

血清を除去しても同じ野生型細胞株を維持する場合、培地の調整だけで老化、ドリフト、または付着損失を止めることは期待できません。 この記事では、培養肉における血清フリーの成功は通常、システムの両側に依存していることを示します:細胞外の定義された培地と、細胞が分裂を続け、付着し、筋原性機能を保持するのを助ける細胞内の編集です。
バイオプロセスエンジニアや細胞培養チームにとって、核心となるポイントは簡単です:
- 血清フリー培地は細胞の挙動を変えます, 成分リストだけではありません。血清フリー条件下では、グルコース、グルタミン、グリシン、シスチンの取り込みが変化する可能性があります。
- 一次衛星細胞は早期に細胞株の限界に達します。野生型のブタ細胞はしばしば継代10 . までに筋原性特性を失います。
- CDKN2Aノックアウトは、記事の中で最も明確な例の一つです:編集された豚の衛星細胞株は15回以上の継代 , で拡大され、 >90%の生存率, を維持し、いくつかのクローンでは野生型コントロールよりもPAX7が継代20で約194倍高いことが示されました。
- トレードオフがあります。 より良い拡大が後期継代の分化を保証するわけではありません。編集されたラインの中には、継代30. で依然として低い分化を示すものもあります。
- 検証は生産セットアップで行う必要があります: 同じ無血清培地、同じ培養モード、成長、廃棄物の蓄積、系統マーカー、融合のための同じ読み出し。
短いバージョン: 製造に移行できる無血清プロセスを望むなら、メディア設計、遺伝子編集、クローン選別、バイオリアクターフィットを4つの別々の作業ではなく、1つの連携したワークフローとして扱うべきです。
| フォーカスエリア | 最初に確認すること | 重要な理由 |
|---|---|---|
| 細胞周期制御 | CDKN2A, 継代寿命, PAX7 | 早期老化なしでラインが拡張できるかどうかを示すのに役立ちます |
| 成長シグナル伝達 | IGF1R、EGFR、FGFR応答 | 血清フリーシステムは外部シグナルサポートが低い |
| ストレス耐性 | 生存率、アポトーシスマーカー、せん断応答 | 血清除去と継代が細胞を喪失に追い込む可能性がある |
| 栄養処理 | グルコース使用、乳酸、アンモニア、アミノ酸取り込み | より速い取り込みは、より速い廃棄物の蓄積を意味することもある |
| アイデンティティ保持 | PAX7, MYOD, MYOG, 融合指数 | 急成長するラインは、ターゲット組織を形成しなくなった場合、役に立ちません |
その後、血清不使用の培養が失敗する箇所、失敗点に対応する編集、およびプロセス移行前に編集されたラインをどのように検証するかを説明します。
哺乳類細胞株におけるCRISPR-Casゲノム編集 | プロトコールプレビュー
無血清培養における主な生物学的障壁
