي مع مؤشرات الاندماج بحوالي 50-60%
- زادت الشبكات الجينية المهندسة في الخلايا الجذعية المستحثة من التمايز المستهدف من 52% إلى 81%
- دفعت الدوائر الاصطناعية في الخلايا الجذعية الوسيطة المعدلة التمايز القلبي إلى 76%
- بعض خطوط Tet-On-PAX7 الخنزيرية حافظت على إمكانات عضلية عالية بعد 40 تمريرة
- حوالي 20% من الخلايا الجذعية البشرية متعددة القدرات قد تحمل طفرات مرتبطة بالسرطان، ولهذا السبب تعتبر وحدات الأمان القابلة للتحفيز مهمة
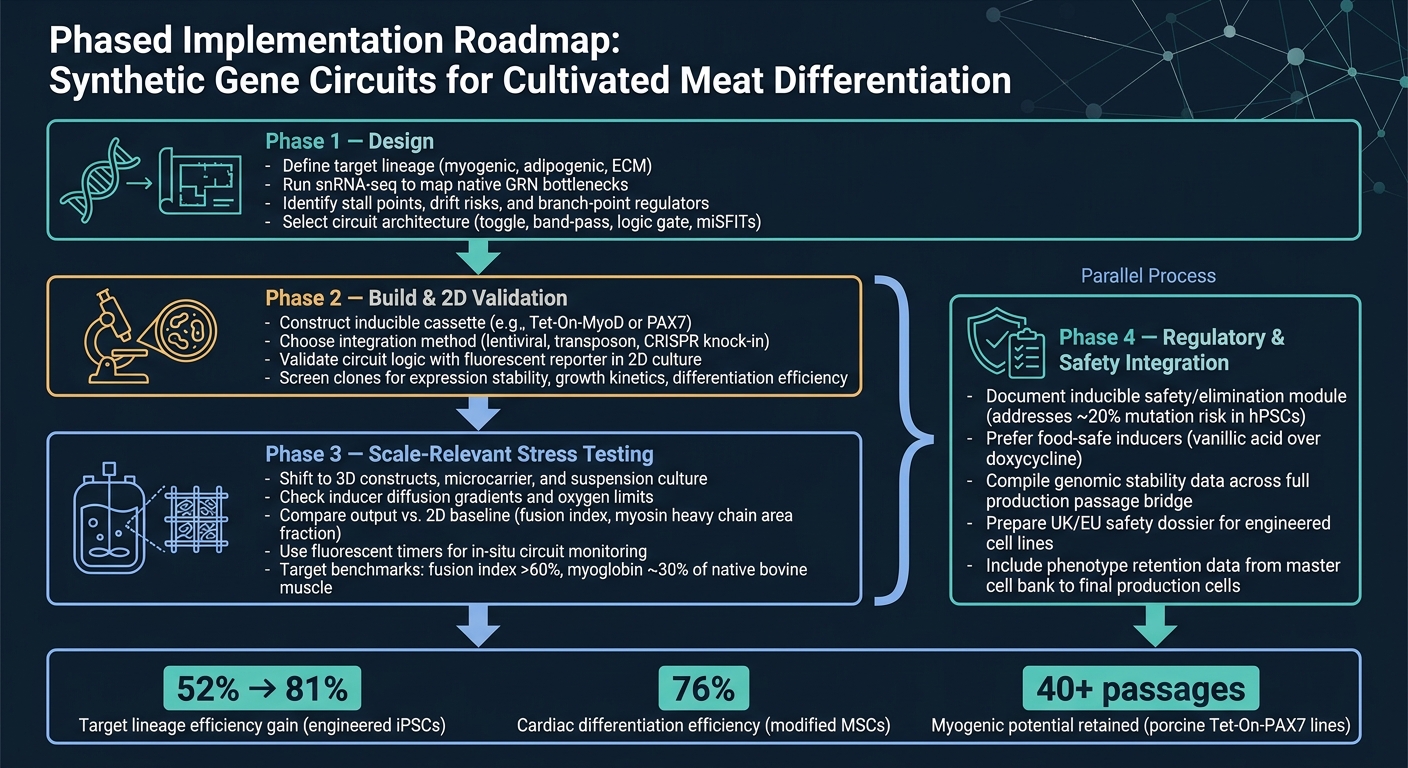
سير عمل الدائرة الجينية الاصطناعية لتمايز اللحوم المزروعة
قصص العلماء: مايكل إلوفيتز، الدوائر الجينية في الخلايا الحية
مقارنة سريعة
| الموضوع | ما يقوله المقال بعبارات بسيطة |
|---|---|
| العنق الرئيسي | التمايز، وليس التوسع |
| لماذا تساعد الدوائر | إنها تضيف التحكم في العتبة وذاكرة حالة الخلية داخل الخلية |
| أفضل خطوة أولى | رسم خريطة التحكم في المصير الطبيعي باستخدام snRNA-seq والأدوات ذات الصلة |
| خيارات الدائرة الرئيسية | التبديل، التغذية الأمامية، تمرير النطاق، بوابات المنطق، miSFITs، CRISPRa/i |
| مثال على تكوين العضلات | Tet-On-MyoD لفصل النمو عن التمايز النهائي |
| تكوين الدهون / التحكم في ECM | miSFITs وتصميمات القوس لخرج متدرج من نوع PPARG/BMP4 |
| تحدي القطع الكامل | التحكم في النسبة عبر العضلات، الدهون والأنسجة الضامة |
| مخاطر التوسع | قد لا يكون السلوك ثنائي الأبعاد صالحًا في الأبعاد الثلاثية أو في المفاعلات الحيوية |
| خيارات التكامل | الفيروسات البطيئة، الترانسبوزونات، CRISPR الإدخال، النواقل الإبيسومية |
| نقطة تنظيمية | تحتاج الخطوط المهندسة إلى حزمة أمان أوسع؛ يفضل استخدام المحفزات الآمنة للأغذية مثل حمض الفانيليك على DOX حيثما أمكن |
بعبارات بسيطة: هذا ليس مجرد مقال عن تصميم الدوائر. سأقرأها كدليل لربط هندسة الدوائر، علم الأنساب البيولوجي، اختيار النسخ، أداء المفاعل الحيوي ووثائق السلامة في المملكة المتحدة/الاتحاد الأوروبي في استراتيجية تمايز واحدة.
