

培養肉の生産は、初代細胞系と不死化細胞系における細胞の成長の遅さと早期の老化によって制限されています. CRISPR遺伝子編集は、これらの課題を克服するためのターゲットを絞ったソリューションを提供します。
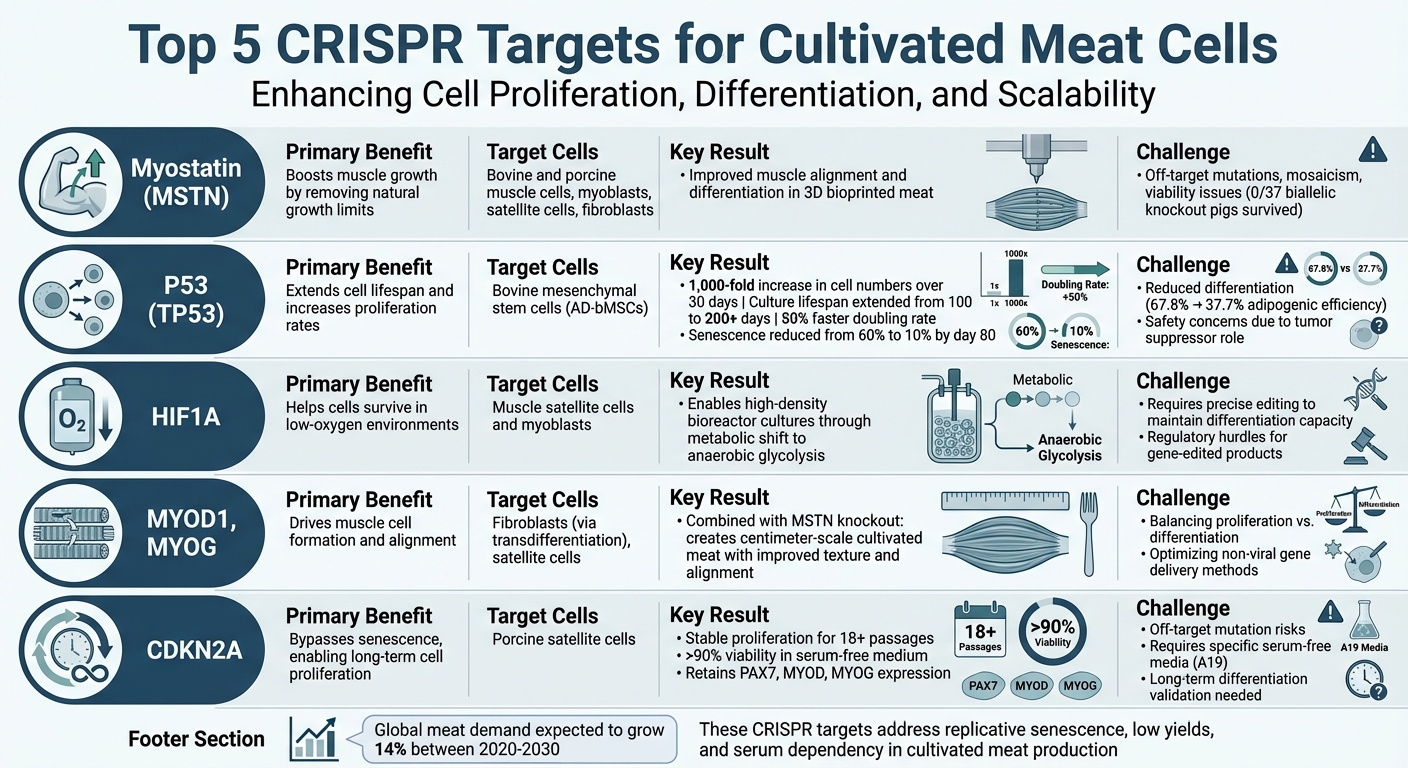
培養肉の細胞増殖、分化、スケーラビリティを改善するためのトップ5のCRISPRターゲットとその役割は次のとおりです:
- ミオスタチン (MSTN): 自然な成長制限を取り除くことで筋細胞の成長を促進します。
- P53 (TP53): 細胞の寿命を延ばし、増殖率を高めますが、分化を減少させる可能性があります。
- HIF1A: 低酸素環境での細胞の生存を助け、密集したバイオリアクター培養に不可欠です。
- 筋原性調節因子 (MYOD1, MYOG): 筋細胞の形成と整列を促進します。
- CDKN2A: 老化を回避し、長期的な細胞増殖を可能にします。
これらのターゲットは、複製老化、低収率、血清依存性といった重要な問題に対処します。しかし、増殖と分化のバランスを取り、安全性を確保することが成功の鍵となります。
htmlクイック比較:
| CRISPRターゲット | 主な利点 | 課題 |
|---|---|---|
| ミオスタチン (MSTN) | 筋肉成長を促進 | オフターゲット効果のリスク; 生存率の問題 |
| P53 (TP53) | 寿命を延ばし、増殖を促進 | 分化の減少; 安全性の懸念 |
| HIF1A | 低酸素環境での生存をサポート | 中断を避けるための正確な編集が必要 |
| MYOD1, MYOG | 筋形成を強化 | 増殖と分化のバランス |
| CDKN2A | 長期増殖を可能にする | オフターゲットのリスク; 無血清培地が必要 |
CRISPR技術は、培養肉の生産方法を再構築しており、より高い収量と低い生産コストを目指しながら、倫理的な懸念にも対処しています。
培養肉のためのCRISPRターゲットトップ5:利点と課題の比較
1. ミオスタチン遺伝子 (MSTN)
筋肉成長の自然なブレーキを解除することは、MSTN遺伝子をノックアウトすることで可能です。このプロセスは、過形成と肥大の両方を通じて筋肉細胞の増殖と分化を促進します [5][6].
主な利点
2025年3月、ソウル大学の研究者たちは、MSTN ノックアウト牛細胞とデジタルライトプロセッシング3Dバイオプリンティングを組み合わせることで大きな進展を遂げました。このアプローチは筋肉の配列と分化を改善し、従来のステーキに似た特性を持つ培養肉を生み出しました [5].
