

对于培养肉生产的研究人员来说,减少细胞凋亡对于提高生物反应器中细胞的存活率和生产力至关重要。营养耗尽、渗透失衡和废物积累等压力因素常常会引发细胞死亡,降低产量。抗凋亡基因可以通过延长细胞在培养过程中的寿命来缓解这些挑战。以下是顶级基因及其作用的快速概述:
- BCL-2: 防止线粒体孔形成,在凋亡开始时阻止其发生。对未分化细胞有效,但需要与促凋亡蛋白仔细平衡。
- BCL-xL: 在分化过程中保护细胞并支持能量代谢。适用于生物反应器中的高压阶段。
- MCL-1: 对营养变化提供快速响应,并在分化过程中保持稳定。与其他基因结合效果良好。
- BIRC5 (Survivin) : 抑制caspases以阻止下游的细胞凋亡。在快速分裂的细胞中支持增殖。
- XIAP: 一种有效的caspase抑制剂,在极端压力条件下有效,例如高密度培养。监测这些条件需要选择用于培养肉类生物反应器的传感器以实时跟踪营养水平和废物积累。
快速比较
| 基因 | 关键作用 | 分化期间的稳定性 | 最佳使用案例 |
|---|---|---|---|
| BCL-2 | 阻止早期凋亡(BAX/BAK) | 稳定 | 保持未分化细胞 |
| BCL-xL | 防止caspase激活,支持代谢 | 阶段特异性 | 压力下的分化细胞 |
| MCL-1 | 对营养变化的快速响应 | 稳定 | 多阶段生存 |
| BIRC5 | 抑制下游caspase | 随分化减少 | 快速分裂细胞 |
| XIAP | 广泛的caspase抑制 | 稳定 | 高压生物反应器条件 |
1.BCL-2
BCL-2 是一个研究透彻的抗凋亡基因,在内在(线粒体)凋亡途径中起着关键作用。该途径是细胞死亡的主要机制,通常在生物反应器压力下,如营养短缺或低氧水平时,在培养肉细胞中被触发。
BCL-2 通过结合并中和像 BAX 和 BAK 这样的促凋亡蛋白来发挥作用。这一作用防止了线粒体孔的形成,阻止了细胞色素 c 的释放,并中止了凋亡的下游级联反应。此机制对于延长培养肉生产中细胞的可存活寿命至关重要。正如 Rønning SB 等人所解释的:
"Bcl-2 和 Bax 之间的比例决定了细胞经历凋亡的易感性。"[5]
除了其在线粒体中的作用外,BCL-2 还存在于内质网 (ER) 中。在这里,它降低了钙水平并抑制了IP3受体介导的钙释放,从而减轻了钙诱导的细胞凋亡——这是高密度生物反应器培养中的常见问题。管理这些扩展挑战是行业的主要关注点。这种双重定位使BCL-2能够保护细胞免受多种凋亡触发因素的影响。 BCL-2的分子结构由一个八α螺旋束和四个明确的BH结构域组成,使其成为基因改造的理想候选者。诸如CRISPR/Cas9介导的过表达或稳定载体整合等技术可以利用BCL-2在培养肉细胞系中的保护能力。此外,由于BCL-2在牛和猪等哺乳动物物种中高度保守,从一个细胞系的研究结果通常适用于其他用于培养肉生产的细胞系[3].