اقرأ المزيد إذا كنت تريد المسار الكامل من رسم الشبكة الأصلية إلى اختيار البناء، فحوصات التوسع والملاءمة التنظيمية.
2. مبادئ التصميم لدوائر التحكم في التمايز
2.1 رسم شبكة مصير الخلية الأصلية قبل تصميم الدائرة
قبل تصميم دائرة، تحتاج إلى صورة واضحة لما تفعله الخلية بالفعل.
تسلسل الحمض النووي الريبي أحادي النواة (snRNA-seq) هو مكان جيد للبدء. يمكن أن يظهر مجموعات فرعية خاملة، بما في ذلك الخلايا الاحتياطية التي تم تحديدها بواسطة NOTCH2 و HEYL , ويشير إلى أهداف المسار التي قد تحسن التمايز [3].
من هناك، يساعد تحليل المسار واستنتاج شبكة تنظيم الجينات (GRN) في رسم ترتيب تفعيل المنظمين وتسليط الضوء على الأماكن التي من المرجح أن تتوقف فيها الخلايا. في تكوين العضلات، يمر الشلال الرئيسي عبر MYOD1 و MYOG . في تكوين الدهون، العقد الرئيسية هي PPARG و CEBPA , مع مصير الخلايا الجذعية الليفية الدهنية (FAP) الذي يعمل كنقطة تفرع رئيسية. يلخص الجدول أدناه المنظمين الرئيسيين والعوائق.
| النسب | منظمو المفاتيح الرئيسيون | مسارات الإشارة الحرجة | عنق الزجاجة المحددة |
|---|---|---|---|
| العضلي المنشأ | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | تكوين الخلايا الاحتياطية (السكون) |
| الشحمي المنشأ | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | مصير السلف الليفي الشحمي (FAP) |
| متعدد القدرات | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | التمايز العفوي / التباين |
طبقة مفيدة أخرى هي توصيف تعبير miRNA. miRNAs الذاتية مثل miR-302a, مرتبطة بتعدد القدرات، و miR-375, مرتبطة بالتمايز، يمكن أن تعمل كمصنفات داخلية في تصميمات الاستشعار والاستجابة.هذا يسمح للدائرة بقراءة الحالة الفعلية للخلية بدلاً من الاعتماد فقط على المحفزات الخارجية [5].
يجب أن تدفع هذه الاختناقات لاختيار الدائرة. إذا كانت المشكلة الرئيسية هي الانجراف، فقد تحتاج إلى قفل. إذا كانت المشكلة في التوقيت، فقد يكون النبض مناسبًا أكثر. إذا كان التحكم في المصير يعتمد على أكثر من إشارة واحدة، فإن المنطق متعدد المدخلات يكون أكثر منطقية عادةً.