以前、2022年5月に、中国の西北農林科技大学の科学者たちは、最適化されたCRISPR/Cas9デリバリーシステム(100 ng/μLのCas9 mRNAと200 ng/μLのsgRNA)を使用して、ホモ接合型MSTNノックアウト羊を作成しました。生まれた16匹の子羊のうち、4匹がホモ接合型ノックアウトであることが確認されました。これらの子羊は、編集されていない対照群と比較して、30日、60日、90日で有意に高い体重を示し、pH、筋内脂肪、粗タンパク質レベルなどの肉質パラメータを維持しました[6] .
細胞タイプの適用性
MSTN遺伝子の編集は、一次筋芽細胞、衛星細胞、線維芽細胞(MYOD1駆動のトランス分化を通じて)、および間葉系幹細胞を含むさまざまな細胞タイプの筋原性の可能性を高めます。これは、細胞増殖の自然な限界を克服することによって達成されます [5][1].
潜在的な課題
その利点にもかかわらず、MSTNノックアウトには合併症がないわけではありません。生体動物における生存率の問題や、オフターゲット変異やモザイク現象などの技術的な障害が関連付けられています。例えば、2022年6月の研究では、MSTN編集された豚が筋肉成長の増加を示したものの、37匹の二対立遺伝子ノックアウト豚のうち生存したものはいなかったと報告されています[7] [8][6].
"MSTNノックアウトは、MYOD1を介したステーキタイプの培養肉の生産を促進します。" [5]
次に、P53腫瘍抑制遺伝子と持続的な細胞増殖を確保する上でのその重要性を探ります。
sbb-itb-ffee270
2. P53腫瘍抑制遺伝子
TP53遺伝子を無効にすると、重要な細胞周期チェックポイントが除去され、細胞増殖が大幅に加速されます。P53は腫瘍抑制因子として中心的な役割を果たし、細胞ストレスに応答して細胞周期の停止と老化を開始します。このチェックポイントがないと、細胞はバイオマスをはるかに速く蓄積し、より長い培養期間を維持することができます。[1].
主な利点
2025年初頭、Communications Biologyは、ウシ間葉系幹細胞におけるTP53編集の変革的効果を強調する研究を発表しました。結果は驚くべきものでした:30日間で細胞数が1,000倍に増加し、培養寿命が100日から200日以上に延長されました。編集された細胞は50%速い細胞倍加率を示し、80日目までに老化レベルは大幅に低下しました - 編集されていない細胞では約60%から、修正された細胞ではわずか10%に。さらに、これらの細胞は「若い」遺伝子発現プロファイルを保持し、DNA複製の強化と持続的なタンパク質合成が見られ、初期継代細胞を反映しています [1].
