

Wenn Sie Zellen expandieren können, sie aber nicht zur richtigen Zeit in das richtige Schicksal überführen können, wird Ihr Prozess bei der Differenzierung ins Stocken geraten. Das ist der Kernpunkt hier: Synthetische Genkreise geben Ihnen innerhalb der Zelle Kontrolle über Verpflichtung, Timing, Gedächtnis und Abstammungsmischung, wo Medienwechsel allein oft heterogene, teilweise verpflichtete Populationen hinterlassen.
Wenn ich einen Workflow zur Differenzierung von kultiviertem Fleisch aufbauen würde, würde ich sofort vier Punkte aus diesem Artikel übernehmen:
- Beginnen Sie mit dem nativen Netzwerk, nicht mit dem Konstrukt. Verwenden Sie snRNA-seq, Trajektorienanalyse, GRN-Inferenz und miRNA-Profiling, um herauszufinden, wo Zellen ins Stocken geraten, abdriften oder in das falsche Schicksal verzweigen.
-
Passen Sie den Schaltungstyp an das Prozessproblem an.
Ein Umschalter eignet sich für Lock-in, ein Feedforward- oder Bandpass-Design eignet sich für die Zeitsteuerung, ein Logikgatter eignet sich für die Mehrsignalsteuerung, und miSFITs eignen sich für abgestufte Ausgaben. - Entwerfen Sie von Anfang an für geringe Leckage, geringe Geräuschentwicklung und Sicherheit. Orthogonale Teile, negative Autoregulation, iFFLs, cm-Transgene und ein induzierbares Kill- oder Wachstumsstopp-Modul sind Teil des Aufbaus, keine nachträgliche Überlegung.
- Validieren Sie frühzeitig unter maßstabsrelevanten Bedingungen. Ein Schaltkreis, der in 2D funktioniert, kann sich in 3D, Mikrokörper oder gerührter Suspension aufgrund von Induktorgradienten, Sauerstoffgrenzen und Scherung verschieben.
Der Artikel macht auch einen praktischen Punkt, der für Prozessteams wichtig ist: Einzelstammsteuerung und Verhältnissteuerung sind unterschiedliche Aufgaben. Ein Tet-On MyoD-Kassette kann den myogenen Eintritt fördern, aber Whole-Cut-Produkte benötigen die Kontrolle der Muskeln, Fett- und ECM-Anteile, , was normalerweise Feedback, parakrine Signalgebung und intensiveres Klonscreening bedeutet.
Einige Zahlen verankern diese Botschaft:
- Die Standard-Myogenese-Differenzierung kann bei Fusionsindizes von etwa 50–60% ins Stocken geraten
- Engineered GRNs in iPSCs erhöhten die Ziel-Linien-Differenzierung von 52% auf 81%
- Synthetische Schaltkreise in modifizierten MSCs führten zu einer kardialen Differenzierung von 76%
- Einige porcine Tet-On-PAX7-Linien behielten ein hohes myogenes Potenzial über 40 Passagen hinaus
- Rund 20% der menschlichen pluripotenten Stammzellen können krebsassoziierte Mutationen tragen, weshalb induzierbare Sicherheitsmodule wichtig sind
Synthetischer Gen-Schaltkreis-Workflow für die Differenzierung von kultiviertem Fleisch
Wissenschaftlergeschichten: Michael Elowitz, Genetische Schaltkreise in lebenden Zellen
Schneller Vergleich
| Thema | Was der Artikel in einfachen Worten sagt |
|---|---|
| Hauptengpass | Differenzierung, nicht Expansion |
| Warum Schaltkreise helfen | Sie fügen Schwellenwertkontrolle und Zellzustandsspeicher innerhalb der Zelle hinzu |
| Bester erster Schritt | Natürliche Schicksalskontrolle mit snRNA-seq und verwandten Werkzeugen kartieren |
| Hauptschaltkreisoptionen | Toggle, Feedforward, Bandpass, Logikgatter, miSFITs, CRISPRa/i |
| Myogenese-Beispiel | Tet-On-MyoD, um Wachstum von terminaler Differenzierung zu trennen |
| Adipogenese / ECM-Kontrolle | miSFITs und Bow-Tie-Designs für abgestufte PPARG/BMP4-Ausgabe |
| Whole-Cut-Herausforderung | Verhältnissteuerung über Muskel-, Fett- und Bindegewebe |
| Skalierungsrisiko | 2D-Verhalten hält möglicherweise nicht in 3D oder Bioreaktoren stand |
| Integrationsoptionen | Lentivirus, Transposons, CRISPR Knock-in, episomale Vektoren |
| Regulatorischer Punkt | Engineerte Linien benötigen ein umfassenderes Sicherheitsprofil; lebensmittelsichere Induktoren wie Vanillinsäure werden gegenüber DOX bevorzugt, wo möglich |
Also, einfach ausgedrückt: dies ist nicht nur ein Artikel über Schaltungsdesign. Ich würde es als Leitfaden lesen, um Schaltungsarchitektur, Abstammungsbiologie, Klonauswahl, Bioreaktorleistung und UK/EU-Sicherheitsdokumentation in eine Differenzierungsstrategie zu integrieren.
