

การดัดแปลงฮิสโตนเป็นการเปลี่ยนแปลงทางเคมีของโปรตีนที่มีผลต่อกิจกรรมของยีนโดยไม่เปลี่ยนแปลง DNA การดัดแปลงเหล่านี้มีความสำคัญต่อการพัฒนาสายเซลล์ที่ใช้ในการผลิตเนื้อสัตว์เพาะเลี้ยง ช่วยให้เซลล์เติบโต รักษาเอกลักษณ์ และแยกแยะเป็นเนื้อเยื่อกล้ามเนื้อ บทความนี้สำรวจว่าการทำเครื่องหมายฮิสโตนเฉพาะ เช่น H3K4me3 (การกระตุ้นยีน), H3K27ac (กิจกรรมของเอนแฮนเซอร์) และ H3K27me3 (การกดขี่ยีน) ควบคุมพฤติกรรมของเซลล์อย่างไร
ประเด็นสำคัญที่ครอบคลุม:
- H3K4me3 สนับสนุนยีนที่ใช้งานและการแยกแยะอย่างรวดเร็ว
- H3K27ac ควบคุมเอนแฮนเซอร์สำหรับการแสดงออกของยีนในช่วงการเจริญเติบโต
- H3K27me3 ทำให้โปรแกรมยีนที่ไม่ต้องการยังคงไม่ทำงาน
- สถานะโครมาตินที่ถูกกำหนดโดยเครื่องหมายเหล่านี้แตกต่างกันไปในแต่ละสายพันธุ์และประเภทของเซลล์ ส่งผลต่อคุณภาพการผลิต
บทความยังเน้นการวิจัยล่าสุด รวมถึงการแสดงออกของยีนในตำแหน่งใน เซลล์สุกร ที่ส่งผลต่อคุณภาพเนื้อสัตว์และการแก้ไขอีพิเจเนติกส์ที่มีเป้าหมายสามารถปรับปรุงประสิทธิภาพของสายเซลล์ ทิศทางในอนาคตรวมถึงการปรับปรุงเครื่องมืออีพิเจเนติกส์และการศึกษาสถานะโครมาตินเพื่อเพิ่มประสิทธิภาพ การผลิตและขนาด.
การปรับเปลี่ยนฮิสโตนที่อธิบาย | การอะเซทิลเลชัน, การเมทิลเลชัน & การควบคุมยีน
sbb-itb-ffee270
ประเภทของการปรับเปลี่ยนฮิสโตนและหน้าที่ของพวกมัน