。然而,有一个关键的警告:必须仔细管理BCL-2和促凋亡蛋白如BAX之间的平衡。如果促凋亡信号过强,即使BCL-2表达水平很高,也可能无法防止凋亡[2]. 。监测这种平衡对于实现最佳细胞活力至关重要。
sbb-itb-ffee270
2. BCL-xL
BCL-xL, 由BCL2L1基因编码,在BCL-2家族中起着核心作用,通过定位于外线粒体膜并防止凋亡。它通过对抗促凋亡蛋白如BAX和BAK来实现这一点。此外,它抑制了切割的caspase-3(CASP3),这对于阻止细胞死亡至关重要。此机制在高密度生物反应器培养, 中尤为重要,因为代谢压力可能威胁细胞活力。
有趣的是,BCL-xL的活性与特定的分化阶段一致。在某些阶段,其表达增加,而其他抗凋亡蛋白如BCL-2和MCL-1则保持不变。这突显了其在分化过程中维持细胞存活的重要性。如在细胞死亡&疾病:
中所述:"BCL-xL/BCL2L1是一种关键的抗凋亡蛋白,促进分化...细胞的存活。" [2]
除了在凋亡中的作用外,BCL-xL还支持细胞能量代谢。它增强了糖酵解和氧化磷酸化,确保高代谢活性。抑制BCL-xL已被证明可以减少代谢基因的表达,并降低基础和最大线粒体呼吸。这一功能对于依赖持续代谢输出的培养肉细胞尤为重要。
BCL-xL与培养肉研究中常用的基因编辑策略高度兼容。像慢病毒转导这样的技术允许BCL2L1基因的稳定整合,而多西环素诱导的CRISPR/Cas9系统则提供了对其表达的精确时间控制 [2][6]. 这种精确度通常通过先进的生物过程控制软件. 进行管理。这些属性使BCL-xL成为提高培养肉生产中细胞系活力的有力候选者。
对于代谢需求高的分化阶段,BCL-xL可能比BCL-2更有效。研究人员可以使用抑制剂WEHI-539来测试细胞系对BCL-xL的依赖性,然后再进行永久性基因改造。此外,与MCL-1共同表达BCL-xL可以进一步提高细胞存活率,因为这些蛋白质在某些耐药细胞类型中被观察到协同工作。 3. MCL-1 MCL-1(髓系细胞白血病-1)在调节内在凋亡途径中起关键作用。它位于线粒体外膜,通过结合和隔离促凋亡蛋白BAX和BAK,阻止它们的寡聚化和随后的膜通透化,从而防止凋亡。这一作用阻止了细胞色素c的释放,在凋亡级联反应到达执行阶段之前将其阻止。此外,MCL-1以高亲和力结合BH3-only蛋白,如Bim、PUMA和NOXA[8]. 与BCL-2和BCL-xL类似,MCL-1在抵抗凋亡信号方面至关重要,尤其是在生物反应器压力期间。
MCL-1的一个独特属性是其短半衰期,使其表达对营养可用性和代谢信号高度响应,特别是通过AMPK/mTOR通路。研究表明,卡路里摄入减少25%可以使MCL-1翻译减少约39% ± 10% [7]. 这种敏感性对于培养肉生产尤其重要,因为在大规模悬浮培养过程中(需要仔细生产规模规划)生长介质成分波动或营养耗尽时,MCL-1水平可能显著降低。这种减少会损害细胞活力,削弱通过抗凋亡策略实现的IVCC(整体活细胞浓度)改善。为此,支持强大mTORC1活性的无血清培养基配方是必不可少的[7].
MCL-1的另一个显著特点是在分化过程中的稳定性。在胰腺祖细胞模型中,MCL-1的表达在整个17天的分化过程中保持稳定,而BCL-xL则表现出阶段依赖性变化[2]. 这种稳定性使MCL-1在培养肉应用中特别有利,因为细胞需要在多个成熟阶段存活,而无需精确计时的干预。&
基因编辑工具可以用来修改MCL-1,就像其他抗凋亡基因一样,使其成为细胞系工程的多功能目标。
当与其他抗凋亡基因结合使用时,MCL-1提供了额外的好处。例如,将MCL-1与BCL-xL配对显示出协同效应——同时抑制这两种蛋白质将生存药物的EC50从约10 μM降低到不到20 nM[6]. 这种方法可以显著改善培养肉生产高压阶段的细胞存活率。
4. BIRC5 (Survivin)
BIRC5, 通常被称为Survivin,是凋亡抑制蛋白(IAP)家族的成员[2]. 与在线粒体膜上作用以防止凋亡启动的BCL-2家族蛋白不同,BIRC5在更下游起作用。它阻止负责执行凋亡的caspases,有效地作为对抗程序性细胞死亡的最后一道防线[10].
在悬浮培养中,营养耗尽、代谢废物积累和机械剪切应力等压力因素可以触发细胞凋亡。通过在后期阶段抑制caspase活性,BIRC5的过表达有助于延长细胞的存活率和生产力。这导致了活细胞浓度时间积分 的改善 - 这是优化细胞培养性能的关键指标[9]. KAIST的研究员Eric Baek, 解释道:
"通过克服细胞死亡,特别是细胞凋亡,提高活细胞浓度时间积分,是高效生产治疗性蛋白质[和细胞]的最广泛使用的策略之一。" [9]
这种下游干预已显示出提高培养肉细胞系生物反应器产量的效果,包括猪卫星细胞和牛成肌细胞.