血清を除去すると、シグナル伝達、付着、細胞のアイデンティティという3つのボトルネックが露呈します。問題は細胞内, で始まり、メディアの配合だけではありません。それが重要なのは、チームが時間を費やす場所、つまりメディアの調整、遺伝子編集、またはその両方の組み合わせを形作るからです。
| 特徴 | 血清補充培養 | 無血清培養 |
|---|---|---|
| 増殖 | 多様な成長因子によってサポートされる強力な増殖 | 変動性あり; 複製老化およびG1/S停止に陥りやすい |
| 付着 | 血清由来のECMタンパク質(フィブロネクチン、ビトロネクチン)によってサポートされる | 外因性のコーティングまたは添加物が必要; 分離リスクが増加 |
| 栄養輸送 | アルブミンやトランスフェリンなどのキャリアタンパク質によって促進される | 緩衝されていない取り込みに依存; 最適化されたITS-Xおよび脂質濃度が必要 |
| アポトーシスリスク | 低い; PI3K-AKTおよびMAPK-ERK経路が強く活性化される | 酸化ストレスと代謝廃棄物に対する感受性が高い |
| アイデンティティの安定性 | 初期から中期の継代で一般的に安定 | 表現型のドリフトのリスクが高い; 幹細胞マーカーは急速に低下することが多い |
成長と生存シグナルの喪失
血清が除去されると、成長因子のレベルが急激に低下する。細胞は、PI3K-AKTおよびMAPK-ERKの活動を高く保つための外部サポートの多くを失います。実際には、それはより多くのアポトーシスと弱い増殖を意味し、スケールアップに直接的な問題を引き起こします。
接着、栄養素の取り込み、およびストレスのボトルネック
血清は細胞に栄養を与えるだけではありません。接着と拡散をサポートするECMタンパク質も供給します。フィブロネクチン、ビトロネクチン、および関連因子がないと、一次衛星細胞は分離してアポトーシスに入る可能性が高くなります。特にバイオリアクター条件下での剪断応力下では。Y-27632によるROCK阻害はある程度役立ちますが、接着の問題を解決するわけではありません。
栄養素の取り扱いも難しくなります。血清キャリアタンパク質がないと、グルコース、グルタミン、グリシン、およびシスチンの取り込みが緩衝されにくくなります[1] . 同時に、アンモニアや乳酸のような代謝廃棄物が蓄積し、成長を抑制する可能性があります[3]. そのため、基礎培地が紙の上では問題ないように見えても、輸送と廃棄物のバランスが制限要因になることがあります。
血清不使用適応中の表現型ドリフト
血清不使用の適応は、新しい条件に耐えるサブポピュレーションを選択することがありますが、それが製品仕様に合わなくなることがあります。それが罠です:細胞はよく増殖するかもしれませんが、意図した組織を形成する能力を失う可能性があります。
連続継代を通じて、PAX7, MYOD, およびMYOGのようなマーカーが減少することがあります[2]. 適応中に系統マーカーを追跡し、長いメディア最適化サイクルの後ではなく、早期にドリフトを示すようにします。これらは、遺伝子編集が安定化する必要がある経路です。