2.2 اختر بنية الدائرة المناسبة
هنا تظهر المقايضات بوضوح. تعتمد البنية الصحيحة على ثلاث نقاط عملية: مدى ديمومة الالتزام، مدى الحاجة إلى التحكم في التوقيت بدقة، وكمية الحمولة الجينية التي يمكن للخلية حملها دون مشاكل.
مفاتيح التبديل ثنائية الاستقرار تناسب الحالات التي تحتاج فيها الالتزام بالنسب إلى البقاء مغلقًا. المشكلة الرئيسية هي التقلب العفوي للحالة الناجم عن الضوضاء النسخية.
مرشحات النطاق الترددي تناسب الحالات التي يكون فيها عامل النسخ مطلوبًا فقط ضمن نافذة تطورية محددة. المشكلة هي أن مستويات المحفز تحتاج إلى تحكم دقيق، وإلا فإن التوقيت ينزلق.
البوابات المنطقية تضيف التحديد من خلال طلب أكثر من مدخل واحد في نفس الوقت. على سبيل المثال، يمكن السماح بالتمايز فقط عندما يكون المحفز الخارجي موجودًا و تظهر الخلية الملف الشخصي الصحيح للـ miRNA الداخلي. هذا يساعد في تقليل خطر الالتزام الخاطئ.
الجدول أدناه يوضح البنى الرئيسية والمقايضات التي تأتي معها.
| الهندسة المعمارية | القابلية للعكس | الدقة الزمنية | تعقيد التكامل | حالة الاستخدام الرئيسية | المخاطر الرئيسية |
|---|---|---|---|---|---|
| مفتاح ثنائي الاستقرار | منخفض (بمجرد القفل) | متوسط | متوسط | الالتزام الدائم بالنسب | التقلب العفوي بسبب الضوضاء |
| مرشح النطاق الترددي | مرتفع (يعتمد على التركيز) | مرتفع | مرتفع | مراحل التطور العابرة | يتطلب تحكمًا محكمًا في المحفز |
| بوابة منطقية (AND/OR/NOT) | متغير | متوسط | متوسط-مرتفع | تنشيط خاص بنوع الخلية | التسرب في حالات الإيقاف |
| ربطة عنق / مدخلات متعددة | عالي | متوسط | متوسط | تكامل الإشارات المتعددة | يعتمد على استقرار miRNA الداخلي |
| miSFITs | عالي | متوسط | منخفض–متوسط | ضبط المخرجات المتدرجة | نطاق ديناميكي ضيق إذا كان الضبط ضعيفًا |
"من خلال تقليل عدد الطبقات الحسابية مع الحفاظ على الوظائف، تعالج هذه الاستراتيجية حواجز التوسع في هندسة الدوائر الجينية." - Nature Communications [9]
كل طبقة تنظيمية مضافة تزيد من الحمولة وتستنزف الموارد الخلوية. في الممارسة العملية، إذا كان تصميمان يقومان بنفس الوظيفة، فإن الأبسط عادة ما يكون الخيار الأفضل عندما يكون الحجم مهمًا.
بمجرد تحديد البنية، تكون المهمة التالية هي جعلها تصمد أمام التسرب المنخفض، وقمع الضوضاء والتحكم الآمن.
2.3 البناء من أجل الموثوقية، التسرب المنخفض والسلامة
يجب أن يبقى الدائرة مستقرة على مدى فترة طويلة من الثقافة. الأداء الجيد لفترة قصيرة لا يكفي للاستخدام الإنتاجي.
الأجزاء المتعامدة هي الدفاع الأول. المروجون، وعوامل النسخ والعناصر التنظيمية التي لا تتفاعل مع الآلات الأصلية تساعد في الحد من التأثيرات غير المستهدفة وتقليل فرصة أن تقوم الإشارات الذاتية بتشغيل الدائرة أو إيقافها.تم استخدام المحفزات المعدلة ذات الضيق العالي مثل PCREm لتقليل التسرب الأساسي في الأنظمة الثديية القابلة للتحفيز [6].
التنظيم الذاتي السلبي يستحق الإضافة أيضًا حيثما أمكن. إنه أحد الأنماط المعروفة لتقليل الضوضاء النسخية وإنتاج استجابة أكثر خطية لتركيز المحفز [6]. يمكن للحلقات الأمامية غير المتماسكة (iFFLs) إضافة طبقة أخرى من التحكم عن طريق تصفية التقلبات العشوائية، بحيث تستجيب الخلايا للإشارات المستمرة بدلاً من الارتفاعات الضوضائية القصيرة.
الإصدارات المعدلة للكودون (cm ) من عوامل النسخ الاصطناعية تجعل التوصيف أسهل أيضًا. إنها تتيح لك فصل التعبير المدفوع بالدائرة عن التعبير الجينومي الداخلي (g) أثناء التحقق [1]. قد يبدو ذلك كأنه تفصيل صغير، لكنه يوفر الوقت عندما تحاول معرفة ما إذا كان القراء يأتي من الدائرة أو الجينوم المضيف.
