

Ribosoomengineering hervormt de productie van gekweekt vlees door de eiwitsynthese op cellulair niveau te verbeteren. Ribosomen, de eiwitfabrieken van de cel, zijn cruciaal voor de productie van actine, myosine en andere eiwitten die de textuur en voedingswaarde van vlees bepalen. Standaard cellijnen zijn echter niet geoptimaliseerd voor de hoge productiviteit die nodig is voor grootschalige vleesproductie.
Belangrijke ontwikkelingen omvatten:
- Geoptimaliseerde ribosomaal RNA-varianten: Screeningsbibliotheken met 1,7 × 10⁷ varianten hebben potentieel getoond voor verhoogde translationele activiteit.
- Orthogonale ribosomen: Deze ontworpen ribosomen zijn gespecialiseerd in het produceren van specifieke eiwitten, zoals myosine, zonder de normale cel functies te verstoren.
- Codonoptimalisatie: Het aanpassen van mRNA-sequenties aan ribosoomvoorkeuren heeft tot 72-voudig hogere eiwitexpressie opgeleverd.
- Myokine signalering: Eiwitten zoals IL-15 en myonectine bevorderen ribosoom biogenese en eiwitsynthese tijdens spierdifferentiatie.
Uitdagingen blijven bestaan in het balanceren van energiebehoeften, het handhaven van celstabiliteit en het opschalen van productie naar industriële niveaus. Bijvoorbeeld, overactiviteit van ribosomen kan leiden tot verkeerd gevouwen eiwitten of metabole belasting, terwijl beperkingen in nutriëntendiffusie in bioreactoren de weefselgroei beperken tot 200 μm. Het aanpakken van deze problemen vereist de integratie van ribosoomengineering met geavanceerde bioprocessingstrategieën.
Dit artikel onderzoekt hoe deze methoden de toekomst van gekweekt vlees vormgeven en de hindernissen die overwonnen moeten worden om commerciële levensvatbaarheid te bereiken.
Ribosomen en Eiwitbiosynthese: Een Inleiding
Ribosoomstructuur en -functie in zoogdiercellen
Ribosomen staan centraal in de eiwitsynthese, waarbij mRNA-sequenties worden vertaald in functionele eiwitten.In zoogdiercellen worden ribosomen geclassificeerd als 80S-deeltjes, bestaande uit twee subeenheden: de 40S kleine subeenheid, die mRNA decodeert, en de 60S grote subeenheid, verantwoordelijk voor het katalyseren van peptidebindingvorming. Het vertaalproces omvat drie hoofd stappen: initiatie, waar de startcodon wordt herkend; elongatie, waar aminozuren opeenvolgend worden toegevoegd aan de groeiende polypeptideketen; en terminatie, die optreedt wanneer een stopcodon wordt bereikt.
Twee specifieke gebieden van de grote subeenheid zijn bijzonder belangrijk voor technische toepassingen: het peptidyltransferasecentrum (PTC), dat peptidebindingvorming vergemakkelijkt, en de uitgangstunnel, waardoor de nieuw gesynthetiseerde polypeptide uitkomt.[3].
Het begrijpen van deze kernmechanismen is essentieel voor het verkennen van hoe de prestaties van ribosomen geoptimaliseerd kunnen worden om de productie van gekweekt vlees te verbeteren.
Waarom Eiwitsynthese Belangrijk is voor Gekweekt Vlees
De efficiëntie van eiwitsynthese is een cruciale factor in de ontwikkeling van gekweekt vlees, met name tijdens in vitro myogenese. Dit proces transformeert spiersatellietcellen (MSCs) in multinucleaire myofibrillen die rijk zijn aan contractiele eiwitten zoals actine en myosine. Ribosomen spelen een centrale rol in deze transformatie [4].
"ongeveer acht biljoen spiercellen zijn nodig om 1 kg eiwit te produceren uit een traditionele bioreactor met een capaciteit van 5.000 L" [5]
Deze verbluffende eis benadrukt hoe zelfs kleine verbeteringen in ribosomale efficiëntie de productierendementen aanzienlijk kunnen verhogen, wat direct invloed heeft op de commerciële haalbaarheid van gekweekt vlees.