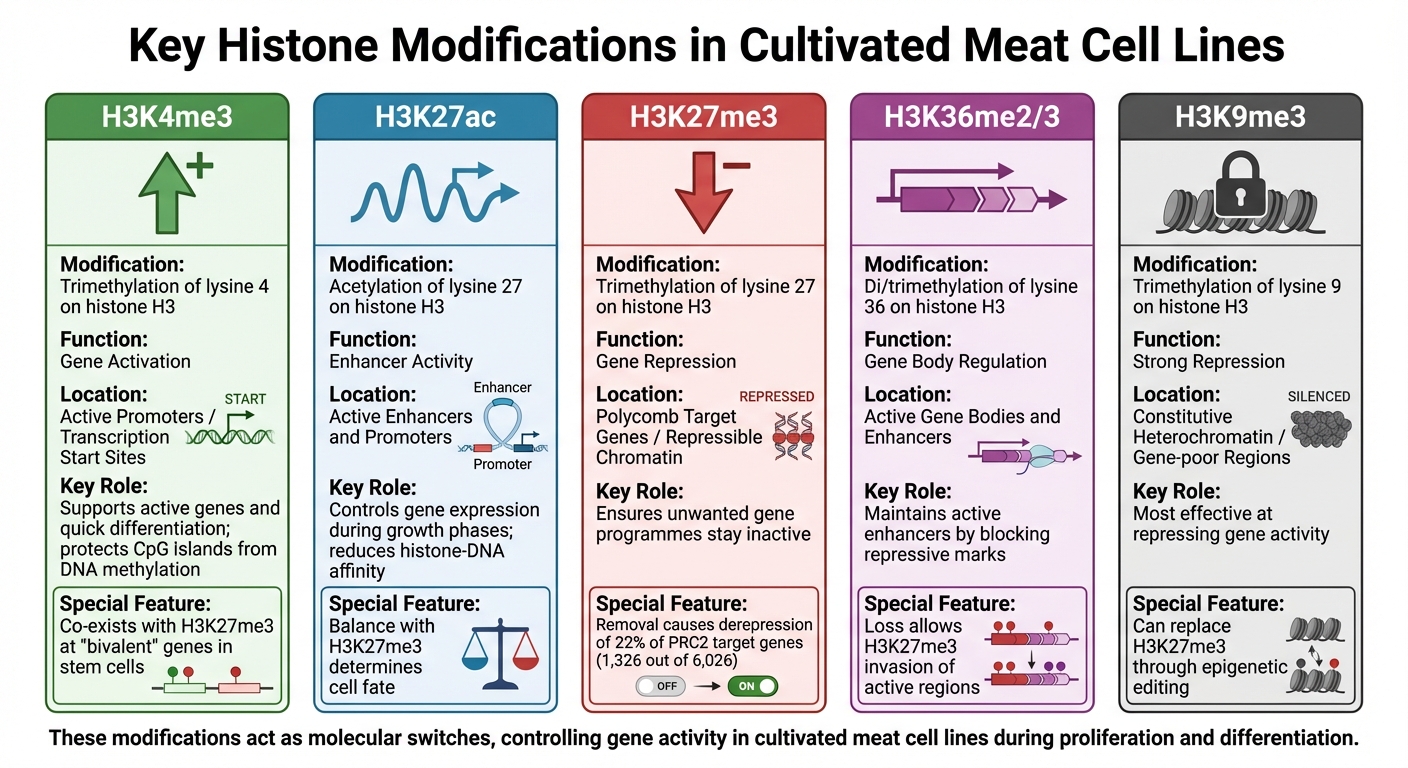
การปรับเปลี่ยนฮิสโตนที่สำคัญในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง: หน้าที่และบริบททางจีโนม
การปรับเปลี่ยนฮิสโตนมีบทบาทสำคัญในการควบคุมกิจกรรมของยีน ทำหน้าที่เหมือนสวิตช์โมเลกุลเพื่อควบคุมว่ายีนจะเปิดหรือปิดในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยงแท็กเคมีเหล่านี้ - ส่วนใหญ่คือเมทิลเลชันและอะเซทิลเลชัน - จะติดกับสารตกค้างเฉพาะบนฮิสโตน สร้างรูปแบบจีโนมที่แตกต่างกัน การปรับเปลี่ยนแต่ละครั้งมีหน้าที่เฉพาะ และโดยการทำความเข้าใจบทบาทเหล่านี้ นักวิจัยสามารถคาดการณ์และมีอิทธิพลต่อพฤติกรรมของเซลล์ได้ดีขึ้นในระหว่างการผลิต ความรู้นี้มีความสำคัญต่อการเพิ่มประสิทธิภาพกระบวนการใน การประมวลผลชีวภาพเนื้อสัตว์ที่เพาะเลี้ยง.
นี่คือการแยกย่อยของการปรับเปลี่ยนฮิสโตนหลักที่มีอิทธิพลต่อการควบคุมยีนในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง
H3K4me3 และการกระตุ้นยีน
H3K4me3 (การไตรเมทิลเลชันของไลซีน 4 บนฮิสโตน H3) เกี่ยวข้องกับโปรโมเตอร์ยีนที่ใช้งานและอำนวยความสะดวกในการถอดรหัสที่จุดเริ่มต้นของยีน โดยเฉพาะอย่างยิ่งสำหรับยีนที่เกี่ยวข้องกับการเจริญเติบโตและการเผาผลาญของเซลล์ การปรับเปลี่ยนนี้ยังปกป้องโปรโมเตอร์เกาะ CpG จากการเมทิลเลชันของ DNA ใหม่ ทำให้มั่นใจได้ว่ายีนที่จำเป็นยังคงสามารถเข้าถึงได้สำหรับการถอดรหัส [4].