الوحدات الأمنية مطلوبة. حوالي 20% من الخلايا الجذعية البشرية متعددة القدرات تحمل طفرات مرتبطة بالسرطان [7]. لذلك، إذا كانت الدائرة ستدخل في خط مشتق من الخلايا الجذعية، فيجب أن تتضمن وحدة توقف النمو أو الإزالة القابلة للتحفيز. حمض الفانيليك هو محفز مفيد يجب إعطاؤه الأولوية هنا لأنه مضاف غذائي مرخص، مما يساعد في استخدامه كمحفز للدائرة في خطوط خلايا اللحوم المزروعة [1].
"توفر البيولوجيا التركيبية للمهندسين الوسائل لنشر الدوائر لضبط تعبير جينات متعددة بسهولة ودقة... للقضاء على الآثار الجانبية المحتملة غير المستهدفة." - npj Systems Biology and Applications [6]
تحدد هذه الخيارات الدوائر الخاصة بالنسب في القسم 3.
sbb-itb-ffee270
3. استراتيجيات الدوائر للتمايز العضلي، الشحمي والتحكم بالنسب
3.1 دوائر عضلية تفصل بين التوسع والتمايز النهائي
بمجرد تحديد بنية الدائرة، تكون المهمة التالية هي النشر الخاص بالنسب. بالنسبة لتكوين العضلات، فإن المشكلة الرئيسية بسيطة في التعبير ولكنها صعبة في التنفيذ: تحتاج الخلايا إلى التكاثر أولاً، ثم التحول إلى التمايز عند الطلب، دون الانجراف مبكرًا في أي اتجاه.
يُعتبر كاسيت Tet-On-MyoD أحد أكثر الطرق مباشرة للقيام بذلك. في هذا الإعداد، تتكاثر الخلايا تحت الظروف القياسية عند غياب الدوكسيسيكلين (DOX). أضف DOX، وستقود الدائرة الالتزام العضلي.استخدم الباحثون في مؤسسات صينية متعددة هذا النهج في الخلايا الليفية الجنينية للدجاج وأبلغوا عن تكوين فعال للأنابيب العضلية بعد التحفيز [4].
يقدم مفتاح تبديل ثنائي الاستقرار تحكمًا أكثر إحكامًا في الحالة. يمكن للأنظمة المبنية من مثبطات متبادلة مثل E-KRAB وPip-KRAB تحويل نبضة DOX قصيرة إلى برنامج عضلي مستقر [6]. في الممارسة العملية، يعني ذلك أن الحالة المتمايزة تبقى ثابتة بعد التحفيز بدلاً من التلاشي بمجرد إزالة الإشارة. كما أن إضافة التنظيم الذاتي السلبي يساعد أيضًا في تقليل الضوضاء العشوائية والحد من التمايز المتسرب خلال مرحلة التوسع [6].
هذا مهم لأن التمايز العضلي القياسي في المختبر غالبًا ما يتوقف في منتصف الطريق. عادة ما تكون مؤشرات الاندماج حوالي 50-60% فقط، مما يترك مجموعة احتياطية كبيرة غير مدمجة [3]. يمكن أن يؤدي تنشيط MyoD المدفوع بالدائرة إلى تحسين الالتزام، ولكن الالتزام وحده لا يضمن الاندماج الموحد. عندما يتم دمج دوائر MyoD مع تعديل MEK وNOTCH وRXR، يصبح الاندماج شبه الكامل في الثقافة ثنائية الأبعاد ممكنًا [3]. تتحكم الدائرة في توقيت دخول السلالة؛ تدفع الجزيئات الصغيرة خروجًا أنظف وأكثر انتظامًا من دورة الخلية.
3.2 التحكم في تكوين وبنية الأنسجة الدهنية والليفية
العضلات وحدها ليست كافية. بمجرد وضع التحكم العضلي، تكون المشكلة التالية هي التكوين: كم من الدهون يتشكل، وكم من ECM يتم ترسيبه، ومتى يتم تشغيل تلك البرامج. هنا، يكون التحكم الثنائي ON/OFF غالبًا ما يكون حادًا جدًا. ما تحتاجه الفرق عادة هو مخرجات متدرجة، خاصة حول نقطة التفرع الدهني-الليفي.