Lesen Sie weiter, wenn Sie den vollständigen Weg von der nativen Netzwerkabbildung zur Konstruktionswahl, Skalierungsprüfungen und regulatorischer Eignung erfahren möchten.
2. Gestaltungsprinzipien für Differenzierungskontrollschaltungen
2.1 Kartieren Sie das native Zellschicksalsnetzwerk, bevor Sie die Schaltung entwerfen
Bevor Sie eine Schaltung entwerfen, benötigen Sie ein klares Bild davon, was die Zelle bereits tut.
Einzelkern-RNA-Sequenzierung (snRNA-seq) ist ein guter Ausgangspunkt. Sie kann ruhende Subpopulationen zeigen, einschließlich Reservezellen, die durch NOTCH2 und HEYL , markiert sind, und auf Signalwegziele hinweisen, die die Differenzierung verbessern könnten [3].
Von dort aus helfen Trajektorienanalyse und Genregulationsnetzwerk (GRN)-Ableitung, die Reihenfolge der Regulatoraktivierung zu kartieren und hervorzuheben, wo Zellen am wahrscheinlichsten ins Stocken geraten. In der Myogenese verläuft die Hauptkaskade über MYOD1 und MYOG . In der Adipogenese sind die Hauptknoten PPARG und CEBPA , wobei das Schicksal der fibro-adipogenen Vorläuferzellen (FAP) als Hauptverzweigungspunkt-Risiko fungiert. Die folgende Tabelle fasst die Hauptregulatoren und Engpässe zusammen.
| Abstammung | Wichtige Hauptregulatoren | Kritische Signalwege | Identifizierte Engpässe |
|---|---|---|---|
| Myogen | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Reservezellbildung (Ruhezustand) |
| Adipogen | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Fibro-adipogene Vorläufer (FAP) Schicksal |
| Pluripotent | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Spontane Differenzierung / Heterogenität |
Eine weitere nützliche Ebene ist miRNA-Expressionsprofilierung. Endogene miRNAs wie miR-302a, verknüpft mit Pluripotenz, und miR-375, verknüpft mit Differenzierung, können als interne Klassifikatoren in Sense-and-Respond-Designs fungieren. Das ermöglicht es dem Schaltkreis, den tatsächlichen Zustand der Zelle zu lesen, anstatt sich nur auf externe Induktoren zu verlassen [5].
Diese Engpässe sollten die Schaltkreisauswahl bestimmen. Wenn das Hauptproblem Drift ist, benötigen Sie möglicherweise einen Lock-In. Wenn das Timing das Problem ist, könnte ein Puls besser passen. Wenn die Schicksalskontrolle von mehr als einem Signal abhängt, macht Multi-Input-Logik normalerweise mehr Sinn.
2.2 Wählen Sie die richtige Schaltungsarchitektur
Hier werden die Kompromisse deutlich sichtbar. Die richtige Architektur hängt von drei praktischen Punkten ab: wie dauerhaft das Engagement sein muss, wie genau das Timing kontrolliert werden muss und wie viel genetische Nutzlast die Zelle ohne Probleme tragen kann.
Bistabile Kippschalter eignen sich für Fälle, in denen die Linienbindung gesperrt bleiben muss. Das Hauptproblem ist das spontane Umschalten des Zustands, verursacht durch transkriptionelles Rauschen.
Bandpassfilter eignen sich für Fälle, in denen ein Transkriptionsfaktor nur innerhalb eines definierten Entwicklungsfensters benötigt wird. Der Haken ist, dass die Induktorlevel streng kontrolliert werden müssen, sonst verschiebt sich das Timing.
Logikgatter erhöhen die Spezifität, indem sie mehr als einen Eingang gleichzeitig erfordern. Zum Beispiel kann die Differenzierung nur erlaubt werden, wenn ein exogener Induktor vorhanden ist und die Zelle das richtige endogene miRNA-Profil zeigt. Das hilft, das Risiko einer Fehlverpflichtung zu verringern.