ใน เซลล์ไลน์หลักหรือเซลล์ที่ทำให้เป็นอมตะ ที่ใช้สำหรับเนื้อสัตว์ที่เพาะเลี้ยง, H3K4me3 มักจะอยู่ร่วมกับเครื่องหมายที่กดทับเช่น H3K27me3 ที่ยีน "สองค่า" ยีนเหล่านี้ยังคงพร้อมสำหรับการกระตุ้น, ช่วยให้เกิดการแยกแยะอย่างรวดเร็วเป็นเนื้อเยื่อกล้ามเนื้อเมื่อจำเป็น [4].
น่าสนใจ, H3K4me3 มีปฏิสัมพันธ์กับการดัดแปลงอื่น ๆ ตัวอย่างเช่น, การสะสมของ H3K36me3 สามารถยับยั้ง H3K4 methyltransferases, ลดระดับ H3K4me3 ที่โปรโมเตอร์และเปลี่ยนแปลงรูปแบบการแสดงออกของยีน [4].
H3K27ac และกิจกรรมของ Enhancer
H3K27ac (การอะเซทิลเลชั่นของไลซีน 27 บนฮิสโตน H3) เป็นเครื่องหมายของ enhancer และโปรโมเตอร์ที่ใช้งาน โดยการลดความสัมพันธ์ระหว่างฮิสโตนและ DNA, H3K27ac สร้างสภาพแวดล้อมที่ส่งเสริมการถอดรหัส [5]. ในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง การเปลี่ยนแปลงระดับ H3K27ac ในช่วงการเจริญเติบโตต่างๆ กำหนดว่าเซลล์จะแสดงออกยีนใดเมื่อเคลื่อนจากการเพิ่มจำนวนไปสู่การแยกแยะ ความสมดุลระหว่าง H3K27ac และการดัดแปลงที่กดขี่เช่น H3K27me3 เป็นกุญแจสำคัญในการกำหนดชะตากรรมของเซลล์ ตัวอย่างเช่น การสูญเสีย H3K36me2 ซึ่งสนับสนุนกิจกรรมของ enhancer สามารถทำให้ H3K27me3 เข้าครอบครองพื้นที่ที่เคยใช้งานอยู่ ลดระดับ H3K27ac และปิดเสียงยีนเป้าหมาย H3K27me3 และการกดขี่ยีน H3K27me3 (การเติมเมทิลสามครั้งของไลซีน 27 บนฮิสโตน H3) เป็นเครื่องหมายกดขี่ที่ส่งเสริมโครงสร้างโครมาตินที่ปิด ทำให้ยีนถูกปิดเสียง การดัดแปลงนี้ซึ่งถูกเร่งโดย Polycomb Repressive Complex 2 (PRC2) มีความสำคัญต่อการรักษาการกดขี่ของยีนพัฒนาการนับพัน
ในสายเซลล์เนื้อที่เพาะเลี้ยง H3K27me3 ช่วยให้โปรแกรมยีนที่ไม่ต้องการยังคงไม่ทำงานในช่วงการเจริญเติบโตเฉพาะ เพื่อรักษาเอกลักษณ์ที่ตั้งใจไว้ของเซลล์
"H3K27me3 ร่วมกับ H2AK119ub1 มีความสำคัญในการรักษาการยับยั้งการถอดรหัสของยีนเป้าหมาย Polycomb หลายพันยีน" - Nature Communications [4]
การวิจัยแสดงให้เห็นว่าการลบ H3K27me3 ในเซลล์ต้นกำเนิดตัวอ่อนของหนูส่งผลให้เกิดการยกเลิกการยับยั้งของยีนเป้าหมาย PRC2 ประมาณ 22% (1,326 จาก 6,026) [4] . สำหรับเนื้อที่เพาะเลี้ยง การควบคุมการปรับเปลี่ยนนี้สามารถช่วยยับยั้งชะตากรรมของเซลล์ทางเลือก เช่น การสร้างไขมันหรือเนื้อเยื่อเกี่ยวพัน ในขณะที่มุ่งเน้นไปที่การพัฒนากล้ามเนื้อ