最有效的策略涉及组合工程, 将BIRC5与BCL-2或BCL-xL等线粒体保护剂配对。来自约翰霍普金斯大学的Michael Betenbaugh教授强调了这种方法:
“在级联的多个点阻止细胞死亡的策略可能限制这些凋亡信号的放大。”[10]
通过结合BIRC5的半胱天冬酶抑制与上游线粒体保护,研究人员可以建立针对凋亡的多层防御。
BIRC5还可以无缝集成到基因编辑工作流程中。CRISPR/Cas9 是创建具有过表达的稳定细胞系的领先方法[9], 尽管锌指核酸酶提供了一种精确的替代方案。在承诺进行基因组整合之前,siRNA 可用于通路验证[9].
5. XIAP
XIAP(X 链接凋亡抑制蛋白)被认为是 IAP(凋亡抑制蛋白)家族中最有效的半胱天冬酶抑制剂。与 BCL-2 和 MCL-1 等基因一起,XIAP 在靶向凋亡的执行阶段中起着关键作用。如在 Genes & Development:
“XIAP 被认为是体外最有效的半胱天冬酶抑制剂。”[12]
XIAP 采用两种不同的机制来抑制凋亡。首先,其 BIR2 结构域和连接区阻断效应半胱天冬酶-3 和 -7。其次,其BIR3结构域抑制caspase-9,有效阻止了内在线粒体凋亡途径。此外,其C端RING结构域促进了目标caspases的泛素化和随后的蛋白酶体降解。通过干预内在和外在凋亡途径,XIAP在应对营养短缺、代谢副产物和机械应力等凋亡诱因方面表现出高度有效——这些因素在培养肉生产系统中常见。其功能性因其在物种间的强保守性而得到进一步增强。 例如,人类XIAP与牛(Bos taurus)共享87.7%的蛋白质同一性,与小鼠(Mus musculus)共享89.5%的蛋白质同一性。这种高度相似性使得来自哺乳动物模型系统的研究能够可靠地应用于用于培养肉生产的细胞系。
XIAP可以使用shRNA、反义寡核苷酸或CRISPR/Cas9等工具进行调控[11]. 在极端压力下,其RING结构域可能会诱导自泛素化[12], 而SMAC/DIABLO和HTRA2等内源性抑制剂可以将XIAP从caspases中置换出来[11][13]. 这些发现使得XIAP成为基因编辑方法的一个有吸引力的目标,旨在优化用于培养肉开发的细胞系。
快速比较抗凋亡基因
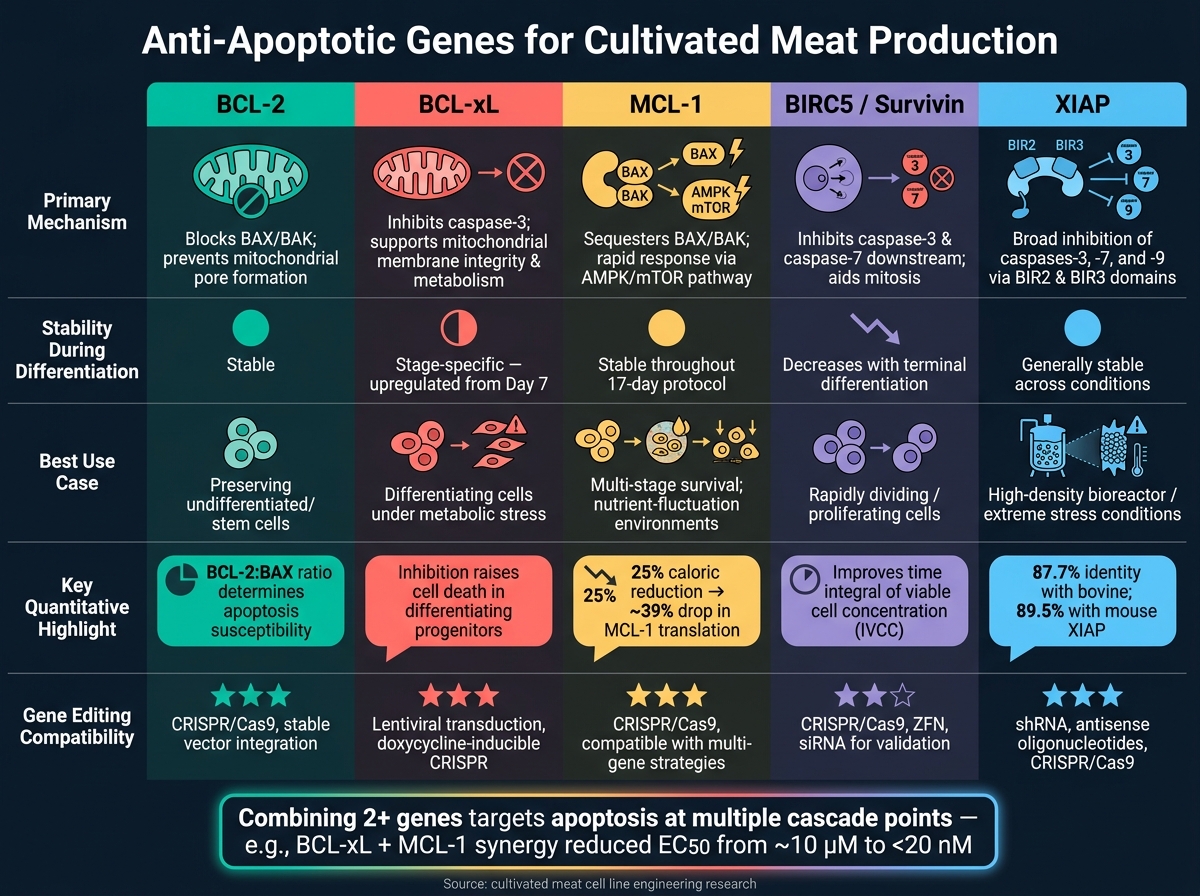
培养肉用抗凋亡基因:并排比较
在培养肉生产中,了解不同抗凋亡基因的功能有助于优化细胞系工程。每个基因都有其独特的机制、分化过程中的行为和潜在应用。下表总结了这些差异,使您更容易决定哪个基因或基因组合最适合您的需求。
| 基因 | 主要机制 | 表达稳定性 | 报告的生存影响 | 编辑兼容性 |
|---|---|---|---|---|
| BCL-2 | 阻止促凋亡的BAX/BAK,确保未分化细胞的存活 [2] | 在分化过程中保持相对稳定[2] | 对于维持初始干细胞库至关重要[2] | 与编辑工具高度兼容 |
| BCL-xL | 抑制切割的caspase-3;维持线粒体膜的完整性和代谢[2] | 从分化第7天开始上调[2] | 支持分化祖细胞至关重要;其抑制增加细胞死亡 [2] | 与编辑工具高度兼容 |
| MCL-1 | 作为BCL-2家族的一部分调节促凋亡信号[2] | 在谱系特化过程中表达保持稳定[2] | 提供广泛的生存益处,但缺乏像BCL-xL那样的阶段特异性效应[2] | 与编辑工具高度兼容 |
| BIRC5 (Survivin) | 阻断caspase-3和caspase-7;在有丝分裂期间帮助染色体分离 | 在增殖细胞中含量高;随着终末分化而减少 | 支持快速分裂细胞的生存和增殖 | 兼容shRNA敲低和CRISPR编辑 |
| XIAP | 抑制多种caspase,提供广泛的凋亡保护 | 在各种条件下通常稳定 | 在压力下特别有效,例如高密度生物反应器条件 | 与编辑工具高度兼容 |
BCL-xL 因其在促进细胞存活和支持代谢活动方面的双重作用而脱颖而出,特别是在促凋亡蛋白如BAK自然下降的关键分化阶段。BCL-2, 另一方面,非常适合保存未分化细胞,而XIAP提供广泛的保护,尤其是在高密度培养等压力环境中。
没有单一基因在每种情况下都能发挥最佳作用。例如,BIRC5在需要快速细胞分裂的情况下特别有用。在实践中,结合两个或多个基因通常能提供最有效的保护,同时应对多种凋亡触发因素。
这些发现为将这些基因纳入细胞系工程策略以用于培养肉生产提供了基础。这包括选择合适的培养肉输入以确保可扩展性。
在培养肉细胞系工程中使用这些基因
为了提高培养肉生产中的细胞活力,战略性地整合关键基因至关重要。识别抗凋亡基因是不够的——将其有效整合到细胞系中才是关键。通常采用两种主要策略:过表达抗凋亡基因如BCL-2, BCL-xL, 和MCL-1以增强细胞存活,或敲除促凋亡基因如BAX, BAK, 和BOK以消除细胞死亡的驱动因素。结合这些方法通常会产生更适合大规模生产的细胞系[1].
现代基因编辑工具如CRISPR/Cas9允许同时编辑,例如一步敲除Bak1, Bax, 和Bok。替代方法如ZFNs或RNA干扰可用于暂时降低caspases的活性(e.g. caspases-3, -7, -8, 和 -9)。对于过表达策略,合成启动子确保基因如 BCL-2在放大过程中保持一致和高水平的表达,这对于在补料分批或连续培养系统. 中维持细胞性能至关重要。这些组合方法加强了用于培养肉应用的细胞系开发。
这种基因改造直接影响改进的整体活细胞浓度 (IVCC), ,这是培养肉生产中的关键指标。细胞死亡在分化的前五天最为明显,因此早期干预如 BCL-2或BCL-xL等基因是必不可少的。发表在 Cell Death & Disease的研究表明,BCL-xL表达随着细胞分化而增加,表明更成熟的祖细胞在很大程度上依赖其保护作用 [2]. 通过监测BCL-2家族基因在生长阶段的表达水平,可以精确地安排干预措施以达到最大效果。
“通过建立稳定的细胞系,过表达抗凋亡基因或下调促凋亡基因,最终产品的产量可以提高,因为细胞对环境压力的抵抗力增强。” - Gyun Min Lee 等人 [1]
对于基于生物反应器的生产,细胞还必须被设计以承受高渗透压应激 和营养剥夺. 在扩大规模之前,必须使用诸如Western blot或FACS等工具验证基因编辑。对于寻求专门细胞系或适合高密度生物反应器环境的遗传材料的研究人员,像
结论
为培养肉细胞系选择抗凋亡基因需要量身定制的方法。像BCL-2, BCL-xL, 和 MCL-1这样的基因在保护细胞方面各自发挥着独特的作用,但它们的成功取决于细胞类型、发育阶段以及生产过程中遇到的特定压力等因素。正如研究中所强调的:
"抗凋亡和促凋亡成员之间的平衡最终决定了细胞的生死"[2]
除了生存之外,抗凋亡工程还保留了代谢功能。例如,像BCL-xL这样的蛋白质与维持糖酵解和氧化磷酸化密切相关。然而,执行不当的干预措施可能会破坏这些关键过程[2] . 确保工程化细胞系在整个生产过程中保持其预期的身份和代谢活性是一个关键但有时被忽视的步骤。这些见解正在塑造细胞系工程的未来。
新的多基因方法正在出现,它们将保护基因的过表达与CRISPR敲除促凋亡基因如BAX , BAK1, 和 BOK结合起来,以创建更强大的工业用细胞系[1]. 代谢分析工具,如生物能量测定,正变得至关重要,以确认这些基因改造是否增强了整体细胞性能。对于研究人员采购 猪细胞系, 遗传材料或生物反应器设备,
常见问题
我应该从哪个抗凋亡基因开始用于我的细胞系?
BCL-2通常被建议作为处理细胞系的起点。这个研究充分的抗凋亡基因因其提高细胞存活能力而被认可,使其在培养肉研究中成为一个受欢迎的选择。其支持细胞活力的功能使其成为早期实验的实用选择。
是过表达抗凋亡基因更好,还是敲除促凋亡基因更好?
在培养肉生产中,增加抗凋亡基因的表达,如BCL-2家族成员BCL-xL , 往往比禁用促凋亡基因产生更好的结果。这一策略支持细胞存活和增殖——生产规模化的关键因素——同时保留细胞的自然调节系统。
通过增强抗凋亡基因活性,细胞在压力条件下对凋亡的抵抗力更强。这使其在培养过程中成为一种更可靠和安全的方法来维持细胞活力。
如何确认抗凋亡编辑提高了我的生物反应器中的IVCC?
要确定抗凋亡基因编辑是否增强了体外细胞活力和增殖(IVCC),您需要一个系统的方法:
- 评估活力和增殖率: 使用细胞计数或流式细胞术等方法来测量基因编辑前后的这些速率。
- 验证基因表达: 使用qPCR或Western blotting等技术可以确认目标基因的成功表达。
- 监测凋亡标志物: 检查如caspase活性等标志物,以确保编辑有效减少凋亡。
为了进行全面评估,关键是要在生物反应器中测试编辑细胞的长期稳定性和增殖能力。这确保了改进在多个培养周期中持续存在。