توفر miSFITs طريقة عملية لضبط التعبير على مراحل.من خلال وضع مواقع مستهدفة miRNA متحورة - على سبيل المثال، مواقع لـ miR-17 - في 3′UTR من الجينات الناتجة مثل PPARG أو BMP4، يمكن للباحثين اختيار مستويات التعبير من مكتبة متغيرة [5]. هذا يجعل تحفيز الخلايا الدهنية أشبه بمفتاح باهتة بدلاً من مفتاح ضوء. بدلاً من دفع الخلايا إلى استجابة كلية أو لا شيء، يمكن للفرق تعديل تكوين الخلايا الدهنية بعناية أكبر [5].
الأرومات الليفية ليست مجرد متفرجين هنا. فهي توفر بروتينات ECM التي تشكل النسيج [10]. هذا يجعل التحكم الليفي جزءًا من تصميم المنتج، وليس مجرد قضية جانبية. يمكن للدوائر المساعدة في إدارة التحول بين الحالات الليفية والدهنية، وفي الدواجن المزروعة، قد يكون تفعيل PPARG المباشر في الأرومات الليفية ضروريًا لتوليد ترسب دهني ذو معنى [10].
تناسب بنية ربطة العنق هذه المشكلة جيدًا لأنها تبقي الاستشعار والإخراج منفصلين. تقرأ طبقة الاستشعار الحالة الحالية للخلية، بينما تقوم طبقة الإخراج بتعديل PPARG، CEBPA أو منظمات النسب الأخرى. يساعد هذا الفصل في منع تشغيل برامج الأنسجة الدهنية أو الضامة قبل أن تصل الخلايا إلى المرحلة التطورية الصحيحة.
3.3 التحكم في نسبة النسب المتعددة والاستشعار المرتد
تتعامل دوائر التحكم في النسبة مع مشكلة مختلفة: ليس ما إذا كان التمايز يحدث، ولكن ما إذا كانت تركيبة السكان النهائية تبقى حيث ينبغي أن تكون. بالنسبة للمنتجات المقطوعة بالكامل، فإن الحصول على العضلات والدهون وECM بالنسب الصحيحة مهم بقدر أهمية جعل أي نسب تتمايز.
تبني هذه الأنظمة التحكم المرتد في الخلايا نفسها. تقيد المحفزات الخاصة بالحالة تعبير بروتين الإشارة للخلايا التي التزمت بالفعل بنسب معينة.تسمح الوحدات الباركرينية الاصطناعية بعد ذلك للخلايا العضلية الملتزمة بإطلاق إشارة مثبطة تقمع الالتزام الشحمي في الخلايا المجاورة. يشبه المنطق التثبيط الجانبي في أنظمة التطور دلتا-نوتش [1] [6]. حيث يصبح التشعب أكثر تعقيدًا، يمكن للبوابات متعددة المدخلات دمج الإشارات الخارجية مع إشارات الحالة الداخلية [9].
تعمل miSFITs أيضًا في هذه المرحلة. من خلال ضبط قوة إخراج BMP4 أو غيرها من المورفوجينات، يمكن للفرق تغيير توازن السلالة دون إعادة كتابة منطق القرار العلوي. في شبكات تنظيم الجينات المهندسة، زاد هذا النوع من التحكم كفاءة تمايز السلالة المستهدفة من 52% في الضوابط إلى 81% في iPSCs المهندسة [2]. في الخلايا الجذعية الميزنشيمية المعدلة، دفعت الدوائر الاصطناعية كفاءة تمايز القلب إلى 76% [2].
يقارن الجدول أدناه بين نهج السلالة الواحدة ونهج التحكم في النسبة باستخدام النقاط الأكثر أهمية في الإنتاج.
| الميزة | دائرة النسب الفردي | دائرة التحكم في النسبة |
|---|---|---|
| التعقيد | منخفض؛ عادةً ما يكون مروجًا ومحفزًا واحدًا قابلًا للتحفيز [4] | مرتفع؛ يتطلب بوابات منطقية وإشارات باراكرينية [6][9] |
| عبء المراقبة | منخفض؛ عادةً ما يتبع مراسلًا واحدًا [4] | مرتفع؛ يتطلب تتبع علامات النسب المتعددة [5] |
| المتانة | متوسط؛ عرضة للتنوع وتكوين الخلايا الاحتياطية [3] | مرتفع؛ يستخدم التغذية الراجعة والتثبيط الجانبي لتحقيق التوازن بين السكان [1] |
| قيمة الإنتاج | مرتفع للكتلة الحيوية؛ محدود لهياكل الأنسجة المعقدة [10] | ضروري للمنتجات المقطوعة بالكامل التي تحتاج إلى العضلات والدهون وECM [4] |
دوائر التحكم في النسبة تضيف حملاً أثقل للتحقق.لكن التغذية الراجعة المدمجة فيها يصعب مطابقتها مع التحكم في العمليات وحده، مما يضع المزيد من الضغط على اختيار النسخ واختبار العمليات.