| การดัดแปลงฮิสโตน | หน้าที่การควบคุม | บริบททางจีโนม |
|---|---|---|
| H3K4me3 | การกระตุ้นยีน | โปรโมเตอร์ที่ใช้งาน / จุดเริ่มต้นการถอดรหัส |
| H3K27ac | กิจกรรมของเอนแฮนเซอร์ | เอนแฮนเซอร์และโปรโมเตอร์ที่ใช้งาน |
| H3K27me3 | การยับยั้งยีน | ยีนเป้าหมายของโพลีคอมบ์ / โครมาตินที่ยับยั้งได้ |
| H3K36me2/3 | การควบคุมภายในยีน | ภายในยีนและเอนแฮนเซอร์ที่ใช้งาน |
| H3K9me3 | การยับยั้งอย่างแรง | เฮเทอโรโครมาตินที่คงที่ / บริเวณที่มียีนน้อย |
สถานะโครมาตินในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง
การดัดแปลงฮิสโตนไม่ได้ทำงานเพียงลำพัง - พวกมันรวมกันเพื่อสร้างสถานะโครมาติน, ซึ่งเป็นสภาพแวดล้อมทางจีโนมที่ไม่ซ้ำกันที่ควบคุมการเข้าถึงยีนสถานะเหล่านี้มีบทบาทสำคัญในการกำหนดพฤติกรรมของสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยงทั้งในระหว่างการขยายตัวและการแยกแยะ ทำให้เป็นกุญแจสำคัญในการเพิ่มประสิทธิภาพกระบวนการชีวภาพ
การระบุสถานะโครมาตินผ่านเครื่องหมายฮิสโตน
นักวิจัยทำแผนที่สถานะโครมาตินโดยศึกษาการรวมกันของเครื่องหมายฮิสโตน เช่น H3K4me3, H3K27ac และ H3K27me3 ตัวอย่างเช่น ในเซลล์ไฟโบรบลาสต์ของตัวอ่อนสุกร (PFF) และเซลล์ทรอเฟคโตเดิร์ม (PTr2) มีการระบุสถานะโครมาตินที่แตกต่างกัน 10 สถานะ รวมถึงจุดเริ่มต้นการถอดรหัสที่ใช้งานอยู่ โปรโมเตอร์สองค่า และตัวเสริมที่คาดการณ์ไว้ [6]. สถานะเหล่านี้ช่วยทำนายกิจกรรมของยีน
สถานะตัวเสริมที่ถูกทำเครื่องหมายโดย H3K27ac ในบริเวณระหว่างยีนและอินทรอน มักจะมีการเพิ่มความเข้มข้นร่วมกับโปรตีนปรับโครงสร้างโครมาติน BRG1 [6].
คุณสมบัติที่โดดเด่นเป็นพิเศษคือการมีอยู่ของ โดเมน H3K4me3 กว้าง, ซึ่งครอบคลุมพื้นที่ 4 kb หรือมากกว่า โดเมนเหล่านี้คิดเป็นเพียง 1.7% ถึง 1.8% ของจุดเริ่มต้นการถอดรหัสที่คาดการณ์ทั้งหมดใน เซลล์ไลน์ของสุกร แต่มีความสำคัญในการทำเครื่องหมายยีนที่เฉพาะเจาะจงต่อการพัฒนาและเนื้อเยื่อ [6] . น่าสนใจว่า ในไฟโบรบลาสต์ของสุกรในครรภ์ 52% ของยีน ที่ถูกทำเครื่องหมายโดยโดเมนกว้างเหล่านี้เป็นยีนเฉพาะเนื้อเยื่อ เมื่อเทียบกับเพียง 25% ในเซลล์ PTr2 [6].
