

ヒストン修飾は、DNAを変化させることなく遺伝子の活動に影響を与えるタンパク質への化学的変化です。これらの修飾は、細胞が成長し、そのアイデンティティを維持し、筋肉組織に分化するのを助ける、培養肉生産に使用される細胞株の開発に不可欠です。この記事では、H3K4me3(遺伝子活性化)、H3K27ac(エンハンサー活性)、H3K27me3(遺伝子抑制)などの特定のヒストンマークがどのように細胞の挙動を調節するかを探ります。
カバーされた重要なポイント:
- H3K4me3 は活性遺伝子と迅速な分化をサポートします。
- H3K27ac は成長段階での遺伝子発現のためのエンハンサーを制御します。
- H3K27me3 は不要な遺伝子プログラムが非活性のままであることを保証します。
- これらのマークによって形成されるクロマチン状態は、種や細胞タイプによって異なり、生産品質に影響を与えます。
この記事では、ブタ細胞における位置的遺伝子発現が肉質に与える影響や、標的エピジェネティック編集が細胞株の性能を向上させる方法など、最近の研究を取り上げています。将来的な方向性としては、エピジェネティックツールの改良や、クロマチン状態の研究を通じて 生産効率と規模を最適化することが含まれます。.
ヒストン修飾の解説 | アセチル化、メチル化 & 遺伝子調節
sbb-itb-ffee270
ヒストン修飾の種類とその機能
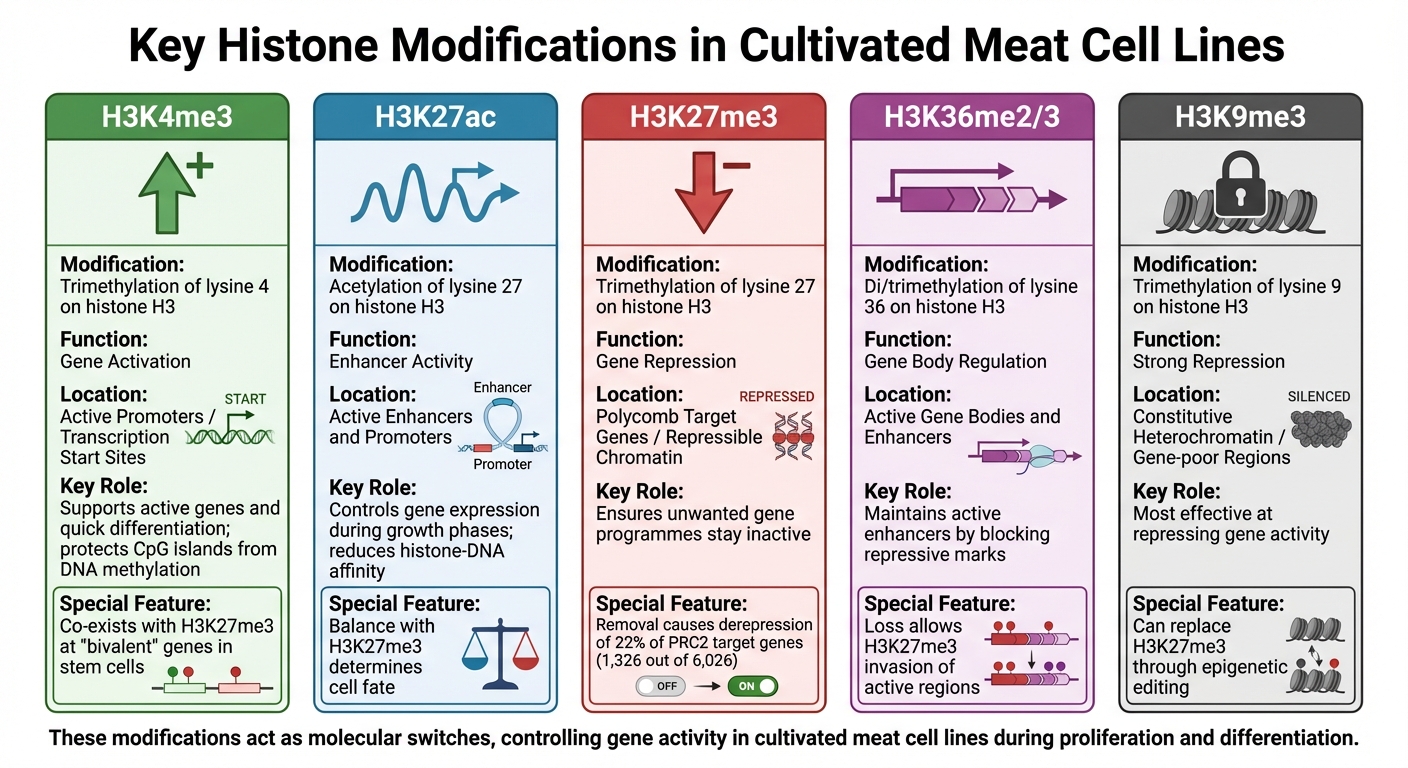
培養肉細胞株における主要なヒストン修飾:機能とゲノムコンテキスト
ヒストン修飾は、培養肉細胞株における遺伝子活動の調節において重要な役割を果たし、遺伝子がオンまたはオフになるかを制御する分子スイッチのように機能します。これらの化学タグ - 主にメチル化とアセチル化 - はヒストンの特定の残基に付着し、独自のゲノムパターンを作り出します。各修飾には特定の機能があり、これらの役割を理解することで、研究者は生産中の細胞の挙動をよりよく予測し、影響を与えることができます。この知識は、培養肉のバイオプロセス.