Die folgende Tabelle skizziert die Hauptarchitekturen und die damit verbundenen Kompromisse.
| Architektur | Reversibilität | Zeitliche Präzision | Integrationskomplexität | Primärer Anwendungsfall | Hauptsächliches Risiko |
|---|---|---|---|---|---|
| Bistabiler Schalter | Niedrig (einmal gesperrt) | Moderat | Moderat | Permanente Abstammungsverpflichtung | Spontanes Umschalten aufgrund von Rauschen |
| Bandpassfilter | Hoch (konzentrationsabhängig) | Hoch | Hoch | Vorübergehende Entwicklungsstadien | Erfordert strenge Induktorsteuerung |
| Logikgatter (UND/ODER/NICHT) | Variabel | Moderat | Moderat–hoch | Zelltypspezifische Aktivierung | Leckage in AUS-Zuständen |
| Fliege / Mehrfacheingabe | Hoch | Moderat | Moderat | Integration mehrerer Signale | Abhängig von der Stabilität endogener miRNA |
| miSFITs | Hoch | Moderat | Niedrig–moderat | Gestufte Ausgangssteuerung | Enger Dynamikbereich bei schlechter Abstimmung |
"Durch die Minimierung der Anzahl der Berechnungsschichten bei gleichzeitiger Beibehaltung der Funktionalität adressiert diese Strategie Skalierbarkeitsbarrieren im Bereich der Genkreiskonstruktion." - Nature Communications [9]
Jede hinzugefügte regulatorische Schicht erhöht die Nutzlast und beansprucht zelluläre Ressourcen. In der Praxis gilt: Wenn zwei Designs die gleiche Aufgabe erfüllen, ist das einfachere in der Regel die bessere Wahl, wenn es auf Skalierung ankommt.
Sobald die Architektur festgelegt ist, besteht die nächste Aufgabe darin, sie unter geringer Leckage, Rauschunterdrückung und ausfallsicherer Steuerung zu stabilisieren.
2.3 Bauen für Zuverlässigkeit, geringe Leckage und Sicherheit
Ein Schaltkreis muss über eine längere Kultur stabil bleiben. Eine kurze Phase guter Leistung reicht für den Produktionseinsatz nicht aus.
Orthogonale Teile sind die erste Verteidigung. Promotoren, Transkriptionsfaktoren und regulatorische Elemente, die nicht mit der nativen Maschinerie kreuzreagieren, helfen, Off-Target-Effekte zu begrenzen und die Wahrscheinlichkeit zu verringern, dass endogene Signale den Schaltkreis einschalten oder abschalten."Modifizierte hochdichte Promotoren wie PCREm wurden verwendet, um die basale Leckage in induzierbaren Säugetiersystemen zu reduzieren [6] .
Negative Autoregulation ist ebenfalls sinnvoll, wo immer möglich. Es ist eines der bekannteren Motive zur Reduzierung von transkriptionalem Rauschen und zur Erzeugung einer lineareren Reaktion auf die Induktorenkonzentration [6] . Inkoherente Feedforward-Schleifen (iFFLs) können eine weitere Kontrollschicht hinzufügen, indem sie stochastische Schwankungen filtern, sodass Zellen auf anhaltende Signale anstatt auf kurze Rauschspitzen reagieren.
Kodon-modifizierte ( cm) Versionen von synthetischen Transkriptionsfaktoren erleichtern ebenfalls die Charakterisierung. Sie ermöglichen es, die durch Schaltkreise gesteuerte Expression von der endogenen genomischen (g) Expression während der Validierung zu trennen [1]. Das mag wie ein kleines Detail erscheinen, aber es spart Zeit, wenn Sie versuchen herauszufinden, ob eine Anzeige vom Schaltkreis oder dem Wirtsgenom stammt.
Sicherheitsmodule sind erforderlich. Ungefähr 20% der menschlichen pluripotenten Stammzellen tragen krebsassoziierte Mutationen [7] . Wenn also ein Schaltkreis in eine aus Stammzellen abgeleitete Linie eingebracht wird, sollte er ein induzierbares Wachstumsstopp- oder Eliminierungsmodul enthalten. Vanillinsäure ist hier ein nützlicher Induktor, da es ein zugelassener Lebensmittelzusatzstoff ist, was den Einsatz als Schaltkreisauslöser in kultivierten Fleischzelllinien unterstützt [1] .
"Die synthetische Biologie bietet Ingenieuren die Mittel, Schaltkreise einzusetzen, um die Expression mehrerer Gene einfach und präzise abzustimmen, um... potenzielle unerwünschte Off-Target-Nebenwirkungen zu eliminieren." - npj Systems Biology and Applications [6]
Diese Entscheidungen richten die abstammungsspezifischen Schaltkreise in Abschnitt 3 ein.
sbb-itb-ffee270
3. Schaltkreisstrategien für myogene, adipogene und verhältnisgesteuerte Differenzierung
3.1 Myogene Schaltkreise, die Expansion von terminaler Differenzierung trennen
Sobald die Schaltkreisarchitektur festgelegt ist, besteht die nächste Aufgabe in der abstammungsspezifischen Bereitstellung. Für die Myogenese ist das Hauptproblem einfach zu formulieren, aber schwer auszuführen: Zellen müssen zuerst proliferieren und dann auf Anforderung in die Differenzierung übergehen, ohne zu früh in eine der beiden Richtungen abzudriften.