"การค้นพบเหล่านี้ช่วยเพิ่มความเข้าใจของเราเกี่ยวกับภูมิทัศน์ทางอีพิเจเนติกที่มีอยู่ในพัฒนาการของสุกรในระยะแรกและให้ข้อมูลเชิงลึกเกี่ยวกับวิธีที่ความแปรปรวนในสถานะโครมาตินเชื่อมโยงกับเอกลักษณ์ของเซลล์" - BMC Epigenetics & Chromatin [6]
โปรไฟล์สถานะโครมาตินเหล่านี้ไม่เพียงแต่แตกต่างกันภายในสายพันธุ์เดียว แต่ยังแตกต่างกันในสายเซลล์สัตว์ต่างๆ ที่ใช้ในการผลิตเนื้อสัตว์เพาะเลี้ยงอีกด้วย
ความแตกต่างของโครมาตินในสายเซลล์สัตว์
รูปแบบสถานะโครมาตินเปลี่ยนแปลงอย่างมีนัยสำคัญขึ้นอยู่กับสายพันธุ์และประเภทของเซลล์ที่ใช้ในการผลิตเนื้อสัตว์เพาะเลี้ยง ตัวอย่างเช่น ในสายเซลล์ไก่ H3K4me3 คิดเป็น 30% ถึง 55% ของการปรากฏตัวในจีโนมที่โปรโมเตอร์ของยีน [7]. อย่างไรก็ตาม ในเซลล์ต้นกำเนิดสืบพันธุ์ดั้งเดิมของไก่ (PGCs) ระดับ H3K4me3 ลดลงอย่างมากเมื่อเทียบกับเซลล์ที่มีความสามารถในการพัฒนาเป็นเซลล์หลายชนิด การลดลงนี้สนับสนุนการเปลี่ยนแปลงของสถานะสองขั้วไปสู่สถานะที่กดทับในระหว่างการกำหนดสเปคของสายพันธุ์สืบพันธุ์ [7].
เซลล์ Porcine trophectoderm (PTr2) แสดง ระดับ H3K27ac ที่สูงขึ้น ในบริเวณโปรโมเตอร์ (57.36%) เมื่อเทียบกับเซลล์ไฟโบรบลาสต์ของทารกในครรภ์ (41.58%) ในขณะที่การเพิ่มขึ้นของ H3K27me3 ต่ำกว่าในเซลล์ PTr2 (7.77%) เมื่อเทียบกับเซลล์ PFF (22%) [6]. ความแตกต่างเหล่านี้สะท้อนถึงความต้องการทางอีพิเจเนติกที่แตกต่างกันของแต่ละขั้นตอนการพัฒนาและมีผลต่อการตอบสนองของเซลล์เหล่านี้ต่อสภาพการเพาะเลี้ยง
ใน เซลล์ดาวเทียมของโค, การเปลี่ยนแปลงไปสู่ ชะตากรรม "เซลล์สำรอง" (Pax7+/Ki-67-) ถูกขับเคลื่อนโดยสถานะโครมาตินที่สงบซึ่งถูกควบคุมโดยสัญญาณ NOTCH และ MAPK/ERK อย่างไรก็ตาม กระบวนการนี้ลดผลผลิตโปรตีน [3]. ความแปรปรวนดังกล่าวเน้นย้ำว่าโครมาตินมีผลกระทบโดยตรงต่อประสิทธิภาพการผลิต การทำความเข้าใจความแตกต่างเหล่านี้อย่างลึกซึ้งเป็นสิ่งสำคัญสำหรับการปรับปรุงประสิทธิภาพของสายเซลล์ในการผลิตเนื้อสัตว์ที่เพาะเลี้ยง
การใช้การดัดแปลงฮิสโตนเพื่อปรับปรุงสายเซลล์
จากความรู้เกี่ยวกับสถานะโครมาติน เรามาสำรวจว่าการดัดแปลงฮิสโตนที่มีเป้าหมายสามารถปรับปรุงประสิทธิภาพของสายเซลล์เนื้อสัตว์เพาะเลี้ยงได้อย่างไร
การเพิ่มการแพร่กระจายและการปรับตัวให้เข้ากับการเจริญเติบโตแบบแขวนลอย
การปรับเครื่องหมายฮิสโตนสามารถเพิ่มการแพร่กระจายของเซลล์ได้อย่างมากและช่วยให้เซลล์เปลี่ยนจากการยึดติดไปสู่การเจริญเติบโตแบบแขวนลอย การเปลี่ยนแปลงนี้มีความสำคัญต่อระบบไบโอรีแอคเตอร์สำหรับเนื้อสัตว์เพาะเลี้ยง. ตัวอย่างเช่น การลดการเมทิลเลชัน H3K36 ทำให้ไฟโบรบลาสต์ตอบสนองต่อ TGFβ น้อยลง ส่งผลให้สถานะเซลล์มีความยืดหยุ่นมากขึ้น[1].