のプロセスを最適化するために不可欠です。培養肉の細胞株における遺伝子調節に影響を与える主なヒストン修飾の内訳を以下に示します。
H3K4me3と遺伝子活性化
H3K4me3(ヒストンH3のリジン4のトリメチル化)は、活性化された遺伝子プロモーターと関連しており、特に細胞成長と代謝に関与する遺伝子の開始部位での転写を促進します。この修飾はまた、CpGアイランドプロモーターを新たなDNAメチル化から保護し、重要な遺伝子が転写のためにアクセス可能な状態を維持します。 [4].
培養肉に使用される一次または不死化細胞株では、H3K4me3はしばしばH3K27me3のような抑制マークと共存し 、「二価」遺伝子で見られます。これらの遺伝子は活性化の準備が整っており、必要に応じて迅速に筋肉組織への分化を可能にします [4].
興味深いことに、H3K4me3は他の修飾と相互作用します。例えば、H3K36me3の付加はH3K4メチルトランスフェラーゼを抑制し、プロモーターでのH3K4me3レベルを低下させ、遺伝子発現パターンを変化させます[4].
H3K27acとエンハンサー活性
H3K27ac(ヒストンH3のリジン27のアセチル化)は、活性エンハンサーとプロモーターのマーカーです。ヒストンとDNAの親和性を低下させることで、H3K27acは転写を促進する環境を作り出します [5]. 培養肉の細胞株において、異なる成長段階でのH3K27acレベルの変化は、細胞が増殖から分化へ移行する際にどの遺伝子が発現されるかを決定します。
H3K27acとH3K27me3のような抑制的修飾のバランスは、細胞の運命を決定する鍵です。例えば、エンハンサー活性をサポートするH3K36me2の喪失は、H3K27me3が以前活性だった領域に侵入することを可能にし、H3K27acレベルを低下させ、標的遺伝子をサイレンシングします[5].
H3K27me3と遺伝子抑制
H3K27me3(ヒストンH3のリジン27のトリメチル化)は、遺伝子を効果的にサイレンシングする閉じたクロマチン構造を促進する抑制的マークです。この修飾は、Polycomb Repressive Complex 2 (PRC2)によって触媒され、数千の発生遺伝子の抑制を維持するために重要です [4].
培養肉の細胞株において、H3K27me3は特定の成長段階で不要な遺伝子プログラムが不活性のままであることを保証し、細胞の意図されたアイデンティティを保持します。
"H3K27me3はH2AK119ub1と共に、数千のポリコーム標的遺伝子の転写抑制を維持するために不可欠です。" - Nature Communications [4]
研究によれば、マウス胚性幹細胞でH3K27me3を除去すると、PRC2標的遺伝子の約22%(6,026のうち1,326)が脱抑制されることが示されています [4]. 培養肉において、この修飾を制御することで、脂肪や結合組織の形成などの代替細胞運命を抑制しながら、筋肉の発達に集中することができます。
| ヒストン修飾 | 調節機能 | ゲノムコンテキスト |
|---|---|---|
| H3K4me3 | 遺伝子活性化 | アクティブプロモーター / 転写開始部位 |
| H3K27ac | エンハンサー活性 | アクティブエンハンサーとプロモーター |
| H3K27me3 | 遺伝子抑制 | ポリコーム標的遺伝子 / 抑制可能なクロマチン |
| H3K36me2/3 | 遺伝子本体の調節 | アクティブな遺伝子本体とエンハンサー |
| H3K9me3 | 強い抑制 | 構成的ヘテロクロマチン / 遺伝子が少ない領域 |
培養肉細胞株におけるクロマチン状態
ヒストン修飾は単独で作用するのではなく、クロマチン状態, を形成するために組み合わさり、遺伝子のアクセス可能性を制御する独自のゲノム環境を作り出します。これらの状態は、拡張と分化の両方の過程で培養肉の細胞株の挙動を形成する上で重要な役割を果たし、バイオプロセシングの最適化において鍵となります。
ヒストンマークを通じたクロマチン状態の特定
研究者たちは、H3K4me3、H3K27ac、H3K27me3のようなヒストンマークの組み合わせを研究することでクロマチン状態をマッピングします。例えば、豚胎児線維芽細胞(PFF)および栄養外胚葉(PTr2)細胞では、10の異なるクロマチン状態が特定されており、活性転写開始部位、二価プロモーター、推定エンハンサーが含まれています[6] . これらの状態は遺伝子活性を予測するのに役立ちます。
エンハンサー状態は、主にH3K27acによって遺伝子間およびイントロン領域でマークされ、しばしばクロマチンリモデリングタンパク質BRG1と共に濃縮されています[6].