4. من البناء إلى العملية: التحقق من الصحة، التوسع والملاءمة التنظيمية
4.1 استراتيجية التكامل واختيار النسخ لأداء مستقر
بعد تصميم الدائرة، يبدأ الجزء الصعب: إدخال تلك الدائرة في الخلايا بطريقة تبقى مستقرة خلال تمرير الإنتاج على نطاق واسع.
التوصيل الفيروسي العدسي غالبًا ما يكون فعالًا ويعطي تكاملات مستقرة بسرعة. لكن التكامل عشوائي. وهذا يعني المزيد من الاهتمام التنظيمي، بالإضافة إلى خطر انخفاض التعبير بمرور الوقت بسبب الإسكات. أنظمة الترانسبوزون مثل PiggyBac و Sleeping Beauty تقع في الوسط. يمكنها الحفاظ على الأداء عبر العديد من التمريرات، لكن لا يزال عليك فحص عدد النسخ ومواقع الإدراج.الجمال النائم , على سبيل المثال، تم استخدامه لتخليد خلايا الأقمار الصناعية البقرية بشكل مستقر عن طريق الإفراط في التعبير عن TERT وCDK4، مع خطوط تحتفظ بالإمكانات العضلية لأكثر من 40 تمريرة [10]. إدخال CRISPR يوفر التحكم الأكثر دقة في مكان هبوط التركيب والإعداد الجينومي الأكثر دقة، على الرغم من أن اختيار النسخ أبطأ والإنتاجية أقل.
| طريقة التكامل | التحكم في الإدراج | الاستقرار | قابلية التوسع | الاعتبارات التنظيمية |
|---|---|---|---|---|
| التوصيل الفيروسي Lentiviral | منخفض (إدماج عشوائي) | مرتفع، ولكنه عرضة للصمت | مرتفع | تدقيق تنظيمي أكبر بسبب الإدراج العشوائي وبقايا الفيروسات |
| Transposons (PiggyBac/SB) | متوسط | مرتفع على مدى العديد من الممرات | مرتفع | يتطلب فحص عدد النسخ ومواقع الإدراج |
| CRISPR Knock-in | مرتفع (محدد الموقع) | مرتفع جداً | متوسط | مفضل؛ يقلل من خطر تعطيل الجينات الذاتية |
| النواقل الإبيسومية | لا يوجد (خارج الكروموسوم) | منخفض؛ يمكن فقدانه أثناء الانقسام | منخفض | عائق تكامل أقل، ولكنه غير مناسب للتوسع طويل الأمد |
يحتاج فحص النسخ إلى القيام بأكثر من تأكيد وجود التركيب.يجب أن يتتبع الانحراف في التعبير، ملف الإدراج، حركية النمو، كفاءة التمايز، واحتفاظ النمط الظاهري عند عدد مرور مرتفع. يعتبر snRNA-seq مفيدًا هنا لأنه يمكن أن يستبعد النسخ المستنسخة الغنية بخلايا Pax7⁺/Ki-67⁻ الاحتياطية - الخلايا التي تترك دورة الخلية دون التمايز - قبل التوسع [3]. احتفظت EPSCs الخنزيرية مع دائرة Tet-On-PAX7 بتمايز عضلي عالي في حاملات دقيقة ثلاثية الأبعاد وثقافة التعليق على مدى 40 مرورًا [8].
4.2 كيف يتغير سلوك الدائرة في الثقافة ثلاثية الأبعاد، الحاملات الدقيقة والمفاعل الحيوي
بمجرد أن يكون لديك نسخة مستنسخة، فإن الاختبار التالي هو ما إذا كانت تتصرف بنفس الطريقة خارج 2D. في كثير من الحالات، لا تفعل ذلك. نادرًا ما ينتقل الأداء في 2D بشكل نظيف إلى التعليق، الحاملات الدقيقة، أو ثقافة السقالة لأن تدرجات الانتشار، حدود الأكسجين، والقص كلها تغير مخرجات الدائرة.
أحد الفحوصات الأولى هو انتشار المحفز. في المفاعلات الحيوية ذات الخزانات المحركة، تحتاج المحفزات الجزيئية الصغيرة إلى الوصول إلى الخلايا بشكل متساوٍ. في الواقع، يمكن أن تتشكل تدرجات، خاصة في الثقافات الكثيفة على الناقلات الدقيقة وداخل التجمعات أو نوى الهياكل. عادة ما تكون ثقافة التعليق أكثر ملاءمة للحوم المزروعة على نطاق واسع لأنها تدعم كثافة خلية أعلى وتوفر تحكمًا أكثر إحكامًا في العملية.