血清不使用のパフォーマンスを向上させる遺伝子編集アプローチ
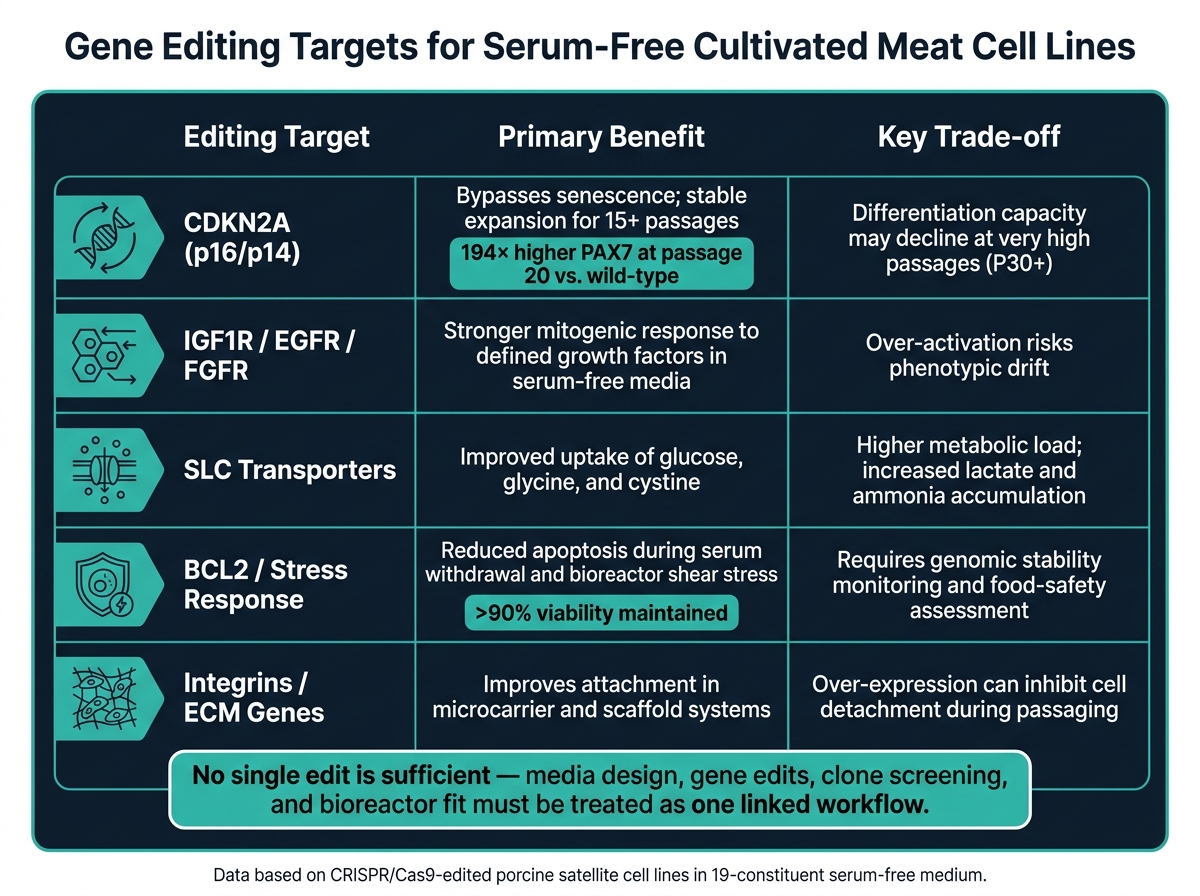
血清不使用の細胞培養における遺伝子編集ターゲット:利点とトレードオフ
これらの障壁は、シグナル伝達、生存、分化の3つの編集クラスに分類されます。
成長因子シグナル伝達と栄養素利用の編集
直接的なルートの一つは、IGF1R, EGFR, およびFGFRをアップレギュレートまたは感作して、細胞がIGF-1、EGF、およびbFGFにより強く反応するようにすることです[2]. これは、成長因子レベルが設計上通常低い血清不使用の培地で重要です。シグナル伝達が改善されると、細胞周期制御が次のボトルネックになることがよくあります。
ここで際立つのは、細胞周期調節因子 CDKN2Aです。CRISPR/Cas9 によるエクソン2のノックアウトは、19成分の無血清培地で15回以上の継代にわたって強く増殖する CDKN2A−/− 豚衛星細胞株を生成しました。特定のクローンでは、PAX7 の発現が野生型コントロールと比較して継代20で約194倍にアップレギュレーションされました [2].
溶質キャリア ( SLC) トランスポーターの編集は、グルコース、グルタミン、グリシン、シスチンのスケールアップ時の取り込み制限を回避するのに役立ちます [1]. しかし、問題があります。取り込みが増えると、乳酸とアンモニアの蓄積も速くなるため、トランスポーターの編集は、メディア交換と廃棄物管理と共に初日から計画する必要があります。取り込みの編集だけでは不十分です。
細胞の生存率と無血清ストレスへの耐性の向上
血清の撤退、ルーチンの継代、バイオリアクターのせん断はすべて、細胞を高アポトーシス条件に追い込みます。BCL2経路を編集すること - 生存促進メンバーをアップレギュレートするか、アポトーシス促進メンバーを抑制すること - は、これらの移行中の細胞損失を削減することができます。これは、細胞が付着ストレスと機械的ストレスの両方に対処するマイクロキャリアシステムでは、さらに重要になります。
生存を改善するか、増殖を延ばすための編集は、製造の全パッセージ範囲にわたるゲノム安定性チェックが必要です。CDKN2A−/−豚衛星細胞は、連続した無血清増殖中に90%以上の生存細胞率を維持しました [2]. それでも、チームは安定性が持続すると仮定するのではなく、設定されたパッセージ間隔で染色体の完全性を確認する必要があります。
接着、増殖、分化能力のバランス
最も難しい部分は、拡張と分化の間の引っ張りを管理することです。CDKN2Aノックアウトは、10回の継代を通じて筋原性の可能性を保持しますが、血清フリー条件下の野生型細胞は筋原性の特性をほぼ完全に失います。編集された系統では、融合指数が16.3%から56.3%と報告されています[2]. しかし、30回の継代までには、編集された細胞でさえ分化能力の低下を示すことがあります[2] .