ในเดือนธันวาคม 2022 นักวิจัยที่ Believer Meats ประสบความสำเร็จในการพัฒนาครั้งสำคัญกับไฟโบรบลาสต์ไก่ (HUN-CF-2 และ HUN-CF-4)พวกเขาแสดงให้เห็นถึงการเป็นอมตะโดยธรรมชาติใน วัฒนธรรมแขวนลอยที่ปราศจากเซรั่ม, ถึง 100 ล้านเซลล์ต่อมิลลิลิตร (10⁸ เซลล์/มล.) และบรรลุผลผลิตที่ 36% w/v. ทีมงานที่นำโดย Yaakov Nahmias ใช้เลซิติน - โมเลกุลขนาดเล็กที่ปลอดภัยต่ออาหาร - เพื่อกระตุ้นเส้นทาง PPARγ และส่งเสริมการสร้างไขมันโดยไม่ต้องพึ่งพาการดัดแปลงพันธุกรรม ต้นแบบไก่ที่เพาะเลี้ยงของพวกเขาได้รับคะแนนประสาทสัมผัส 4.5 จาก 5.0 [2].
"การเป็นอมตะโดยไม่ต้องดัดแปลงพันธุกรรมและการผลิตที่ให้ผลผลิตสูงเป็นสิ่งสำคัญสำหรับการทำให้เนื้อเพาะเลี้ยงเป็นจริงในตลาด" - Yaakov Nahmias, หัวหน้าเจ้าหน้าที่วิทยาศาสตร์, Believer Meats [2]
การค้นพบเหล่านี้เน้นย้ำถึงศักยภาพของเครื่องมืออีพิเจเนติกที่แม่นยำในการปรับปรุงการพัฒนาสายเซลล์ให้ดียิ่งขึ้น
ความแม่นยำในการแก้ไขอีพิเจเนติก
เพื่อเสริมการเปลี่ยนแปลงของเซลล์เหล่านี้ วิธีการแก้ไขอีพิเจเนติกที่แม่นยำช่วยให้สามารถจัดการเครื่องหมายฮิสโตนได้อย่างตรงเป้าหมาย การศึกษาในปี 2025 เกี่ยวกับเซลล์ต้นกำเนิดจากตัวอ่อนของหนูแสดงให้เห็นว่าตัวเรียกใช้ไคเมอริก (S12N) ที่หลอมรวมกับโดเมนเร่งปฏิกิริยาจาก SUV39H2 หรือ SETD2 สามารถแทนที่ H3K27me3 ด้วย H3K9me3 หรือ H3K36me3 ที่ยีนหลายพันตัว ในบรรดานี้ H3K9me3 พิสูจน์แล้วว่ามีประสิทธิภาพมากกว่าในการยับยั้งกิจกรรมของยีน [8].