Naarmate cellen rijpen, ondergaat hun ribosomale activiteit een verschuiving.Tijdens de proliferatiefase geven MSC's prioriteit aan snelle deling. Echter, drie tot vijf dagen na differentiatie verschuift de focus naar het synthetiseren van volwassen isoformen van contractiele eiwitten en het mogelijk maken van de fusie van cellen tot myotubes [4]. Deze overgang wordt gereguleerd door specifieke signaalmoleculen, of myokines.
Bijvoorbeeld, Interleukin‑15 (IL‑15) bevordert de accumulatie van Myosin Heavy Chain (MyHC) eiwit terwijl het eiwitafbraak vermindert, en fungeert als een belangrijke anabole factor tijdens spierontwikkeling [4]. Evenzo ondersteunt Myonectin spiergroei door eiwitsynthese te verbeteren via de PI3K/Akt/mTOR signaalroute [4]. Begrijpen hoe deze signaalroutes ribosoomactiviteit beïnvloeden is essentieel voor het ontwerpen van schaalbare cellijnen die voldoen aan de productie-eisen. Deze inzichten vormen de basis voor de technische strategieën die in de volgende secties worden besproken.
Huidig Onderzoek naar Ribosoom Engineering
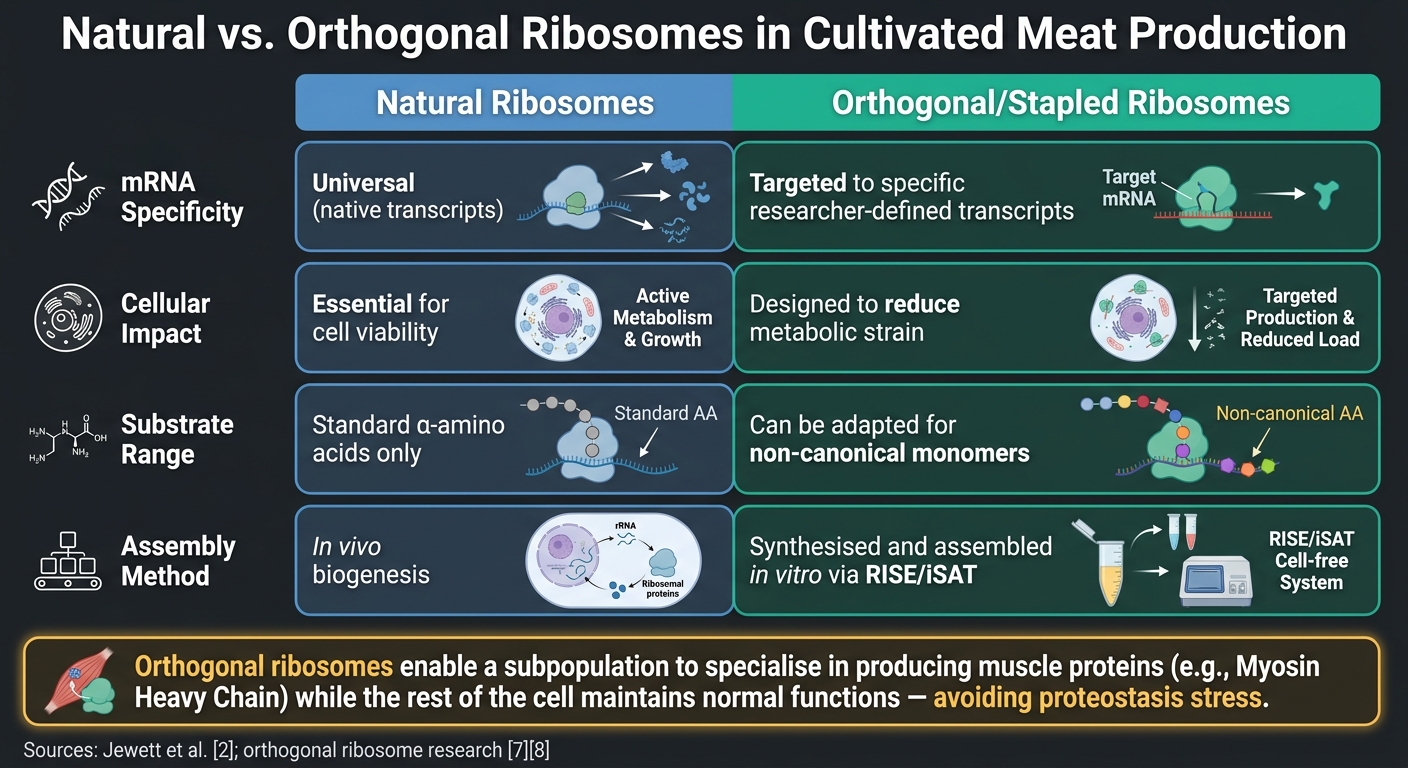
Natuurlijke vs. Orthogonale Ribosomen in de Productie van Gekweekt Vlees
Ribosoom Biogenese en Translatie Controle
Ribosoom biogenese, het proces waarmee cellen nieuwe ribosomen construeren, is een sterk gereguleerde en energie-intensieve activiteit. In zoogdiercellen vertegenwoordigt het een groot deel van de metabole output van de cel. Alleen al translatie kan tot wel 75% van het totale energieverbruik van een cel [8], verbruiken, waardoor het een van de meest hulpbron-intensieve cellulaire processen is.