| 編集対象 | 無血清培養における主な利点 | 主要なトレードオフ |
|---|---|---|
| CDKN2A (p16/p14) | 老化を回避し、15回以上の継代で安定した拡張 [2] | 非常に高い継代(P30+)では分化能力が低下する可能性あり [2] |
| IGF1R / EGFR / FGFR | 定義された成長因子に対する強い増殖反応 [2] | 過剰活性化は表現型のドリフトのリスクを伴う |
| SLCトランスポーター | グルコース、グリシン、シスチンの取り込みが改善 [1] | 代謝負荷が高く、乳酸とアンモニアの蓄積が増加 [1] |
| BCL2 / ストレス応答 | 離脱およびせん断ストレス時のアポトーシスの減少 [2] | ゲノム安定性の監視と食品安全性の評価が必要 [2] |
| インテグリン / ECM遺伝子 | マイクロキャリアおよびスキャフォールドシステムでの付着を改善 [2] | 過剰発現は継代中の細胞の剥離を抑制する可能性がある [2] |
接着の編集は、マイクロキャリアまたはスキャフォールドのセットアップで最も有用です。 それらは、すべての無血清プロセスの修正としてではなく、フォーマット固有のツールとして扱う方が良いです。
誘導性CRISPRシステムは、チームに拡張と分化のトレードオフを処理する実用的な方法を提供します。アイデアは簡単です:誘導性編集を使用して拡張フェーズを分化から分離します。
これらの編集のどれも、表現型が意図した無血清培地で保持されない場合には重要ではありません。
sbb-itb-ffee270
無血清培養のための編集された細胞株の構築と検証
適切な編集を見つけることは仕事の一部に過ぎません。より難しい部分は、その編集を無血清製造に対応できる安定した細胞株に変えることです。それには、編集、クローン選択、検証を一つのパイプラインで結びつける厳密なワークフローが必要です。そして、そのパイプラインは、すでに特定されたシグナル伝達、生存、付着の制約を直接テストする必要があります。
編集ツールと配信方法の選択
ターゲットがCDKN2Aの場合、細胞周期抑制因子を除去し、長期的な拡張をサポートすることを目的とする場合、CRISPR/Cas9ノックアウトは実用的な第一歩です[2]. 一次家畜細胞では、一般的な配信ルートにはLipofectamineのような非ウイルス性トランスフェクションシステムやlentiCRISPR v2のようなウイルスシステムが含まれます[2][4]. クローン作業に移る前に、配信効率を確認してください。
時には過小評価されがちな重要なポイントがあります:生産のために計画された正確な培地と培養モードで各クローンをスクリーニングすること. 製造プロセスが定義された無血清培地、静的接着培養、マイクロキャリアまたは他のセットアップを使用する場合、それがスクリーニング中に細胞が直面すべき条件です。
生産用無血清製剤における編集細胞のスクリーニング
一般的な方法は、希釈限界によってクローンを分離し、ターゲット遺伝子座での編集をサンガーシーケンシングで確認することです[2]. 編集が確認されたら、製造を目的とした同じ無血清製剤と培養モードでスクリーニングを続けるべきです[2][1].
この段階では、クローンが編集をただ生き延びるのではなく、プロセスと共存できるかどうかを示す基本を測定します:
- 成長
- 生存率
- グルコース消費
- 乳酸生成
- アンモニア蓄積
また、PAX7 RT-qPCRを早期に追加することも理にかなっています。なぜなら、幹細胞性の喪失は、より明白な形でラインが失敗する前に現れることがあるからです[1][2].
プロセス移行前の編集細胞の特性評価
プロセス移行前には、検証は4つの関連領域をカバーする必要があります: ゲノム編集、経路応答、継代安定性、機能。それぞれが異なる問題に答えます。ゲノムチェックは表現型ドリフトのリスクに対処します。使用済み培地分析は、栄養素の取り込みと廃棄物の蓄積限界を示します。融合指数は、筋原性分化がまだ存在するかどうかを示します[2][1].
| アッセイタイプ | 測定内容 | 血清不使用の培養肉ラインにおける重要性 |
|---|---|---|
| T7 エンドヌクレアーゼ I / サンガーシーケンシング | 編集効率と正確なゲノム配列 | スケーリング前に遺伝子ノックアウトまたはノックインの成功を確認[2] |
| RT-qPCR (PAX7, MYOD, MYOG, BAX, CCND1) | 幹細胞性、分化、アポトーシスマーカーの転写レベル | 長期継代にわたる細胞の健康状態と分化能力をモニター[2][4] |
| 免疫蛍光 (MyHC / CK18) | 系統特異的なタンパク質発現 | 編集と適応後も細胞が筋肉または上皮のアイデンティティを保持することを保証します [2][4] |
| 使用済み培地分析 | グルコース、アミノ酸、乳酸およびアンモニアのプロファイル | 栄養素の要件を決定し、バイオリアクターの供給戦略に情報を提供します [1] |
| 融合指数 | 多核筋管に組み込まれた核の割合 | 血清なしで筋原性分化能力が保持されていることを確認します [2] |
| テクスチャープロファイル分析 (TPA) | 3D構造物の硬さ、弾力性、噛みごたえ | 編集された細胞が肉のような物理的特性を持つ最終製品を生産することを検証します [2] |
ゲノム検証 は、T7エンドヌクレアーゼIアッセイと個々のクローンのサンガーシーケンシングに依存しています[2]. 経路確認は、RT-qPCRまたはウェスタンブロットを使用して、計画された転写物またはタンパク質の変化が実際に起こったことを示します。マーカーには、PAX7, MYOD, MYOGおよびMyHCが含まれます。[2] [4].
長期安定性 のために、基準は成長、活力、およびマーカー発現の繰り返しチェックを伴う15-30パッセージです。CDKN2A ノックアウト豚衛星細胞は、無血清条件で15パッセージ以上90%の生存細胞率を維持しましたが、分化能力は30パッセージで低下し始めました。[2].