Ein Tet-On-MyoD-Kassette ist eine der direktesten Möglichkeiten, dies zu tun. In diesem Setup proliferieren Zellen unter Standardbedingungen, wenn Doxycyclin (DOX) fehlt. Fügen Sie DOX hinzu, und der Schaltkreis treibt die myogene Verpflichtung voran.Forscher an mehreren chinesischen Institutionen verwendeten diesen Ansatz in Hühnerembryo-Fibroblasten und berichteten über eine effiziente Myotubenbildung nach Induktion [4] .
Ein bistabiler Kippschalter bietet eine straffere Zustandskontrolle. Systeme, die aus sich gegenseitig hemmenden Repressoren wie E-KRAB und Pip-KRAB bestehen, können einen kurzen DOX-Impuls in ein stabiles myogenes Programm umwandeln [6] . In der Praxis bedeutet das, dass der differenzierte Zustand nach der Induktion beibehalten wird, anstatt zu verblassen, sobald das Signal entfernt wird. Die Hinzufügung negativer Autoregulation hilft auch, stochastisches Rauschen zu reduzieren und undichte Differenzierung während der Expansionsphase zu begrenzen [6].
Dies ist wichtig, weil die standardmäßige in vitro myogene Differenzierung oft auf halbem Weg ins Stocken gerät. Die Fusionsindizes liegen normalerweise nur bei etwa 50–60 %, was eine große nicht-fusionierende Reservepopulation hinterlässt [3]. Schaltungsgetriebene MyoD-Aktivierung kann das Engagement verbessern, aber Engagement allein garantiert keine einheitliche Fusion. Wenn MyoD-Schaltungen mit MEK-, NOTCH- und RXR-Modulation kombiniert werden, wird eine nahezu vollständige Fusion in 2D-Kultur möglich [3]. Die Schaltung steuert das Timing des Linien-Eintritts; die kleinen Moleküle fördern einen saubereren und gleichmäßigeren Zellzyklus-Austritt.
3.2 Adipogene und fibrogene Kontrolle für Zusammensetzung und Struktur
Muskel allein ist nicht genug. Sobald die myogene Kontrolle eingerichtet ist, stellt sich die nächste Frage der Zusammensetzung: wie viel Fett sich bildet, wie viel ECM abgelagert wird und wann diese Programme aktiviert werden. Hier ist die binäre EIN/AUS-Steuerung oft zu grob. Was Teams normalerweise brauchen, ist ein abgestufter Ausgang, insbesondere um den adipogenen-fibrogenen Verzweigungspunkt.
miSFITs bieten eine praktische Möglichkeit, die Expression in Schritten zu steuern.Durch das Platzieren mutierter miRNA-Zielstellen - zum Beispiel Stellen für miR-17 - in der 3′UTR von Ausgabegenen wie PPARG oder BMP4 können Forscher Ausdrucksniveaus aus einer Variantenbibliothek auswählen [5]. Das macht die Adipozyteninduktion eher wie einen Dimmer als einen Lichtschalter. Anstatt Zellen in eine Alles-oder-Nichts-Reaktion zu drängen, können Teams die Adipogenese sorgfältiger titrieren [5].
Fibroblasten sind hier auch nicht nur Zuschauer. Sie liefern ECM-Proteine, die die Textur formen [10]. Das macht die fibrogene Kontrolle zu einem Teil des Produktdesigns und nicht nur zu einem Nebenthema. Schaltkreise können helfen, den Wechsel zwischen fibrogenen und adipogenen Zuständen zu steuern, und in kultiviertem Geflügel kann eine direkte PPARG-Aktivierung in Fibroblasten erforderlich sein, um eine bedeutende Fettablagerung zu erzeugen [10].
Eine Bow-Tie-Architektur passt gut zu diesem Problem, da sie das Erfassen und die Ausgabe getrennt hält. Die Erfassungsschicht liest den aktuellen Zustand der Zelle, während die Ausgabeschicht PPARG, CEBPA oder andere Linienregulatoren anpasst. Diese Trennung hilft, adipogene oder Bindegewebsprogramme davon abzuhalten, sich einzuschalten, bevor die Zellen das richtige Entwicklungsstadium erreicht haben.
3.3 Kontrolle des Multi-Linien-Verhältnisses und Feedback-Erfassung
Verhältnis-Kontrollkreise befassen sich mit einem anderen Problem: nicht ob die Differenzierung stattfindet, sondern ob die endgültige Populationsmischung dort bleibt, wo sie sein sollte. Für Whole-Cut-Produkte ist es genauso wichtig, Muskel, Fett und ECM in die richtigen Proportionen zu bringen, wie eine einzelne Linie zur Differenzierung zu bringen.