كما يصبح مراقبة حالة الخلية أصعب مع توسع النظام. يمكن أن تصبح إشارات المراسل الفلورية التي يسهل قراءتها بواسطة المجهر في 2D صعبة الحل في الهياكل ثلاثية الأبعاد المعتمة. يمكن أن توفر المؤقتات الفلورية - المجسات التي تحول الانبعاث من الأزرق إلى الأحمر مع نضوج البروتين - بيانات تنشيط الدائرة في الوقت الحقيقي في الموقع [1]. المسار المنطقي هو التحقق التدريجي: أولاً في 2D، ثم في 3D، ثم تحت ظروف المفاعل الحيوي النهائية [3] [8].
4.3 التوصيف، وثائق سلامة الغذاء واعتبارات المملكة المتحدة/الاتحاد الأوروبي
بعد اختبار العملية، يجب أن يظهر التوصيف أن وظيفة الدائرة، والظاهرة، والسلامة لا تزال قائمة. يجب أن تتضمن مجموعة البيانات الأساسية قياس التدفق الخلوي، qPCR مع تسلسلات cm، دورات زمنية لـ RNA-seq، وقراءات وظيفية مثل نسبة مساحة سلسلة الميوسين الثقيلة وتعبير الميوجلوبين [1]. لقد أظهرت وسائل التمايز الخالية من المصل المحسنة أنها ترفع تعبير الميوجلوبين إلى حوالي 30% من المستويات الموجودة في عضلات الأبقار الأصلية [3]. هذا يعطي الفرق معيارًا واضحًا بدلاً من هدف غامض.
تحتاج أيضًا إلى توثيق ملفات البروتين والأحماض الأمينية والدهون، إلى جانب الخصائص الحسية [10][3].
من وجهة نظر تنظيمية، ترسم المملكة المتحدة والاتحاد الأوروبي خطًا واضحًا بين خطوط الخلايا التي تتخلد تلقائيًا (غير المعدلة وراثيًا) والخطوط المعدلة وراثيًا. تحتاج الأخيرة إلى ملفات أمان أوسع [10][3]. يجب أن تظهر حزم الاستقرار الاحتفاظ بالنمط الظاهري والاستقرار الجينومي عبر الجسر الإنتاجي الكامل - من بنك الخلايا الرئيسي إلى خلايا الإنتاج النهائية - ويجب أن تأخذ سجلات التتبع في الاعتبار كل مرحلة بينهما [10]. إذا كان الدائرة تعتمد على محفز كيميائي، فإن إضافة آمنة غذائيًا أو مرخصة مثل حمض الفانيليك مفضلة على الدوكسيسيكلين [1].
يجب أن يكون المراقبة الجينية الروتينية أمرًا ضروريًا، ويجب توثيق مفتاح الانتحار أو الإزالة القابل للتحفيز كإجراء أساسي للتحكم في المخاطر [7]. يجب أيضًا تضمين وظيفته في ملف الأمان، خاصة مع استمرار تشكيل القواعد في المملكة المتحدة والاتحاد الأوروبي للحوم المزروعة.
5. خارطة طريق عملية وخاتمة
5.1 خارطة طريق تنفيذية مرحلية لفرق اللحوم المزروعة
أنظف طريق من المفهوم إلى الإنتاج هو سير العمل المرحلي.
المرحلة 1 هي التصميم. ابدأ بتحديد السلالة المستهدفة، ثم استخدم snRNA-seq لتأكيد العوائق الرئيسية قبل اختيار بنية الدائرة. هذه الخطوة مهمة لأن الدائرة يمكنها فقط حل القيود التي حددتها بالفعل.
المرحلة 2 هي البناء والتحقق ثنائي الأبعاد. قم ببناء التركيب وتحقق من أن الدائرة تعمل كما هو مقصود في ثنائي الأبعاد، باستخدام قراءة بسيطة للمراسل.في هذه المرحلة، الهدف بسيط: تأكيد أن المنطق يعمل قبل الانتقال إلى نماذج أصعب وأكثر تكلفة.
المرحلة 3 هي اختبار الإجهاد ذو الصلة بالحجم. الانتقال إلى الأنظمة ثلاثية الأبعاد والظروف ذات الصلة بالمفاعل الحيوي، ثم مقارنة المخرجات مع الأساس ثنائي الأبعاد. هنا تبدأ العديد من التصاميم في إظهار نقاط ضعفها، خاصة عندما تدخل تأثيرات النقل الجماعي، القص، والمصفوفة في اللعب.