Wanneer ribosoomtoewijzing inefficiënt is - bijvoorbeeld wanneer ribosomen vastlopen in vroege coderingsregio's - creëert het knelpunten die de beschikbaarheid van vrije ribosomen verminderen, wat uiteindelijk de eiwitproductie beperkt.Computational models hebben aangetoond dat het aanpakken van deze knelpunten door slechts 100 genen te manipuleren de ribosoomallocatie met 35% in gist (Saccharomyces cerevisiae) en 57% in Escherichia coli [8]. Deze bevindingen hebben directe implicaties voor het optimaliseren van ribosoomdynamiek in zoogdiercellen, met name in de gekweekte vleesindustrie, waar energie-efficiëntie en eiwitproductie cruciaal zijn.
Ribosoomengineering in de context van gekweekt vlees
Vooruitgang in ribosoomengineering wordt nu toegepast op de productie van gekweekt vlees, voortbouwend op de basiskennis van ribosoombiogenese. Zelfs onderzoek dat niet direct in spiercellen wordt uitgevoerd, levert inzichten op die relevant zijn voor gekweekte vleescellijnen.
In december 2020 toonden Hadas Zur en Tamir Tuller van Tel Aviv University het potentieel aan van Ribosome Traffic Engineering (RTE) om groeisnelheden en eiwitproductie te verbeteren. Met behulp van CRISPR-Cas9, introduceerden ze synonieme mutaties in de rampregio (codons 11–50) van RPO21 en CYS4 in S. cerevisiae. De resulterende dubbele mutant vertoonde verbeterde log-fase groei en celdichtheid. De onderzoekers waarschuwden echter dat de relatie tussen translatie-optimalisatie en groeisnelheid afneemt tijdens de diauxische shift en stationaire fasen, waar factoren buiten translatie beperkend worden [8]. Dit inzicht is bijzonder relevant voor het ontwerpen van differentiatieprotocollen in de productie van gekweekt vlees.
In februari 2020 valideerde het team van Michael Jewett aan Northwestern University de RISE (Ribosome In vitro Synthesis and Evolution) methode. Deze techniek omvat het screenen van een bibliotheek van ongeveer 1.7 × 10⁷ ribosomale RNA-varianten [2]. Door volledig buiten levende cellen te opereren, omzeilt RISE de beperkingen die worden opgelegd door dodelijke ribosoommutaties, die niet in vivo kunnen worden bestudeerd.