細胞タイプの適用性
ウシ脂肪由来間葉系幹細胞(AD‑bMSCs) は、TP53修飾に特に適しています。これらの細胞は自然に複製老化に直面し、増殖の可能性が制限されます。間葉系幹細胞は培養肉生産に使用される細胞源の約25%を占めるため、TP53編集は多能性を維持しつつ産業規模での拡張性をバランスよく提供する実用的な解決策です [1].
潜在的な課題
しかし、このアプローチには課題もあります。大きな欠点の一つは分化能力の低下です。The Communications Biology の研究では、脂肪分化効率が、非編集細胞の67.8%からTP53ノックアウトクローンでは37.7%に低下したと報告されています。トランスクリプトーム解析により、細胞周期遺伝子の活性が増加し、筋肉分化や接着に関連する遺伝子が減少していることが明らかになりました。さらに、TP53は重要な腫瘍抑制因子であり、その不活性化は癌の特徴であるため、この戦略は安全性と規制上の懸念を引き起こします。これらの細胞は医療用途ではなく消費を目的としているものの、そのような問題は慎重に考慮する必要があります。[1].
"すべての候補の中で、TP53ノックアウトが最も顕著な効果を示し、30日目までに1,000倍以上の増加を示しました。"
- Communications Biology [1]
次に、別の重要なCRISPRターゲットを探ってみましょう。
3.低酸素誘導因子1-アルファ (HIF1A)
HIF1Aは、統合センサーを備えたバイオリアクター. でよく遭遇する低酸素環境に適応するために培養肉細胞を助ける重要な役割を果たします。この調節因子は、酸素の浸透が制限されると特に重要になります。CRISPRを使用してHIF1Aを安定化させることで、細胞は酸素レベルが低下してもエネルギー生産を維持し、生存可能な状態を保つことができます。
主な利点
HIF1Aを編集することで、細胞の代謝が酸素依存の呼吸から嫌気性解糖にシフトします。このシフトにより、低酸素条件下でも細胞がエネルギーを生産し続けることが保証されます。その結果は?培養肉生産のスケールアップ, 特に厚い組織構造を作成する際に、酸素欠乏のリスクなしに細胞を高密度で培養する能力です。
細胞タイプの適用性
筋衛星細胞と筋芽細胞は、HIF1Aの編集から最も恩恵を受けます。これらは筋繊維の発達における重要なプレーヤーであり、密集したバイオリアクター内での生存は高収量を達成するために重要です。安定化されたHIF1Aは、これらの細胞が代謝経路を効果的に切り替えることを可能にし、長期間の培養中でも生存可能であることを保証します。
潜在的な課題
主な課題の一つは、編集された細胞が複数回の継代後も機能的な筋繊維に分化する能力を維持することです。これは、分化能力の喪失を避けるために技術的な微調整が必要です。研究室を超えて、規制上の障害や公衆の認識が複雑さを増します。遺伝子編集された肉製品は、市場に出る前に人間の消費と環境への影響に関する広範な安全性評価を通過しなければなりません。一方で、このような製品の消費者受け入れは地域によって大きく異なります[3]. これらの課題は、新しいターゲットに拡大する前に遺伝子編集技術を完璧にする必要性を浮き彫りにしています。次に、筋原性分化をさらに強化する遺伝子を探ります。
4. 筋原性調節因子 (MRFs: MYOD1, MYOG)
MYOD1は細胞を筋原性系譜にコミットさせる重要な役割を果たし、MYOGは筋芽細胞を成熟した筋管に融合させます。興味深いことに、MYOD1を過剰発現させると、線維芽細胞を筋原性細胞に再プログラムすることができ、一次衛星細胞で見られる自然老化の限界を効果的に回避します[5].