Diese Systeme bauen Feedback-Kontrolle in die Zellen selbst ein. Zustandspezifische Promotoren beschränken die Expression von Signalproteinen auf Zellen, die sich bereits einer bestimmten Linie verpflichtet haben.Synthetische parakrine Module lassen dann engagierte myogene Zellen ein hemmendes Signal freisetzen, das die adipogene Verpflichtung in nahegelegenen Zellen unterdrückt. Die Logik ähnelt der lateralen Hemmung in Delta-Notch-Entwicklungssystemen [1] [6]. Wo die Verzweigung komplizierter wird, können Mehrfacheingangstore extrazelluläre Signale mit internen Zustandsignalen kombinieren [9].
miSFITs funktionieren auch in dieser Phase. Durch das Abstimmen der Ausgabestärke von BMP4 oder anderen Morphogenen können Teams das Liniengleichgewicht verschieben, ohne die vorgelagerte Entscheidungslogik neu zu schreiben. In konstruierten genregulatorischen Netzwerken erhöhte diese Art der Kontrolle die Effizienz der Ziel-Linien-Differenzierung von 52 % in Kontrollen auf 81 % in konstruierten iPSCs [2]. In modifizierten mesenchymalen Stammzellen trieben synthetische Schaltkreise die kardiale Differenzierungseffizienz auf 76 % [2].
Die folgende Tabelle vergleicht die Ansätze der Einzelabstammung und der Verhältnissteuerung anhand der wichtigsten Punkte in der Produktion.
| Funktion | Einzelstamm-Schaltung | Verhältnissteuerungsschaltung |
|---|---|---|
| Komplexität | Niedrig; typischerweise ein einzelner induzierbarer Promotor und Regulator [4] | Hoch; erfordert Logikgatter und parakrine Signalgebung [6][9] |
| Überwachungsaufwand | Niedrig; folgt normalerweise einem einzelnen Reporter [4] | Hoch; erfordert die Verfolgung mehrerer Abstammungsmarker [5] |
| Robustheit | Moderat; anfällig für Heterogenität und Reservezellbildung [3] | Hoch; nutzt Feedback und laterale Hemmung, um Populationen auszugleichen [1] |
| Produktionswert | Hoch für Biomasse; begrenzt für komplexe Gewebestruktur [10] | Wesentlich für Ganzschnittprodukte, die Muskel, Fett und ECM benötigen [4] |
Verhältnissteuerkreise erhöhen die Validierungslast. Aber ihr eingebautes Feedback ist schwer allein mit der Prozesskontrolle zu erreichen, was mehr Druck auf die Klonauswahl und das Prozesstesten ausübt.
4. Vom Konstrukt zum Prozess: Validierung, Hochskalierung und regulatorische Eignung
4.1 Integrationsstrategie und Klonauswahl für stabile Leistung
Nach dem Schaltungsdesign beginnt der schwierige Teil: Diese Schaltung in Zellen zu bringen, auf eine Weise, die durch die Produktion im großen Maßstab stabil bleibt.
Lentivirale Lieferung ist oft effizient und liefert schnell stabile Integranten. Aber die Integration ist zufällig. Das bedeutet mehr regulatorische Aufmerksamkeit und ein Risiko, dass der Ausdruck im Laufe der Zeit aufgrund von Silencing abnimmt. Transposonsysteme wie PiggyBac und Sleeping Beauty liegen in der Mitte. Sie können die Leistung über viele Passagen hinweg halten, aber man muss dennoch die Kopienzahl und die Einfügestellen überprüfen. Dornröschen, zum Beispiel wurde verwendet, um bovine Satellitenzellen stabil zu immortalieren, indem TERT und CDK4 überexprimiert wurden, wobei Linien ihr myogenes Potenzial über 40 Passagen hinaus beibehalten [10] . CRISPR-Knock-in bietet die genaueste Kontrolle darüber, wo das Konstrukt landet, und die präziseste genomische Einstellung, obwohl die Klonauswahl langsamer ist und der Durchsatz geringer ist.