"De in vitro benadering overwint beperkingen van cellevensvatbaarheid, waardoor verkenning van dodelijke ribosoommutaties mogelijk wordt." - Michael Jewett et al. [2]
Een andere veelbelovende innovatie voor gekweekt vlees is het gebruik van orthogonale ribosomen. Deze ontworpen ribosoom-mRNA-paren functioneren onafhankelijk van het native translatieapparaat van de cel.Dit stelt onderzoekers in staat om ribosomale activiteit te richten op specifieke doelen, zoals Myosine Zware Ketting (MyHC) isovormen die cruciaal zijn voor de textuur van spieren, zonder essentiële cellulaire processen te verstoren [6]. Vergelijkende studies benadrukken de voordelen van orthogonale ribosomen ten opzichte van natuurlijke:
| Kenmerk | Natuurlijke Ribosomen | Orthogonale/Gestapelde Ribosomen |
|---|---|---|
| mRNA Specificiteit | Universeel (natuurlijke transcripties) | Gemaakt voor specifieke door onderzoekers gedefinieerde transcripties [6] |
| Cellulaire Impact | Essentieel voor levensvatbaarheid | Ontworpen om de metabole belasting te verminderen [7] |
| Substraatbereik | Standaard α-aminozuren | Kan worden aangepast voor niet-canonieke monomeren [7] |
| Assemblage | In vivo biogenese | Gesynthetiseerd en geassembleerd in vitro via RISE/iSAT [2] |
Het belangrijkste punt hier is dat orthogonale ribosomen een subpopulatie van ribosomen in staat stellen zich te specialiseren in het produceren van spierproteïnen, zoals MyHC, terwijl de rest van de cel normale functies behoudt.Dit voorkomt het risico van proteostase stress, die kan ontstaan wanneer het gehele translatie systeem wordt gedwongen om specifieke eiwitten overmatig te produceren.
Strategieën voor het Verbeteren van Ribosoomprestaties
Verhoging van Ribosoom Biogenese
Het verhogen van het aantal ribosomen is een directe manier om de eiwitproductie te verbeteren, en twee hoofdmethoden hebben de aandacht getrokken. De eerste betreft het wijzigen van de epigenetische toestand van ribosomaal RNA (rRNA) genen om hun translatiecapaciteit te vergroten.
"Epigenetische engineering van ribosomale RNA-genen verbetert de eiwitproductie." - Santoro R., Lienemann P., Fussenegger M. [1]
De tweede benadering maakt gebruik van de PI3K/Akt/mTOR signaalroute. Myokines zoals IL-15, myonectine en irisine activeren deze route, wat ribosoom biogenese stimuleert tijdens de rijping van myotubes, zoals eerder besproken.
Echter, deze toename in ribosoomproductie moet zorgvuldig worden uitgebalanceerd met de metabolische capaciteit van de cel, aangezien ribosoomsynthese een van de meest energie-intensieve processen in levende cellen is [1].
Zodra het aantal ribosomen is toegenomen, verschuift de focus naar het ervoor zorgen dat ze volledig betrokken zijn bij translatie.
Verbetering van Translatie Initiatie en Elongatie
Het maximaliseren van de activiteit van alle ribosomen is essentieel, aangezien zelfs in groei-geoptimaliseerde cellen 15–20% van de ribosomen inactief blijft [9]. Dit vertegenwoordigt een aanzienlijke reserve van onbenutte capaciteit in gekweekte vleescellijnen.
De snelheid van translatie elongatie hangt af van twee factoren: de inherente snelheid van het ribosoom en het aandeel ribosomen dat actief betrokken is bij translatie [9]. Om deze te optimaliseren, is het cruciaal om hoge aminozuurspiegels in het kweekmedium te handhaven.Bovendien helpt het manipuleren van cellijnen om ribosomale eiwitten te stabiliseren bij het beschermen van rRNA tegen misvouwing en degradatie, waardoor het typische verlies van 10% rRNA tijdens piekgroeicondities wordt verminderd [9].