主な利点
MYOD1の過剰発現がウシ線維芽細胞でのMSTNノックアウトと組み合わされ、100‑µm溝パターンのハイドロゲル, 上でのDLP 3Dバイオプリンティングと統合されると、結果は印象的です。このアプローチは筋肉の整列と分化を強化し、センチメートル規模の培養肉構造の作成を可能にします。Journal of Animal Science and Biotechnologyに2025年3月に発表された研究では、MYOD1の非ウイルス送達とCRISPRを介したMSTNノックアウトを使用してウシ線維芽細胞を工学的に改変するこの方法が紹介されました[5]. 筋肉分化に対する抑制信号を排除することにより、この戦略は細胞をより強い筋原性アイデンティティに向かわせ、より良い食感の培養肉を生み出します。この二重アプローチは、増殖と分化経路の正確なバランスの重要性を強調しています。
細胞タイプの適用性
線維芽細胞はMYOD1ターゲティングのための優れた出発点です。 簡単な皮膚生検(通常の耳タグ付けに似ています)を通じて容易に得られるこれらの細胞は、中胚葉に由来し、MYOD1誘導による転分化に良好に反応します[5]. 一方、衛星細胞は、新生児の筋核の最大30%に寄与する能力を持ちながら、年齢とともに著しく減少します。これにより、線維芽細胞は産業規模の培養肉生産において、より実用的でスケーラブルな選択肢となります。
潜在的な課題
主な障害の一つは、細胞増殖と分化のバランスを見つけることです。例えば、TP53ノックアウトのような細胞拡張を促進するための遺伝子改変は、重要な筋分化因子を抑制し、細胞が機能的な筋組織に成熟する能力を妨げる可能性があります[1]. さらに、Piggybacトランスポゾンシステムのような非ウイルス法は食品安全の観点から好まれますが、効率的な遺伝子導入を確保するためには慎重な最適化が必要です。3Dプリントされたマイクログルーブのような外部要因は、適切な筋繊維の配列を達成するために重要です[5].
5. 細胞周期調節因子 ( e.g. , CDKN2A)
CDKN2Aは老化を引き起こす重要な役割を果たし、細胞分裂を効果的に停止させます。CRISPR/Cas9を使用してCDKN2Aをノックアウトすることで、研究者はヘイフリック限界を回避できます。これにより、筋肉幹細胞は通常の寿命をはるかに超えて分裂を続けることができ、機能的な筋肉組織に分化する能力を維持します。この画期的な技術は、培養肉生産における最大の課題の一つである、産業規模の製造に必要な大量の生存可能で機能的な細胞を生産するという問題に取り組んでいます。
主な利点
CDKN2Aを直接ターゲットにすることで、培養肉生産における細胞増殖の制限という問題に対処します。
CDKN2Aの編集は、スケーラビリティを向上させ、コストを削減します。例えば、2025年6月に、南京農業大学, のShijie Ding、Chunbao Li、Guanghong Zhouが率いる研究チームが、Food Materials Research. に彼らの研究結果を発表しました。彼らは、CDKN2Aノックアウトを持つCRISPR編集された豚の衛星細胞株を開発することに成功しました。これらの細胞は、A19無血清培地で18回以上の継代にわたって安定した増殖を示し、生存率は90%を超えました。重要なことに、これらの細胞は主要な筋原性調節因子(PAX7、MYOD、MYOG)の発現を保持し、成熟したMyHC陽性筋管に分化しました。植物ベースの3D足場に播種された場合、これらの編集された細胞は、噛みごたえと粘りが改善された肉のような構造を形成しました[2].
"CRISPRベースのCDKN2Aノックアウト細胞は、動物の生検に繰り返し依存することなく、筋肉前駆細胞の再生可能な供給源を提供します。" – 食品材料研究 [2]
細胞タイプの適用性
豚の衛星細胞, 筋肉再生に重要であり、CDKN2A編集に特に良く反応します。このアプローチは他の家畜種にも可能性を秘めています。CDKN2A編集細胞の主な利点は、無血清培地の処方. との互換性です。これにより、高価で倫理的に問題のある胎児ウシ血清の必要性が排除され、バッチ間の変動性が減少し、汚染リスクが最小限に抑えられます[2].
潜在的な課題
南京の研究は重要な利点を強調しましたが、培養肉におけるCRISPRの広範な応用には課題があります。オフターゲット変異は依然として懸念事項であり、慎重に監視する必要があります。さらに、遺伝子組み換え食品製品の規制安全基準を厳格に遵守する必要があります。研究者はまた、最終製品が自然の筋肉組織に近似することを保証するために、長期的な分化を確保する必要があります。これにより、プロトコルの改良と3Dスキャフォールドの徹底的な検証が不可欠となります[2].