機能テストは、次に最も単純な質問をします:これらはまだ必要な細胞ですか?筋原性系統では、融合指数は、編集された細胞が無血清で多核筋管を形成できるかどうかを示します。[2]. テクスチャープロファイル分析(TPA)は、3D構造が肉のような硬さ、弾力性、噛みごたえを示すかどうかを確認します [2].
これらのデータを使用して、無血清製造のためのクローンの移行条件を設定します。
編集された細胞株から無血清製造へ
編集された細胞を培地とバイオリアクターデザインにマッチング
クローンが検証を通過すると、仕事が変わります。その時点での成功は、細胞株がプロセスにどれだけ適合するかに依存します。さらなるスクリーニングでは、プロセスの不適合を修正できません。
使用済み培地の分析は、グルコースの追加、アミノ酸の補充、bFGFやIGF-1などの定義された入力を含む成長因子の投与を促進するべきです[2]. 接着系において、足場の播種密度と接着ウィンドウ - バイオリアクターへの移行の約2時間前 - は、血清含有プロトコルではなく、編集されたラインの付着挙動から設定する必要があります[2]. そのデータは、供給タイミング、播種密度、移行タイミングの決定に直接反映されるべきです。
編集されたラインは、より長い拡張、より高い細胞密度、より安定したマーカー発現をサポートできます。つまり、スケールアップは野生型の仮定ではなく、編集されたラインの測定された挙動に従う必要があります。
実際には、ライン選択は生物学的な決定だけでなく、調達とスケールアップの決定になります。
R&D、製造、調達チームへの重要なポイント
血清フリー適応は単なるメディア配合の問題ではありません。それは細胞株から始まり、メディアの最適化だけでは解決しません。標的遺伝子編集、特に細胞周期抑制因子のノックアウト、例えばCDKN2A, は、血清非依存条件で一次衛星細胞が機能しない原因となる基礎生物学に対処します。CDKN2A−/−豚衛星細胞は、継代20で野生型コントロールと比較して約194倍高いPAX7発現を維持し、継代10で融合指数が最大56.3%に達しました - 非編集細胞が筋原性機能をほぼ失った段階です[2].
開発と製造のチーム間での分担はかなり明確です:
- R&Dチーム は、実際の生産条件下で編集されたクローンを最初からテストする検証パイプラインを構築する必要があります。それには、成長、栄養消費、系統安定性、および3D分化能力が含まれます。
- 生産チームは、血清を含むプロトコルからコピーされた仮定が当てはまらない可能性があるため、編集されたラインの栄養プロファイルを使用して飼料設計とバイオリアクターパラメータを設定する必要があります。[1].
- 調達チームは、編集されたラインの特定の要件に一致する調達計画が必要です。これには、定義された成長因子、脂質、抗酸化物質、およびラインの接着プロファイルに適合する足場またはマイクロキャリアが含まれます。
よくある質問
なぜメディアの最適化だけでは不十分なのですか?
メディアの最適化だけでは不十分です。多くの場合、動物細胞は、大規模生産に必要な特性、例えばせん断応力抵抗性, 代謝効率, および 高密度懸濁液中での生存率. などを持っていません。
血清フリーメディアは重要ですが、それだけでは細胞の内在的な限界を解決できません。 それらの制限には、増殖寿命の制限, バイオリアクターストレスへの感受性, および種や発達段階による異なる栄養ニーズ.
が含まれます。血清不使用培養で最も重要な遺伝子編集はどれですか?
培養肉の生産において、最も重要な編集は、添加された成長因子への依存を削減するものです。一例として、CDKN2Aの削除があり、これは血清不使用条件下での豚衛星細胞の増殖と分化を改善することができます。別の方法として、FGF2と変異型RasG12V. の誘導性過剰発現のために筋肉幹細胞を設計することがあります。この設定は自己分泌シグナル伝達をサポートし、培地中の組換えFGF2の必要性を排除します。
製造のために編集された細胞株はどのように検証されるべきですか?
編集された細胞株は、製造性能と分化の可能性を確認するために、ゲノム、プロテオーム、および機能的なテストを受けるべきです。
実際には、それは編集が意図した通りに行われたことを確認し、 他の場所で問題を引き起こさないことを意味します。研究者は、遺伝子改変が標的組織への分化を妨げないこと、およびストレス耐性や血清非依存性成長などの意図した特性が期待通りに発現することを確認する必要があります。