Zodra de ribosoomactiviteit is gemaximaliseerd, wordt het verfijnen van mRNA-sequenties de volgende stap om de eiwitsynthese verder te versnellen.
mRNA-optimalisatie en codongebruik
De prestaties van ribosomen zijn sterk afhankelijk van de kwaliteit van het mRNA dat ze verwerken. Codonoptimalisatie stemt de coderingssequenties van doeleiwitten af op de tRNA-pool die specifiek is voor de gastheersoort - zoals rund, varken of vis. Deze afstemming voorkomt ribosoomstagnatie tijdens de elongatie en verhoogt de doorvoer voor kritieke myogene eiwitten zoals MyoD en Myf5.
Naast codonoptimalisatie zorgt transcriptionele afstemming voor een juiste balans tussen rRNA- en mRNA-niveaus binnen de cel.Elke mismatch tussen deze componenten kan knelpunten creëren, waardoor de algehele efficiëntie wordt verminderd [1].
Voor praktische toepassing bieden Geïntegreerde Synthese-, Assemblage- en Translatiesystemen (iSAT) een waardevol hulpmiddel. Deze systemen gebruiken celvrije extracten en op fluorescentie gebaseerde assays om geoptimaliseerde mRNA's in vitro te prototypen voordat ze worden geïntegreerd in stabiele cellijnen. Deze iteratieve benadering stelt onderzoekers in staat om snel codon-geoptimaliseerde varianten te vergelijken, waardoor de opbrengst van essentiële myogene eiwitten wordt verbeterd en de schaalbaarheid van de productie van gekweekt vlees wordt versterkt [1].
Afwegingen: Groei, Differentiatie en Productkwaliteit
Het optimaliseren van ribosoomprestaties vereist een delicate balans tussen het stimuleren van eiwitsynthese en het beheersen van de impact op celgroei en differentiatie, zoals eerder uiteengezet.
Metabole Last en Proteostase Stress
Het manipuleren van ribosomen om de eiwitproductie te verbeteren gaat gepaard met een verhoogde energievraag, omdat het ATP en aminozuren afleidt van andere essentiële cellulaire functies. De synthese van ribosomen is al een van de meest energie-intensieve processen binnen een cel, en verdere versterking kan deze energie-uitdagingen verergeren.
Deze verhoogde activiteit kan ook de eiwitkwaliteit beïnvloeden. Overactieve ribosomen kunnen cellulaire chaperonnes overweldigen, resulterend in verkeerd gevouwen eiwitten en activering van de unfolded protein response (UPR). Dergelijke stress kan de groei remmen of zelfs leiden tot celdood. Voor primaire volwassen stamcellen van vee zoals runderen of schapen, die van nature een beperkte proliferatiecapaciteit hebben, kunnen deze extra stressfactoren het aantal levensvatbare celdelingen aanzienlijk verminderen voordat senescentie optreedt [5].
Bij de productie van gekweekt vlees overschrijdt de weefseldikte zelden 200 μm vanwege beperkingen in de diffusie van voedingsstoffen, wat kan leiden tot celdood in de kern van grotere weefselaggregaten [5]. Strategieën die het energieverbruik verhogen, lopen het risico de uitputting van voedingsstoffen in deze kritieke gebieden te versnellen, waar consistente eiwitsynthese essentieel is. Bovendien kan verhoogde metabole belasting de fijn afgestemde signaalroutes verstoren die nodig zijn voor spierdifferentiatie.
Effecten op Spierdifferentiatie en Eiwitsamenstelling
De spanningen die worden geïntroduceerd door ribosoomengineering kunnen verder reiken dan de stofwisseling en mogelijk de spierontwikkeling verstoren.Myogenese, het proces van spiervorming, berust op een strak gereguleerde reeks transcriptiefactoren: Pax7 zorgt ervoor dat stamcellen in rust blijven, Myf5 bevordert de proliferatie van myoblasten, en MyoD triggert differentiatie [5] . Het veranderen van eiwitsynthese zou deze reeks kunnen verstoren, waardoor differentiatie wordt vertraagd of atypische spiervezelcomposities worden geproduceerd. Dit kan resulteren in minder intramusculaire vetafzettingen, die essentieel zijn voor het bereiken van de gewenste textuur en smaak in gekweekt vlees [5].