これらの発見は、他のCRISPRターゲットとともに、以下の比較表にまとめられています。
比較表
表: 以下は、細胞増殖、分化、およびスケーラブルな培養肉生産のための代謝適応を改善する5つのCRISPRターゲットを要約したものです。
| CRISPRターゲット | 主な利点 | ターゲット細胞タイプ | 課題 | |
|---|---|---|---|---|
| ミオスタチン (MSTN) | 筋肉成長を促進 | ウシおよびブタの筋肉細胞 | 詳細なゲノム理解が必要; 注意深く管理しないと意図しない表現型変化のリスクあり[4] | |
| P53 (TP53) | 増殖を劇的に増加させる; 複製老化を遅らせる(30日目で細胞量が1,000倍以上増加)[1] | ウシ間葉系幹細胞 (bMSCs) | 分化能力の低下; 脂肪分化が67.8%から37に低下 | 7%; 筋肉関連遺伝子のダウンレギュレーション [1] |
| HIF1A | 代謝適応の改善 | ウシおよびブタの細胞 | 代謝の混乱を避けるために慎重な編集が必要 [4] | |
| MRFs (MYOD1, MYOG) | 筋繊維の形成と再生の鍵 | ブタの衛星細胞(筋肉幹細胞) [2] | 産業規模での急速な拡大中に高い発現レベルを維持するのは困難 [2] | |
| CDKN2A | 18回以上の継代で安定した増殖をサポートし、>90%の生存率を維持;老化を回避 [2] | ブタの衛星細胞(筋肉幹細胞) [2] | 特定の無血清培地 (e.g. 、A19)幹細胞性と分化を長期培養で維持するために[2] |
適切なターゲットを選択することは、細胞増殖と効果的に分化する能力のバランスを取ることを伴います。これは、培養肉細胞工学におけるこれらのプロセスを微調整することの重要性を強調しています。
結論
CRISPR技術は、限られた細胞増殖、老化、高い生産コストなど、培養肉生産における重要な課題に対処するための大きな可能性を秘めています。. 例えば、TP53ノックアウトは、わずか30日で細胞の豊富さを1,000倍以上に増加させることが示されています[1]. 同様に、CDKN2A編集は、無血清条件 で90%以上の生存率を持ちながら、15〜18パッセージにわたって安定的に細胞を増殖させることができます[2]. これは高価な動物血清への依存を減らし、動物の生検を繰り返す必要性を最小限に抑えます。
しかし、急速な細胞増殖と筋肉組織への分化能力のバランスを取ることは依然として重要な課題です。TP53ノックアウトは細胞数を大幅に増加させますが、分化を妨げる可能性があります。したがって、MYOD1 やMYOGのような調節因子の役割を維持することは、培養肉に適した成熟した筋肉組織を生成するために重要です。
これらの遺伝子戦略を適用しようとする研究チームにとって、
2020年から2030年の間に世界の肉需要が14%増加すると予想される中、[1], これらのCRISPRターゲットは、培養肉生産におけるスケーラブルでコスト効率の高いソリューションへの道を開きます。
よくある質問
分化に影響を与えずに成長を最も促進するCRISPRターゲットはどれですか?
成長を促進しながら分化を維持するための最良のCRISPRターゲットは、血清フリーの遺伝子工学的衛星細胞システム. です。この方法は、一貫した細胞成長と効果的な分化をサポートし、大規模な培養肉生産において強力な選択肢となります。
TP53またはCDKN2Aの編集を培養肉に安全に行うにはどうすればよいですか?
TP53またはCDKN2Aの編集を培養肉に安全に行うために、いくつかの重要なステップが取られます。これには、徹底的な遺伝的安定性試験, 、構造化された細胞バンキングシステムの確立, 、および次世代シーケンシングのような高度なツールを使用して突然変異を検出することが含まれます。さらに、厳格な規制遵守ガイドラインに従うことで、生産プロセス全体の安全性と一貫性が確保されます。
低酸素、高密度バイオリアクターで細胞が繁栄するための編集は何ですか?
適切な栄養素、成長因子、脂質、非必須アミノ酸、抗酸化物質を組み合わせた血清不使用の培地を開発することは、細胞の増殖と分化を促進する上で重要な役割を果たします。これらの調整は、細胞の生存率を向上させるだけでなく、特に低酸素や高密度環境のような困難な条件下での機能性も強化します。