| Integrationsmethode | Einfügekontrolle | Stabilität | Skalierbarkeit | Regulatorische Überlegungen |
|---|---|---|---|---|
| Lentivirale Lieferung | Niedrig (zufällige Integration) | Hoch, aber anfällig für Silencing | Hoch | Erhöhte regulatorische Prüfung aufgrund zufälliger Einfügung und viraler Überreste |
| Transposons (PiggyBac/SB) | Moderat | Hoch über viele Passagen | Hoch | Erfordert Screening auf Kopienzahl und Einfügestellen |
| CRISPR Knock-in | Hoch (ortspezifisch) | Sehr hoch | Moderat | Günstig; reduziert das Risiko der Störung endogener Gene |
| Episomale Vektoren | Keine (extrachromosomal) | Niedrig; kann während der Teilung verloren gehen | Niedrig | Niedrigere Integrationshürde, aber ungeeignet für langfristige Expansion |
Klonscreening muss mehr tun, als nur das Vorhandensein des Konstrukts zu bestätigen.Es sollte Expressionsdrift, Insertionsprofil, Wachstumskinetik, Differenzierungseffizienz und Phänotyperhalt bei hoher Passagenzahl verfolgen. snRNA-seq ist hier nützlich, da es Klone ausschließen kann, die für Pax7⁺/Ki-67⁻ Reservezellen angereichert sind - Zellen, die den Zellzyklus verlassen, ohne sich zu differenzieren - bevor das Hochskalieren erfolgt [3]. Porcine EPSCs mit einem Tet-On-PAX7-Schaltkreis behielten eine hohe Muskeldifferenzierung in 3D-Mikroträger- und Suspensionskultur über 40 Passagen bei [8].
4.2 Wie sich das Schaltkreisverhalten in 3D, Mikroträger- und Bioreaktorkultur verändert
Sobald Sie einen Klon haben, besteht der nächste Test darin, ob er sich außerhalb von 2D gleich verhält. In vielen Fällen tut er das nicht. Die Leistung in 2D überträgt sich selten sauber auf Suspension, Mikroträger oder Gerüstkultur, da Diffusionsgradienten, Sauerstoffgrenzen und Scherung die Schaltkreisausgabe verändern.
Einer der ersten Prüfungen ist die Induktor-Diffusion. In Rührkessel-Bioreaktoren müssen kleine Molekül-Induktoren die Zellen gleichmäßig erreichen. In der Praxis können sich Gradienten bilden, insbesondere in dichten Mikrokulturträgerkulturen und innerhalb von Aggregaten oder Gerüstkernen. Suspensionskultur ist normalerweise besser geeignet für großangelegtes kultiviertes Fleisch, da sie eine höhere Zelldichte unterstützt und eine engere Prozesskontrolle ermöglicht.
Die Überwachung des Zellzustands wird ebenfalls schwieriger, wenn das System skaliert. Fluoreszierende Reporter-Signale, die in 2D leicht mikroskopisch zu lesen sind, können in undurchsichtigen 3D-Konstrukten schwer zu erkennen sein. Fluoreszierende Timer - Sonden, die die Emission von Blau zu Rot verschieben, wenn ein Protein reift - können Echtzeit-Daten zur Schaltkreisaktivierung in situ liefern [1]. Der vernünftige Weg ist eine gestufte Validierung: zuerst in 2D, dann in 3D-Konstrukten, und schließlich unter den endgültigen Bioreaktorbedingungen [3] [8].
4.3 Charakterisierung, Lebensmittelsicherheitsdokumentation und UK/EU-Erwägungen
Nach dem Prozesstest muss die Charakterisierung zeigen, dass Schaltkreisfunktion, Phänotyp und Sicherheit weiterhin bestehen. Der Kerndatenstapel sollte Durchflusszytometrie, qPCR mit cm-Sequenzen, RNA-seq-Zeitverläufe und funktionale Auslesungen wie Myosin-Schwerketten-Flächenanteil und Myoglobin-Expression umfassen [1]. Optimierte serumfreie Differenzierungsmedien haben gezeigt, dass die Myoglobin-Expression auf etwa 30 % der in nativem Rindfleischmuskel gefundenen Werte gebracht werden kann [3]. Das gibt den Teams einen klaren Maßstab anstelle eines vagen Ziels.
Sie müssen auch Protein-, Aminosäure- und Fettprofile sowie sensorische Eigenschaften dokumentieren [10][3].
Aus regulatorischer Sicht ziehen das Vereinigte Königreich und die EU eine klare Linie zwischen spontan immortalisierten (nicht-GVO) Zelllinien und genetisch veränderten Linien. Letztere benötigen umfassendere Sicherheitsdossiers [10][3]. Stabilitätspakete sollten den Phänotyperhalt und die genomische Stabilität über die gesamte Produktionsbrücke hinweg zeigen - von der Masterzellbank bis zu den endgültigen Produktionszellen - und Rückverfolgbarkeitsaufzeichnungen müssen jeden Zwischenschritt berücksichtigen [10]. Wenn der Schaltkreis von einem chemischen Induktor abhängt, ist ein lebensmittelsicherer oder zugelassener Zusatzstoff wie Vanillinsäure gegenüber Doxycyclin vorzuziehen [1].