อย่างไรก็ตาม ความสำเร็จของการปรับเปลี่ยนเหล่านี้ขึ้นอยู่กับสภาพแวดล้อมของโครมาตินที่มีอยู่เป็นอย่างมาก ตัวอย่างเช่น H3K4me3 ที่เหลืออยู่ที่โปรโมเตอร์ของยีนสามารถขัดขวางเครื่องจักรเมทิลเลชันของ DNA ทำให้ยากต่อการบรรลุการปิดเสียงยีนที่ต้องการ [8]. สิ่งนี้บ่งชี้ว่าการเพิ่มประสิทธิภาพของเซลล์มักจะต้องปรับ เครื่องหมายฮิสโตนหลายตัวในเวลาเดียวกัน แทนที่จะมุ่งเน้นไปที่การปรับเปลี่ยนเพียงอย่างเดียว
บทสรุปและทิศทางในอนาคต
ข้อคิดสำคัญ
การปรับเปลี่ยนฮิสโตนมีบทบาทสำคัญในฐานะสวิตช์โมเลกุล, ที่ควบคุมกิจกรรมของยีนในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง โดยเฉพาะ H3K36me2 และ H3K36me3 ช่วยรักษาตัวกระตุ้นที่ใช้งานอยู่โดยการป้องกันไม่ให้เครื่องหมายกดทับเช่น H3K27me2/3 เข้ามาในร่างกายของยีน[9][10]. เมื่อการเมทิลเลชั่น H3K36 สูญเสียไป โครงสร้างโครมาตินจะถูกรบกวน ทำให้เครื่องหมายกดทับเช่น H3K9me3 เข้ามาในพื้นที่ที่ใช้งานอยู่[9]."การเมทิลเลชั่น H3K36 [เป็น] ตัวควบคุมที่สำคัญของสถานะโครมาตินและโครงสร้างจีโนม" - Nature Communications[9]
การโต้ตอบระหว่างเครื่องหมายฮิสโตนมีความสำคัญต่อการปรับปรุงประสิทธิภาพของสายเซลล์การวิจัยชี้ให้เห็นว่าการมุ่งเป้าไปที่การปรับเปลี่ยนฮิสโตนหลายอย่างพร้อมกันมักจะให้ผลลัพธ์ที่ดีกว่าการมุ่งเน้นเพียงอย่างเดียว [4].
ด้วยข้อค้นพบเหล่านี้ในใจ การศึกษาต่อไปต้องใช้เครื่องมืออีพิเจเนติกที่มีความแม่นยำเพื่อให้มั่นใจว่ามีการปรับปรุงอย่างต่อเนื่องในประสิทธิภาพของสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง
โอกาสในการวิจัยในอนาคต
การพัฒนาประสิทธิภาพของสายเซลล์ต้องการแนวทางที่เป็นนวัตกรรม เช่น การจัดลำดับ RNA ของนิวเคลียสเดี่ยว, เพื่อทำแผนที่ภูมิทัศน์อีพิเจเนติกภายในกลุ่มเซลล์ย่อยต่างๆ ซึ่งมีความสำคัญอย่างยิ่งในการระบุและทำความเข้าใจ "เซลล์สำรอง" ที่อยู่ในสภาวะสงบที่ต่อต้านการเปลี่ยนแปลง เซลล์เหล่านี้ซึ่งแสดงเครื่องหมายเช่น PAX7 และ NOTCH2 แทนที่จะมุ่งมั่นที่จะรวมตัวกับกล้ามเนื้อ ก่อให้เกิดความท้าทายที่สำคัญในการผลิตเนื้อสัตว์ที่เพาะเลี้ยง [3].