特に注目すべき特徴は、4 kb以上の領域にわたる広範なH3K4me3ドメイン, の存在です。これらのドメインは、豚の細胞系におけるすべての予測転写開始部位のうち、わずか1.7%から1.8%を占めていますが、発達および組織特異的な遺伝子を示すために重要です [6]. 興味深いことに、豚胎児線維芽細胞では、これらの広範なドメインによって示される遺伝子の 52%が組織特異的であるのに対し、PTr2細胞ではわずか25%です[6].
「これらの発見は、初期の豚の発達におけるエピジェネティックな風景の理解を深め、クロマチン状態の変動が細胞のアイデンティティとどのように関連しているかについての洞察を提供します。" - BMC Epigenetics & クロマチン [6]
これらのクロマチン状態プロファイルは、単一の種内で異なるだけでなく、培養肉生産に使用されるさまざまな動物細胞株間でも異なります。
動物細胞株間のクロマチンの違い
クロマチン状態パターンは、培養肉生産に使用される種や細胞タイプによって大きく変化します。例えば、鶏の細胞株では、H3K4me3は遺伝子プロモーターでのゲノム存在の30%から55%を占めています [7]. しかし、鶏の始原生殖細胞(PGCs)では、H3K4me3レベルは多能性細胞と比較して大幅に低下します。この減少は、生殖系列の特定中に二価状態が抑制状態に移行することをサポートします [7].
ブタ栄養膜外胚葉(PTr2)細胞は、胎児線維芽細胞(41.58%)と比較してプロモーター領域でのH3K27acレベルが高い(57.36%)一方、H3K27me3の濃縮はPTr2細胞(7.77%)で低く、PFF細胞(22%)よりも低いです[6]. これらの変動は、各発生段階の異なるエピジェネティックなニーズを反映しており、これらの細胞が培養条件にどのように反応するかに影響を与えます。
ウシ衛星細胞 , の「予備細胞」運命(Pax7+/Ki-67-)への分化は、NOTCHおよびMAPK/ERKシグナル伝達によって調節される静止クロマチン状態によって駆動されます。しかし、このプロセスはタンパク質収量を減少させます[3]. このような変動は、クロマチン状態が生産効率に直接影響を与えることを強調しています。これらの違いを深く理解することは、培養肉生産における細胞株の性能を微調整するために重要です。
ヒストン修飾を用いた細胞株の改善
クロマチン状態に関する既存の知識を基に、標的ヒストン修飾が培養肉の細胞株の性能をどのように直接改善できるかを探っていきましょう。
増殖の促進と懸濁培養への適応
ヒストンマークの調整は、細胞の増殖を大幅に増加させ、接着性から懸濁培養への移行を助けます。この変化は、培養肉のバイオリアクターシステムにとって重要です。. 例えば、H3K36メチル化の減少は、線維芽細胞をTGFβに対して反応しにくくし、より柔軟な細胞状態をもたらします。[1].
2022年12月、Believer Meatsの研究者たちは、鶏の線維芽細胞(HUN-CF-2およびHUN-CF-4)で画期的な成果を達成しました。彼らは、血清不使用の懸濁培養 , で自発的な不死化を示し、1mlあたり1億個の細胞(10⁸ cells/ml)に達し、36% w/v. の収率を達成しました。ヤアコブ・ナフミアス率いるチームは、遺伝子改変に頼らずに脂肪形成を促進するために、食品安全な小分子であるレシチンを使用してPPARγ経路を活性化しました。彼らの培養チキンのプロトタイプは、官能評価で5.0中4.5 [2].
「遺伝子改変なしの不死化と高収率の製造は、培養肉の市場実現にとって重要です。」 - ヤアコブ・ナフミアス、チーフサイエンティフィックオフィサー、Believer Meats [2]
これらの発見は、細胞株開発をさらに洗練するための正確なエピジェネティックツールの可能性を強調しています。
エピジェネティック編集の精度
これらの細胞変化を補完するために、正確なエピジェネティック編集方法はヒストンマークの標的操作を可能にします。2025年のマウス胚性幹細胞に関する研究では、SUV39H2またはSETD2の触媒ドメインと融合したキメラリクルーター(S12N)が、数千の遺伝子でH3K27me3をH3K9me3またはH3K36me3に置き換えることができることが示されました。これらの中で、H3K9me3は遺伝子活性の抑制においてより効果的であることが証明されました[8].
しかし、これらの修飾の成功は既存のクロマチン環境に大きく依存します。例えば、遺伝子プロモーターに残存するH3K4me3はDNAメチル化機構を妨げ、望ましい遺伝子サイレンシングを達成するのを難しくします[8]. これは、細胞の性能を最適化するには、単一の修飾に焦点を当てるのではなく、複数のヒストンマークを同時に調整する必要があることを示唆しています。
結論と今後の方向性
重要なポイント
ヒストン修飾は、培養肉の細胞株における遺伝子活性を制御する分子スイッチ, として重要な役割を果たします。特に、H3K36me2とH3K36me3は、H3K27me2/3のような抑制的なマークが遺伝子本体に侵入するのを防ぐことで、アクティブなエンハンサーを維持します [9][10]. H3K36メチル化が失われると、クロマチン構造が乱れ、H3K9me3のような抑制的なマークがアクティブな領域に侵入することを許します[9].
"H3K36メチル化は、クロマチン状態とゲノム構造の重要な調節因子である。" - Nature Communications[9]
ヒストンマーク間の相互作用は、細胞株の性能向上に不可欠です。研究によれば、複数のヒストン修飾を同時にターゲットにすることは、単一の修飾に焦点を当てるよりも良い結果をもたらすことが多いです。[4].
これらの発見を念頭に置き、将来の研究では、培養肉の細胞株の性能を継続的に向上させるために、精密なエピジェネティックツールを活用する必要があります。
将来の研究機会
細胞株の性能を向上させるには、シングルヌクレオチドRNAシーケンシング, などの革新的なアプローチが必要です。これは、異なる細胞サブポピュレーション内のエピジェネティックな風景をマッピングするために特に重要です。分化に抵抗する静止状態の「予備細胞」を特定し理解するために重要です。これらの細胞は、PAX7やNOTCH2のようなマーカーを発現し、筋原性融合にコミットする代わりに、培養肉の生産において重要な課題を提示します。[3].
もう一つの有望な方法は、正確で非遺伝的な制御のためのキメラエピジェネティック複合体の開発です。例えば、2025年に研究者たちは、SUZ12のN末端をSUV39H2またはSETD2の触媒ドメインと組み合わせることで、多くの遺伝子でH3K27me3をH3K9me3またはH3K36me3に効果的に置き換えることができることを示しました[4] . さらに、エンハンサーでのH3K36me2のモニタリングは、細胞株の安定性を確保するための品質管理マーカーとして機能する可能性があります[9] .
将来の取り組みは、細胞世代を超えてH3K36メチル化を維持することに焦点を当てるべきです。これにより、エピジェネティックドリフトを防ぎ、研究者や
よくある質問
ヒストンマークは培養肉の細胞株における筋分化にどのように影響しますか?
ヒストンマークは、特に培養肉の細胞株において筋分化の重要な役割を果たします。例えば、分化中のH3K27me3の減少は、筋形成に必要な遺伝子の活性化を可能にする筋原性転写プログラムを引き起こします。 H3K27me3のようなヒストン修飾の微調整は、細胞株が増殖から特定の特性を持つ筋組織を形成する移行をサポートします。これらのエピジェネティックな調整は、培養肉の生産を進めるために不可欠です。
バイオリアクターでの安定した高収量の細胞成長を最もよく予測するヒストン修飾はどれですか?
H3K36メチル化は、バイオリアクターでの安定した高収量の細胞成長の信頼できるマーカーとして際立っています。この修正は、細胞のアイデンティティを保持し、系統プログラムを管理する上で重要な役割を果たします。これらはどちらも、特に培養肉の生産において一貫した細胞増殖を確保するために不可欠です。
エピジェネティック編集は、DNA配列を変更せずに細胞株を改善できるのでしょうか?
エピジェネティック編集は、DNA配列を変更せずに細胞株を改善する方法を提供します。ヒストンマークとクロマチン構造を調整することで、遺伝子発現を制御します。ヒストン修飾に関する研究は、これらの変化が細胞のアイデンティティと機能にどのように影響するかを強調しています。このアプローチは、培養肉の細胞株を改良するための特に有望な方法です。