Routinemäßige genomische Überwachung ist ein Muss, und ein induzierbarer Selbstmord- oder Eliminationsschalter sollte als zentrales Risikokontrollmaß dokumentiert werden [7]. Seine Funktion sollte auch im Sicherheitsdossier enthalten sein, insbesondere da die Vorschriften für kultiviertes Fleisch im Vereinigten Königreich und in der EU weiter Gestalt annehmen.
5. Praktische Roadmap und Fazit
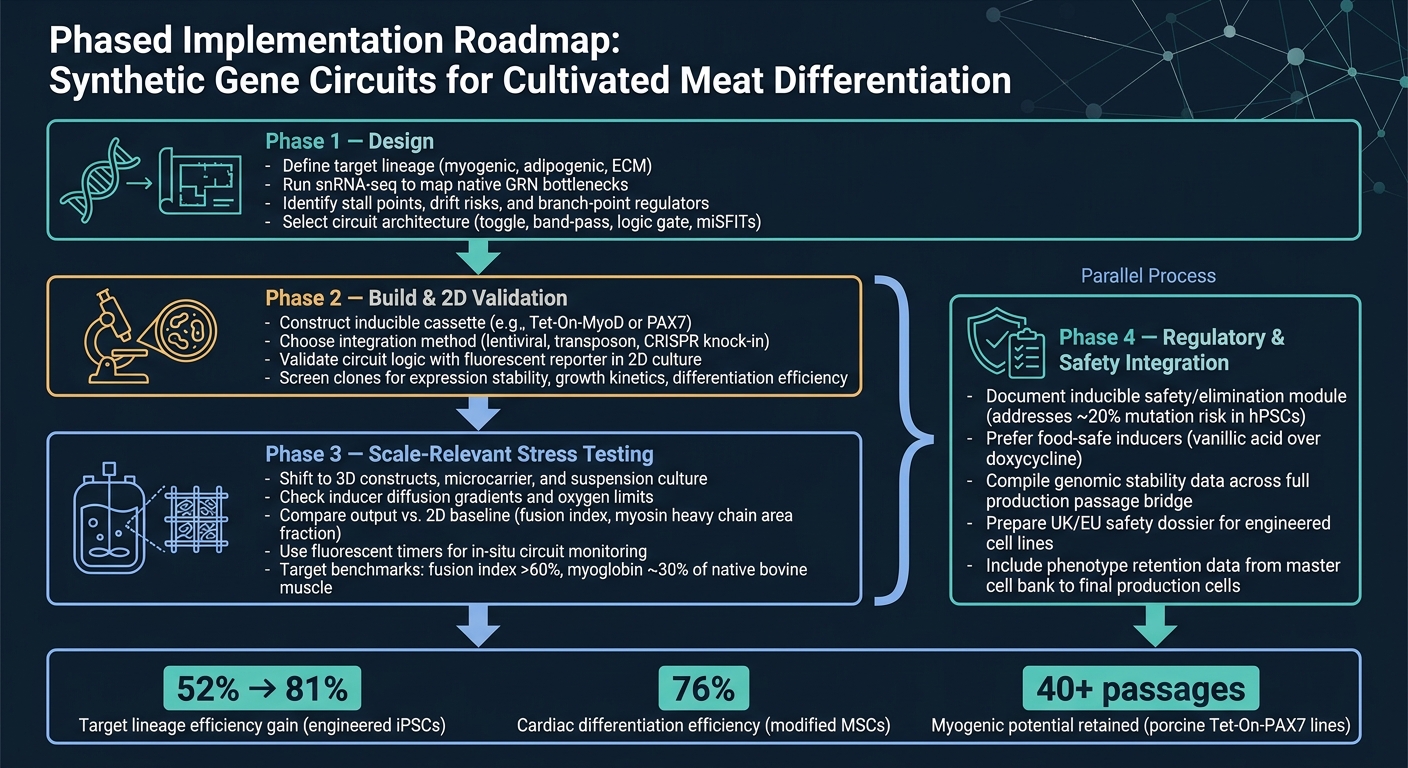
5.1 Ein phasenweiser Implementierungsfahrplan für Teams im Bereich kultiviertes Fleisch
Der sauberste Weg vom Konzept zur Produktion ist ein gestufter Arbeitsablauf.
Phase 1 ist das Design. Beginnen Sie mit der Definition der Zielabstammung und verwenden Sie dann snRNA-seq, um die Hauptengpässe zu bestätigen, bevor Sie eine Schaltungsarchitektur auswählen. Dieser Schritt ist wichtig, da eine Schaltung nur die Einschränkungen lösen kann, die Sie tatsächlich identifiziert haben.
Phase 2 ist der Bau und die 2D-Validierung. Bauen Sie das Konstrukt und überprüfen Sie, ob die Schaltung in 2D wie beabsichtigt funktioniert, indem Sie eine einfache Reporter-Auslesung verwenden.In diesem Stadium ist das Ziel einfach: Bestätigen Sie, dass die Logik funktioniert, bevor Sie zu schwierigeren und kostspieligeren Modellen übergehen.