อีกแนวทางที่มีแนวโน้มดีคือการพัฒนาคอมเพล็กซ์เอพิเจเนติกแบบไคเมอริกสำหรับการควบคุมที่แม่นยำและไม่ใช่พันธุกรรม ตัวอย่างเช่น ในปี 2025 นักวิจัยได้แสดงให้เห็นว่าการรวม N-terminal ของ SUZ12 กับโดเมนที่มีฤทธิ์ทางเคมีจาก SUV39H2 หรือ SETD2 สามารถแทนที่ H3K27me3 ด้วย H3K9me3 หรือ H3K36me3 ที่ยีนจำนวนมากได้อย่างมีประสิทธิภาพ [4]. นอกจากนี้ การตรวจสอบ H3K36me2 ที่ตัวกระตุ้นสามารถทำหน้าที่เป็นเครื่องหมายควบคุมคุณภาพเพื่อให้แน่ใจว่าเส้นเซลล์มีความเสถียร [9].
ความพยายามในอนาคตควรมุ่งเน้นไปที่การรักษาการเมทิลเลชั่นของ H3K36 ข้ามรุ่นเซลล์ ซึ่งจะช่วยป้องกันการล่องลอยของเอพิเจเนติก ทำให้นักวิจัยและบริษัทต่างๆ เช่น
คำถามที่พบบ่อย
เครื่องหมายฮิสโตนมีผลต่อการแยกแยะกล้ามเนื้อในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยงอย่างไร?
เครื่องหมายฮิสโตนเป็นผู้เล่นหลักในการแยกแยะกล้ามเนื้อ โดยเฉพาะสำหรับสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง ตัวอย่างเช่น การลดลงของ H3K27me3 ในระหว่างการแยกแยะจะกระตุ้นโปรแกรมการถอดรหัสกล้ามเนื้อ ทำให้สามารถเปิดใช้งานยีนที่จำเป็นสำหรับการพัฒนากล้ามเนื้อ การปรับแต่งการดัดแปลงฮิสโตนอย่างละเอียด เช่น H3K27me3 สนับสนุนการเปลี่ยนแปลงของสายเซลล์จากการเพิ่มจำนวนไปสู่การสร้างเนื้อเยื่อกล้ามเนื้อที่มีลักษณะเฉพาะ การปรับเปลี่ยนทางพันธุกรรมเหล่านี้มีความสำคัญต่อการพัฒนาการผลิตเนื้อสัตว์ที่เพาะเลี้ยง
การดัดแปลงฮิสโตนใดที่ดีที่สุดในการทำนายการเจริญเติบโตของเซลล์ที่เสถียรและให้ผลผลิตสูงในเครื่องปฏิกรณ์ชีวภาพ?
การเมทิลเลชั่น H3K36 โดดเด่นในฐานะเครื่องหมายที่เชื่อถือได้สำหรับการเจริญเติบโตของเซลล์ที่เสถียรและให้ผลผลิตสูงในเครื่องปฏิกรณ์ชีวภาพ การปรับเปลี่ยนนี้มีบทบาทสำคัญในการรักษาเอกลักษณ์ของเซลล์และการจัดการโปรแกรมสายพันธุ์ - ซึ่งทั้งสองอย่างนี้มีความสำคัญต่อการรับรองการเพิ่มจำนวนเซลล์อย่างสม่ำเสมอ โดยเฉพาะในการผลิตเนื้อสัตว์ที่เพาะเลี้ยง
การแก้ไขอีพิเจเนติกสามารถปรับปรุงสายเซลล์โดยไม่เปลี่ยนแปลงลำดับดีเอ็นเอได้หรือไม่
การแก้ไขอีพิเจเนติกเสนอวิธีการปรับปรุงสายเซลล์โดยไม่เปลี่ยนแปลงลำดับดีเอ็นเอ โดยการปรับเครื่องหมายฮิสโตนและโครงสร้างโครมาติน มันควบคุมการแสดงออกของยีน การวิจัยเกี่ยวกับการปรับเปลี่ยนฮิสโตนเน้นให้เห็นว่าการเปลี่ยนแปลงเหล่านี้สามารถส่งผลต่อเอกลักษณ์และการทำงานของเซลล์ได้อย่างไร วิธีการนี้มีความหวังเป็นพิเศษในการปรับปรุงสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง










