

高通量CRISPR筛选正在通过实现精确的基因改造来提高细胞系性能,从而改变培养肉行业。以下是您需要了解的内容:
- 关键挑战: 培养肉生产需要能够高效生长、抵抗衰老并分化为肌肉和脂肪组织的细胞系。
- CRISPR的作用: 通过同时靶向数千个基因,这些平台识别出能够增强生长、延缓衰老并支持分化的基因编辑。
- 显著发现: 研究表明,在TP53和 PTEN基因在 牛间充质干细胞中敲除可以在30天内将增殖提高至1000倍,并将其寿命从100天延长至200天。
- 应用: CRISPR工具如敲除筛选、CRISPRi和CRISPRa被用于优化细胞生长、调节基因表达,并平衡增殖与分化。
- 行业工具: 先进技术如RMCE、RNA-seq和单细胞平台将CRISPR结果与多组学数据整合,确保精确且可扩展的改进。
对于生物工艺工程师和研发专业人员,这些创新解决了在扩大培养肉类工艺时的关键瓶颈,同时保持细胞质量和功能。CRISPR与自动化系统和定制资源的整合,如
CRISPR-Cas9 基因组范围敲除筛选的基础知识
CRISPR-Cas9 在大规模基因编辑中的工作原理
CRISPR-Cas9 系统依赖于与单导向 RNA (sgRNA) 配对的 Cas9 核酸酶来靶向特定的 DNA 序列。一旦 sgRNA 将 Cas9 引导至所需的基因组位置,酶就会在 DNA 中产生双链断裂。此断裂主要通过非同源末端连接 (NHEJ) 修复,这是一种容易出错的过程,通常会引入小的插入或缺失 (indels)。这些 indels 可以导致移码突变,有效地破坏目标基因的功能[1]. 这种精确的机制是进行基因组范围敲除筛选的基础,这对于识别细胞行为的关键调节因子至关重要。
对于大规模筛选,研究人员使用多样化的sgRNA库,通常通过慢病毒转导传递到混合细胞群中。为了确保每个细胞仅接受一个基因改变,保持低感染复数(MOI约为0.3)[1]. 随着时间的推移,具有优势突变的细胞往往比其他细胞更成功地增殖,这一现象在各种细胞类型和实验条件下都被观察到。
替代传递方法,如重组酶介导的盒交换(RMCE),通过靶向特定基因组“着陆垫”来提供额外的精确性,以减少整合位点的变异性。例如,一项使用CHO-K1细胞的研究采用无病毒的RMCE方法筛选了21,585个基因中的111,651个独特gRNA。该方法在16天和37天的时间段内识别出对细胞适应性至关重要的基因[7].
基因组范围筛选的好处
基因组范围的敲除筛选利用CRISPR-Cas9的准确性系统地研究数千个基因。这使研究人员能够发现影响细胞存活、生长和应对压力的基因。除了遗传因素,优化表面功能化对于改善这些系统中的细胞附着和生长至关重要。这种无偏探索对于培养肉生产, 尤其重要,因为间充质干细胞(约占细胞来源的25%)通常面临有限增殖和早期衰老等挑战[1].
sbb-itb-ffee270
混合CRISPR文库筛选方法
构建混合CRISPR文库
混合CRISPR文库始于精心挑选的单导向RNA(sgRNA)集合。在培养肉研究的背景下,目标文库通常设计为专注于特定的基因家族,例如转录因子或细胞增殖的调节因子。这种方法有助于在保持对所需表型相关特征的关注的同时,平衡成本与可扩展性。[1].
该过程始于合成寡核苷酸作为一个池,通过PCR扩增它们,并将其克隆到一个递送载体中。例如,2025年初构建的一个牛特异性文库包括3,000个sgRNA,靶向603个基因,以识别影响干细胞扩增的因素。[1]. 在更大规模上,全基因组筛选可以达到更高的复杂性。一个例子是中国仓鼠卵巢(CHO)细胞筛选,使用了111,651个独特的gRNA来靶向21,585个基因。[7].
慢病毒转导通常用于以低感染复数(约0.3)传递这些文库,确保每个细胞仅经历一次基因修饰[1]. 或者,无病毒方法如重组酶介导的盒交换(RMCE)将gRNA文库整合到主细胞系内的预定基因组“着陆垫”中。该技术实现了99.9%的gRNA覆盖率,偏差极小[7].
为了保持统计可靠性,研究人员确保高覆盖率——通常每个sgRNA有500到600个细胞[1] [7]. 一些平台使用可诱导的Cas9(iCas9)系统,使基因编辑的发生时间得到精确控制。例如,可以在细胞达到特定状态后触发编辑,如高密度或衰老开始。这种时间控制对于研究非增殖阶段特别有用,这对于通过选择原代细胞系与永生化细胞系来克服衰老障碍以扩大培养肉生产规模至关重要 [4].
一旦构建了文库,研究人员就会进行靶向筛选测定以评估基因功能。
培养肉细胞系的筛选方法
在构建文库后,研究人员使用竞争测定和功能分选技术评估细胞性能。一种广泛使用的方法是基于竞争的增殖测定,它识别赋予生长或衰老抵抗的遗传变化——这是优化培养肉细胞系的关键特征。
短期筛选(约30天)识别立即影响细胞周期的基因,而长期筛选(最长200天)则专注于帮助细胞克服复制性衰老的基因。这是扩大培养肉生产规模的关键挑战 [1]. 对于更复杂的特征,如增强蛋白质分泌或特定标记的表达,采用荧光激活细胞分选(FACS)。一个例子是“冷捕获分泌测定”,通过在分选前捕获细胞表面的分泌蛋白来分离生产性细胞群体 [7] [5].
验证是确认筛选结果的关键步骤。例如,细胞适应性(CelFi)测定跟踪随时间变化的移码突变与非移码突变的比例。如果具有移码突变的细胞从群体中消失,这表明目标基因对于细胞适应性至关重要 [2].
2025年6月,由丁世杰领导的研究人员在南京农业大学使用CRISPR/Cas9创建了CDKN2A–/– 猪卫星细胞系. 这些经过改造的细胞在无血清条件下至少维持了15代的稳定增殖,同时保留了干性标记。当播种到植物基3D可食用支架上时,它们形成了具有改进质地的肉状结构,包括增强的咀嚼性和粘性 [8] .
“这些发现展示了CRISPR筛选在优化牛干细胞特性方面的实用性,并为未来更大规模的培养肉生产提供了一条途径。" – 通讯生物学 [1]
哺乳动物细胞中的汇集CRISPR基因筛选 | 协议预览
用于可逆基因调控筛选的CRISPRi和CRISPRa
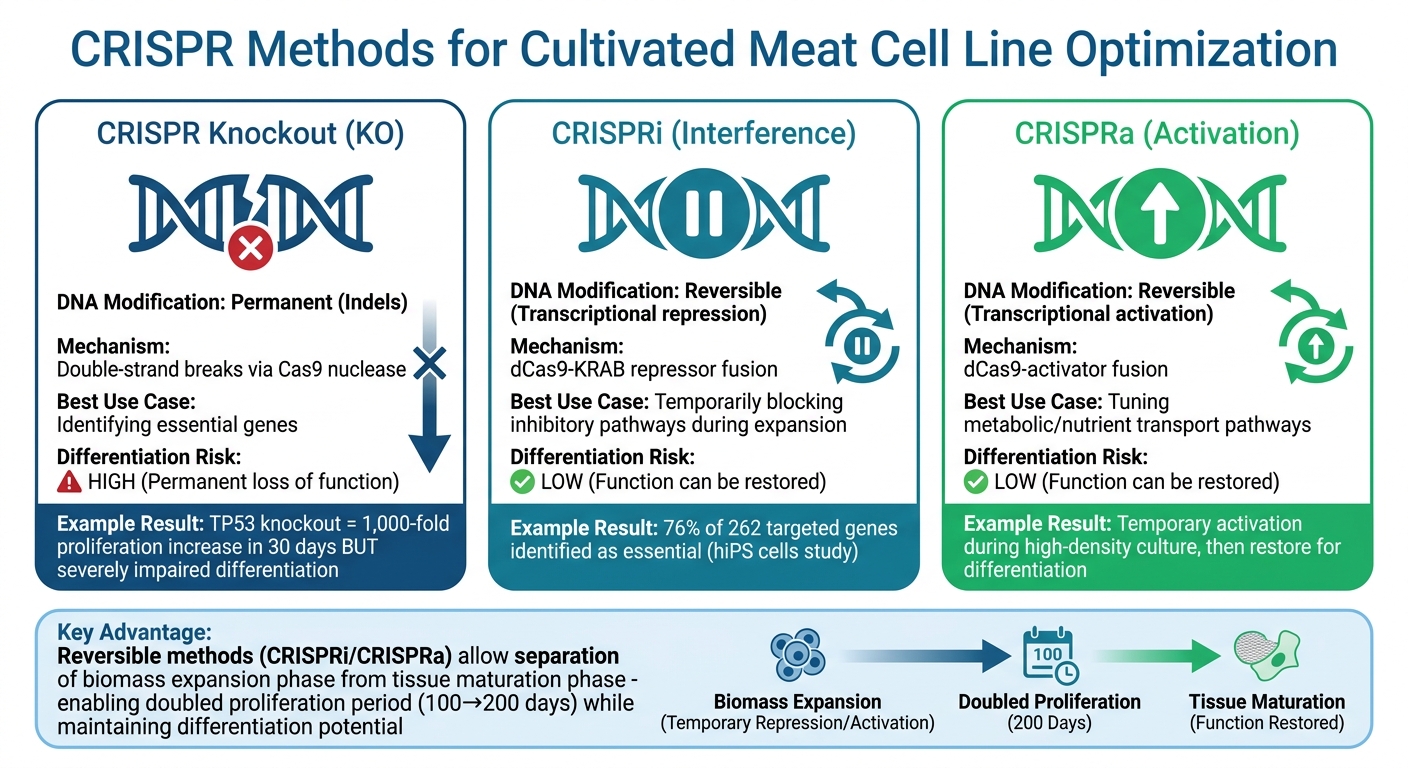
培养肉的CRISPR基因编辑方法:敲除与CRISPRi/CRISPRa比较
在功能基因组学中使用CRISPRi和CRISPRa