Phase 3 ist maßstabsrelevantes Stresstesten. Wechseln Sie zu 3D-Systemen und bioreaktor-relevanten Bedingungen und vergleichen Sie dann die Ausgabe mit der 2D-Basislinie. Hier zeigen viele Designs ihre Schwachstellen, insbesondere wenn Massenübertragung, Scherung und Matrixeffekte ins Spiel kommen.
Phase 4 ist die Integration von Regulierung und Sicherheit und sollte parallel zu Phase 3 ablaufen. Sicherheits- und Regulierungsarbeiten sollten nicht bis zum Ende warten. Führen Sie sie parallel zur Skalierung durch, einschließlich der Dokumentation für jedes induzierbare Sicherheitsmodul.
5.2 Beschaffung von unterstützenden Werkzeugen und Materialien durch Cellbase
Sobald der Arbeitsablauf festgelegt ist, wird die Beschaffung oft zum geschwindigkeitsbestimmenden Schritt.
- Zelllinien
- serumfreie und chemisch definierte Medien
- Gerüste
- Bioreaktorkomponenten
- Sensoren
- Analytische Geräte
Zuverlässiger Zugang zu kompatiblen Materialien in jeder Phase hat einen direkten Einfluss darauf, wie schnell das Schaltkreisverhalten unter maßstabsrelevanten Bedingungen charakterisiert werden kann.
5.3 Wichtige Erkenntnisse
Synthetische Genschaltkreise geben Teams für kultiviertes Fleisch programmierbare Kontrolle über Timing, Schwellenwerte und Liniengleichgewicht, die Medien-Only-Protokolle nicht erreichen können. Die Wahl der Architektur beeinflusst Umkehrbarkeit, Leckage und Sicherheit.Induzierbare Systeme werden in der Regel bevorzugt, da sie eine bedingte Kontrolle und eine geringere metabolische Belastung bieten [6].
"Das synthetische Biologie-Toolkit kann verwendet werden, um Zelllinien mit abstimmbarer Genexpression zu etablieren, die in Verbindung mit PAT und computergestützter Modellierung geschlossene Regelkreissysteme ermöglichen können, um optimalen Produktertrag und -qualität zu liefern." - npj Systems Biology and Applications [6]
Der erfolgreiche Einsatz ist nicht nur ein biologisches Problem. Er hängt von einer engen Kopplung zwischen Schaltungsentwicklung, Bioprozessdesign, regulatorischer Dokumentation und Beschaffung ab.
FAQs
Wie verbessern synthetische Genkreise die Konsistenz der Differenzierung?
Synthetische Genkreise können die Differenzierung konsistenter machen, weil sie Ihnen programmierten Kontrolle über das Zellverhalten und die Linienbindung geben.In der Praxis bedeutet das, modulare logische Operationen zu verwenden, um die Expression von Genen und Transkriptionsfaktoren mit präzisem Timing zu steuern.
Dieses Timing ist wichtig. Es hilft den Zellen, definierte Zustandsänderungen in der richtigen Reihenfolge zu durchlaufen, anstatt in gemischte oder unerwünschte Zustände abzudriften. Es reduziert auch die Differenzierung außerhalb des Ziels und verringert das Rauschen in der Kultur.
Der Nutzen ist klar: einheitlichere, stabilere und reifere Zellpopulationen für die Produktion von kultiviertem Fleisch.
Welches Schaltungsdesign eignet sich für myogene oder adipogene Kontrolle?
In der Forschung zu kultiviertem Fleisch können dieselben Hühnerfibroblasten in beide Linien gedrängt werden. Myogenese folgt einem Satz von Induktionsprotokollen, während Adipogenese durch das Aussetzen der Zellen an Eingaben wie Hühnerserum oder Fettsäuren aktiviert werden kann.
Von dort aus können diese Zellschicksale Schritt für Schritt in 3D-Hydrogelgerüsten kontrolliert werden, um Fleischstrukturen mit definierten Fett- und Kollagenverhältnissen aufzubauen.
Warum verhalten sich Genschaltkreise in 3D-Kulturen oft anders?
In 3D-Kulturen, verhalten sich Genschaltkreise oft anders, weil Zellen mit physischen und strukturellen Eingaben umgehen, die in 2D-Monolayern einfach nicht existieren. Diese Eingaben umfassen mechanische Spannung, Scherstress, Matrixsteifigkeit und lokale Zelldichte.
Diese Signale können Signalwege wie Notch verschieben. Sie können auch ändern, wie synthetische Schaltkreise Kraft erkennen und nachgelagerte Reaktionen koordinieren, einschließlich Zell-Zell-Adhäsion und Gewebemorphogenese.