المرحلة 4 هي دمج التنظيم والسلامة، ويجب أن تعمل بالتوازي مع المرحلة 3. لا ينبغي أن تنتظر أعمال السلامة والتنظيم حتى النهاية. قم بتشغيلها جنبًا إلى جنب مع التوسع، بما في ذلك توثيق أي وحدة أمان قابلة للتحفيز.
5.2 الحصول على الأدوات والمواد الممكنة من خلال Cellbase
بمجرد تحديد سير العمل، غالبًا ما يصبح الحصول على المواد هو الخطوة المحددة للمعدل.
- خطوط الخلايا
- الوسائط الخالية من المصل والمحددة كيميائيًا
- الهياكل الداعمة
- مكونات المفاعلات الحيوية
- المستشعرات
- معدات التحليل
الوصول الموثوق إلى المواد المتوافقة في كل مرحلة له تأثير مباشر على سرعة توصيف سلوك الدائرة تحت ظروف ذات صلة بالحجم.
5.3 النقاط الرئيسية
تمنح الدوائر الجينية الاصطناعية فرق اللحوم المزروعة تحكمًا برمجيًا في التوقيت والعتبات وتوازن السلالة الذي لا يمكن للبروتوكولات المعتمدة على الوسائط فقط أن تضاهيه. يؤثر اختيار الهندسة المعمارية على القابلية للعكس والتسرب والسلامة.تُفضل الأنظمة القابلة للتحفيز عادةً لأنها توفر تحكمًا شرطيًا وحملًا أيضيًا أقل [6].
"يمكن استخدام مجموعة أدوات البيولوجيا التركيبية لإنشاء خطوط خلوية ذات تعبير جيني قابل للتعديل، والتي، عند اقترانها مع PAT والنمذجة الحاسوبية، يمكن أن تمكن أنظمة التحكم ذات الحلقة المغلقة من تقديم أفضل إنتاجية وجودة للمنتج." - npj Systems Biology and Applications [6]
لا يعتمد النشر الناجح على مشكلة بيولوجية فقط. بل يعتمد على الربط الوثيق بين هندسة الدوائر، تصميم العمليات الحيوية، الوثائق التنظيمية، والتوريد.
الأسئلة الشائعة
كيف تحسن الدوائر الجينية التركيبية من اتساق التمايز؟
يمكن للدوائر الجينية التركيبية أن تجعل التمايز أكثر اتساقًا لأنها تمنحك تحكمًا مبرمجًا في سلوك الخلية والالتزام بالنسب.في الممارسة العملية، يعني ذلك استخدام عمليات منطقية معيارية لضبط تعبير الجينات وعوامل النسخ بتوقيت دقيق.
هذا التوقيت مهم. يساعد الخلايا على التحرك عبر تغييرات حالة محددة بالترتيب الصحيح، بدلاً من الانجراف إلى مصائر مختلطة أو غير مرغوب فيها. كما يقلل من التمايز غير المستهدف ويقلل الضوضاء عبر الثقافة.
العائد واضح: مجموعات خلايا أكثر تجانسًا واستقرارًا ونضجًا لإنتاج اللحوم المزروعة.
أي تصميم دائرة يناسب التحكم في التولد العضلي أو التولد الشحمي؟
في أبحاث اللحوم المزروعة، يمكن دفع نفس الخلايا الليفية للدجاج إلى أي من السلالتين. التولد العضلي يتبع مجموعة من بروتوكولات التحريض، بينما يمكن تشغيل التولد الشحمي عن طريق تعريض الخلايا لمداخل مثل مصل الدجاج أو الأحماض الدهنية.
من هناك، يمكن التحكم في مصائر هذه الخلايا خطوة بخطوة داخل هياكل هيدروجيل ثلاثية الأبعاد لبناء هياكل لحم بنسب محددة من الدهون والكولاجين.
لماذا تتصرف الدوائر الجينية بشكل مختلف في الثقافة ثلاثية الأبعاد؟
في الثقافة ثلاثية الأبعاد , تتصرف الدوائر الجينية بشكل مختلف لأن الخلايا تتعامل مع مدخلات فيزيائية وهيكلية لا توجد ببساطة في الطبقات الأحادية ثنائية الأبعاد. تشمل هذه المدخلات التوتر الميكانيكي، إجهاد القص، صلابة المصفوفة، وكثافة الخلايا المحلية.
يمكن أن تغير هذه الإشارات مسارات الإشارة مثل Notch. كما يمكنها تغيير كيفية اكتشاف الدوائر الاصطناعية للقوة وتنسيق الاستجابات اللاحقة، بما في ذلك التصاق الخلايا وتكوين الأنسجة.