在提高培养肉生产方面,CRISPR干扰(CRISPRi)和CRISPR激活(CRISPRa)提供了强大的工具。这些技术使用与抑制剂或激活剂配对的非活性Cas9蛋白,使研究人员能够在不对DNA进行永久性改变的情况下暂时调整基因表达 [10].
这种可逆性对于解决一个重大挑战尤为重要:促进细胞快速生长的基因往往会干扰其向肌肉或脂肪组织分化的后期阶段。例如,在牛间充质干细胞中永久敲除TP53基因可以在短短30天内将增殖能力提高1000倍以上,但严重损害其分化能力[1]. CRISPRi通过在生物反应器中培养肉类的生物质扩增期间暂时阻断抑制分化的途径,提供了一种更灵活的解决方案一旦细胞准备好进行组织成熟,正常的基因功能可以恢复。
2025年10月,来自 荷兰癌症研究所的研究人员如Gabriele Casagrande Raffi和Roderick L. Beijersbergen开发了一种可诱导的CRISPR系统。这种方法将基因编辑推迟到细胞达到特定状态时进行——例如高密度或非增殖阶段——有助于保持细胞活力 [4].
与传统的RNA干扰(RNAi)相比,CRISPRi因其精确性而脱颖而出。RNAi通常导致结果不一致和非靶向效应,而CRISPRi则提供更可靠和特定的基因抑制 [2] . 另一个优点是CRISPRi避免了触发与p53相关的毒性,这通常是由DNA损伤反应引起的。在2025年,由王立勤领导的中山大学癌症中心的研究中, ,研究人员使用了一种多西环素诱导的KRAB–dCas9系统来筛选人类诱导多能干细胞(hiPS细胞)中的262个基因。他们发现76%的靶向翻译相关基因(262个中的200个)对生长是必需的,证明了该系统的有效性 [10].
这种微调基因表达的能力使CRISPRi和CRISPRa成为在功能基因组学研究中平衡细胞增殖和分化的宝贵工具。
适应性可逆筛选在培养肉应用中的应用
可逆基因调控为培养肉生产中的关键挑战提供了解决方案。例如,CRISPRa可以在高密度培养期间暂时激活参与营养物质运输或代谢途径的基因。一旦细胞达到所需密度,系统可以将基因表达恢复到正常水平,支持其正确分化为肌肉或脂肪组织。
诱导系统还可以将生物质扩增阶段与组织成熟分开。在规模化过程中,CRISPRi可以抑制与衰老相关的基因,有效地将牛细胞的增殖期从大约100天延长到超过200天[1]. 在获得足够的生物量后,研究人员可以恢复正常的基因表达以实现分化。这种方法对间充质干细胞特别有用,因为它们在培养中往往较早进入衰老阶段[1].
“针对这两个过程的基因编辑可以优化MSC扩展效率,同时保持其基本的多能性和分化潜力,最终推动可扩展的培养肉系统的发展。" – 通讯生物学 [1]
下表突出显示了可逆和永久基因调控方法之间的差异:
| 特征 | CRISPR 基因敲除 (KO) | CRISPRi / CRISPRa |
|---|---|---|
| DNA 修改 | 永久 (插入缺失) | 可逆 (转录) |
| 机制 | 双链断裂 | dCas9-效应器融合 |
| 最佳使用案例 | 识别关键基因 | 调节代谢/生长途径 |
| 分化风险 | 高 (永久功能丧失) | 低 (功能可恢复) |
此比较说明了如何定制可逆基因调控方法以应对开发用于培育肉生产的细胞系的特定挑战。
结合CRISPR筛选与细胞面板和基因分型技术
将CRISPR筛选与多组学分析相结合
将多组学和自动基因分型整合到CRISPR筛选中,特别是在推进培养肉细胞系开发方面,提高了其实用性。
将CRISPR筛选与多组学(如RNA测序)相结合,使研究人员能够绘制特定基因敲除对细胞通路的影响。这对于培养肉尤其重要,因为了解细胞如何平衡增殖和分化至关重要。
例如,针对牛脂肪来源的间充质干细胞, 的600个基因的组合CRISPR敲除筛选与RNA-seq配对,发现敲除TP53和PTEN延缓了衰老。这些细胞保持了年轻的基因表达特征,细胞周期基因上调,导致转导后第50天倍增率增加了50% [1].
像CROP-seq这样的单细胞平台更进一步,同时检测单个细胞中的sgRNA和转录组变化 [6] . 这种精确度对于识别增强肌肉分化或蛋白质合成的基因改造至关重要——这些是实现培养肉所需质地和营养特性 的关键因素。
另一种有前景的方法是细胞面板筛选,其中在来自不同供体、解剖部位和物种的多样化细胞系中测试CRISPR扰动。例如,研究人员在来自七个供体的人类肌母细胞系上验证了MyoCRISPR-KOLib文库。使用分裂毒素选择系统,他们识别了250个对肌母细胞融合至关重要的基因。其中,41个基因通过医学数据库确认在骨骼肌形态中发挥作用[6]. 这种多线验证确保了基因靶点在生物变异中保持稳健,这是扩大培养肉生产规模的关键考虑因素。
这些见解正在为自动化、可扩展的平台铺平道路,这些平台将基因筛选与详细基因分型结合用于工业应用。
集成平台中的自动化和可扩展性
自动化对于处理由集成CRISPR和基因分型平台生成的大量数据集和样本至关重要。RMCE系统能够实现无病毒、特定位点的sgRNA库传递,是向前迈出的重要一步。这些平台确保每个细胞接收一个一致的sgRNA拷贝,从而减少变异性。RMCE已经在中国仓鼠卵巢(CHO)细胞中展示了高文库覆盖率和最小偏差[5].
“一个无偏见的高通量基因筛选平台对于下一代CHO工厂的发展至关重要。” - 中国仓鼠卵巢研究团队[5]
通过像细胞适应性(CelFi)测定这样的验证工具,扩展性得到了进一步增强。该测定使用靶向深度测序来监测时间上的插入缺失(indel)谱,跟踪框内与框外突变的比例。通过将这些突变与生长优势或劣势相关联,研究人员可以有效验证培养肉细胞系中的基因靶点[2].
| 技术 | 整合方法 | 培养肉的主要优势 |
|---|---|---|
| RNA-seq / 多组学 | 将CRISPR命中与转录组学谱系联系起来 | 了解基因如何调节生长和分化[1][6] |
| 分裂毒素系统 | 将细胞融合与生存选择联系起来 | 定量选择具有融合能力或缺陷的细胞[6] |
| RMCE平台 | gRNA文库的位点特异性整合 | 高通量、无病毒筛选,具有一致的基因拷贝数[5] |
| CROP-seq | 单细胞CRISPR + RNA-seq | 同时检测sgRNA和转录组变化[6] |
| CelFi Assay | 靶向深度测序插入缺失突变 | 通过追踪等位基因频率变化快速验证遗传靶点[2] |
这些先进的平台简化了从识别遗传靶点到验证其对细胞适应性影响的过程。这种效率支持了足够强大的细胞系的发展,以进行大规模培养肉生产。
使用CRISPR筛选来改善细胞系的生长和增殖
CRISPR筛选方法已成为增强细胞系性能的强大工具,为培养肉生产提供了直接的好处。
CRISPR基础的细胞系改进示例
CRISPR筛选在培养肉研究中成功改善了细胞系性能。例如,在牛脂肪来源的间充质干细胞中,针对600个基因的混合敲除筛选识别出TP53和PTEN是生长的关键抑制因子。敲除TP53在30天内显著增加了细胞丰度[1]. 此外,编辑后的牛间充质干细胞平均显示出12%的更高倍增率[1].
通过靶向肿瘤抑制基因,研究人员将细胞的增殖寿命从大约100天延长到超过200天,有效地绕过了海夫利克极限。这种衰老的延迟使得生物质在工业相关的时间框架内得以扩展[1] .
在另一个例子中,由丁世杰、李春宝和周光宏领导的南京农业大学研究人员使用CRISPR/Cas9开发了CDKN2A−/−猪卫星细胞系。这些工程化细胞在定制的19组分无血清培养基(A19)中至少保持了18代的稳定增殖。它们还成功地播种到可食用支架, 上,创造出具有改善的咀嚼性和粘性的肉样结构[8]. 这些细胞在无血清条件下的多次传代中保持了超过90%的活力[8].
"基于CRISPR的CDKN2A敲除细胞提供了可再生的肌肉祖细胞来源,减少了对反复动物活检的依赖。"
这些例子突出了CRISPR筛选如何识别出能够提高生长速度、延缓细胞老化并实现无血清培养的基因改造——这三个方面对于扩大培养肉生产至关重要。
CRISPR优化细胞系的扩展挑战
虽然CRISPR优化的细胞系显示出明显的优势,但将其扩展到工业用途仍然面临挑战。增强的增殖通常以分化为代价。例如,TP53 在牛间充质干细胞中的敲除与肌肉分化基因的表达减少有关,这可能会阻碍其成熟为可食用组织的能力[1]. 为了解决这个问题,可能需要额外的策略,例如添加培养基补充剂或激活特定的转录因子,以在扩增后恢复分化[1].
另一个关键问题是维持遗传稳定性。基因拷贝数的变化(非整倍性)和CRISPR编辑期间的脱靶效应可能导致筛选研究中的结果不一致或假阳性[2]. 像细胞适应性(CelFi)测定这样的工具通过监测随时间变化的移码插入缺失的比例来帮助降低这些风险,确保观察到的生长优势直接与预期的编辑相关 [2].
经济和技术障碍仍然存在。间充质干细胞在培养肉行业中约占细胞来源的25%,面临着诸如生长因子成本高、需要优化的无血清培养基, 以及开发大规模生物反应器(10,000–50,000 L容量)[1][9][11]. 此外,确保细胞在3D支架上播种时的期望质地仍然是一项复杂的任务[11].
“目前的培养肉面临重大挑战,包括高成本、规模化问题以及对进一步技术进步的需求。"
- 通讯生物学 [1]
克服这些挑战需要一种综合方法,将基因优化与培养基配方、生物反应器技术和分化协议的进步相结合。虽然CRISPR筛选提供了关键的基因见解,但将这些发现转化为可扩展的解决方案需要集成系统和严格的验证过程。这些努力对于将培养肉生产从实验室推向商业可行性至关重要。
如何Cellbase 支持培养肉中的CRISPR研究
CRISPR筛选已经显示出其潜力,但将其扩展到工业用途需要获得专业工具和资源。这就是
通过 Cellbase 获取CRISPR资源
与广谱制药供应商不同,
在2025年11月,
)连接起来,解决了供应链碎片化的问题。“我们与之交谈的每一家培养肉公司都在同样的采购难题上浪费时间。寻找关键组件的供应商意味着要在不理解食品应用的制药供应商中进行谷歌搜索。”
- Cultigen Group创始人David Bell表示。 [15]
通过集中这些资源,
在培育肉类开发中促进合作
该平台旨在处理大型商业项目的需求,例如 Believer Meats 和 Aleph Farms. 这些项目需要50,000升生物反应器的基础设施和 优化的生产供应链, ,
结论
高通量CRISPR筛选已从一个有前途的概念转变为推进培养肉类开发的关键工具。这项技术在优化细胞系方面的影响是不可否认的。例如,最近的突破显示,基因改造可以将牛干细胞的增殖寿命从100天延长到200天,将衰老细胞群体从60%减少到仅10%,并在一个月内实现细胞丰度惊人的1000倍增长[1]. 这些进展标志着从实验研究到实际工业应用的明确转变。
紧凑的平台和针对性的文库正在解决该领域一些最紧迫的挑战。数字微流体系统现在允许每个条件仅用3,000个细胞进行筛选,使得使用有限的、非商业可用的初级动物细胞成为可能。同时,像 MyoCRISPR-KOLib 这样的专注库能够有效地靶向 90% 的相关转录本,同时仅覆盖三分之一的基因组[3][6] . 这种精确性和效率对于克服资源限制和扩大生产规模.
"这些发现展示了 CRISPR 筛选在优化牛干细胞特性方面的实用性,并为未来更大规模的培养肉生产提供了一条路径。" [1]
尽管有这些进展,成功仍然取决于获得合适的基础设施。研究人员需要特定物种的 gRNA 库,为食品应用设计的培养基, 兼容的生物反应器,以及专为培养肉生产而非制药用途量身定制的分析工具。为了解决这些需求,
对于致力于开发下一代培养肉细胞系的团队,工具和技术已经准备就绪。现在的挑战在于快速有效地部署CRISPR筛选,以实现其全部潜力。
常见问题解答
如何在CRISPR敲除、CRISPRi和CRISPRa之间选择用于筛选?
这些系统之间的选择取决于您的具体生物学问题和您所追求的结果:
- CRISPR敲除: 这种方法完全破坏基因功能,非常适合研究基因丧失或失活的影响。
- CRISPRi: 通过在不切割DNA的情况下抑制基因表达,这种方法非常适合研究必需基因或需要可逆抑制的情况。
- CRISPRa : 如果您需要上调基因表达,该系统是首选。它特别适用于研究过表达的影响,例如促进细胞增殖或分化。
在做决定时,请考虑您的细胞模型、目标基因以及实验的整体目标。
如何在不损害肌肉或脂肪分化的情况下促进增殖?
在保持肌肉或脂肪细胞分化能力的同时促进其增殖是培养肉生产中的一个关键挑战。一种有前景的方法涉及基于CRISPR的基因编辑, ,它允许精确操控基因以增强生长或延长细胞寿命。例如,靶向肌生成抑制素 (MSTN)可以促进细胞生长,而编辑CDKN2A有助于细胞绕过衰老。
话虽如此,实现增殖和分化之间的平衡至关重要。对某些靶标的管理不当,例如P53 (TP53), 可能会损害分化,可能会影响组织质量。为了应对这些复杂性,高通量CRISPR筛选是至关重要的。这项技术识别出最有效的基因调节因子,为规模化和健康的培养肉生产中的组织发展铺平了道路。
在扩大细胞系之前,需要验证CRISPR筛选命中的哪些内容?
验证用于培养肉生产的CRISPR筛选命中需要一种系统的方法。首先,必须通过独立实验(如基因敲除)确认基因功能,以确保观察到的效果是可重复的。接下来,评估这些基因的生物学相关性至关重要,需检查它们对细胞增殖、活力和寿命等因素的影响。
安全评估同样重要,以排除可能影响过程的非目标效应或遗传不稳定性。在模拟工业环境(如生物反应器)的条件下进行功能验证是另一个关键步骤。这确保了基因编辑在大规模生产环境中按预期执行。在考虑扩大规模之前,每个阶段的彻底测试都是不可或缺的。