Als gevolg hiervan is het handhaven van rigoureuze kwaliteitscontrole door het monitoren van de expressie van myogene markers gedurende het hele engineeringproces essentieel om een goede spierontwikkeling en productkwaliteit te waarborgen.
sbb-itb-ffee270
Onderzoekshiaten en Toekomstige Richtingen
Vooruitgang in ribosoomengineering toont potentieel, maar de toepassing ervan op commerciële productie van gekweekt vlees kent nog aanzienlijke obstakels. Om deze hiaten te overbruggen, moeten onderzoekers zich richten op geavanceerde moleculaire profileringstechnieken en schaalbare bioprocesstrategieën die bestand zijn tegen de eisen van langdurige productie.
Multi-Omics en Langetermijn Stabiliteitsstudies
Een grote uitdaging ligt in het gebrek aan langetermijn stabiliteitsgegevens voor geëngineerde cellijnen. Na verloop van tijd kunnen deze cellen spontane mutaties ophopen, wat mogelijk hun fenotype verandert. Ivana Pajčin van de Universiteit van Novi Sad benadrukt deze zorg: geïmmortaliseerde cellen "zijn niet altijd representatief voor de primaire cultuur vanwege mogelijke spontane mutaties tijdens langdurige cultivatie" [13]. Voor ribosoom-gemanipuleerde lijnen zijn de inzet nog hoger - mutaties in ribosomale componenten kunnen de translatie-efficiëntie ondermijnen zonder onmiddellijke detectie.
Multi-omics benaderingen bieden een manier om deze problemen aan te pakken. Door transcriptomics, proteomics en metabolomics te integreren, kunnen onderzoekers kritieke myogene markers monitoren zoals Pax7, MyoD, en Myogenin, evenals verschuivingen in MyHC isoformen. Genoom-schaal metabole modellen kunnen deze inzichten vervolgens vertalen naar bruikbare veranderingen in de samenstelling van het medium om te voldoen aan de unieke eisen van gemanipuleerde ribosomen [5][11]. Voor gekweekt vlees is het essentieel om consistente eiwitproductie over langere cycli te waarborgen. Zonder dergelijke longitudinale monitoring is het moeilijk om duurzame verbeteringen te scheiden van kortstondige effecten.
Naast genetische en metabole stabiliteit, brengt het opschalen van deze innovaties naar industriële niveaus zijn eigen uitdagingen met zich mee.
Integratie en Opschaling van Bioprocessen
Het opschalen van ribosoom-gemodificeerde cellen van kleine flessen naar industriële bioreactoren is geen geringe prestatie. Het produceren van slechts 1 kg eiwit in een 5.000 L roertankbioreactor vereist ongeveer acht biljoen spiercellen [5]. Bij deze dichtheden worden nutriëntengradiënten een cruciaal probleem. De 200 μm diffusielimiet voor zuurstof en andere voedingsstoffen betekent dat cellen in de kern van 3D-weefselstructuren mogelijk honger lijden, vooral wanneer hun vraag naar middelen op zijn hoogtepunt is vanwege hoge eiwitsynthese.
Schuifspanning door bioreactoragitatie voegt een extra laag complexiteit toe. Terwijl ongemodificeerde cellen deze turbulentie kunnen verdragen, kunnen gemodificeerde cellen met aangepast translatiemachinerie kwetsbaarder zijn.De stress kan niet alleen cellulaire routes verstoren, maar ook fysiek schade toebrengen aan cellen die al onder metabolische druk staan [13]. Het aanpakken van deze problemen vereist het integreren van real-time data met digitale biomanufacturingmodellen, inclusief simulaties van computationele vloeistofdynamica, om de diverse micro-omgevingen binnen grootschalige vaten beter te begrijpen en te voorspellen [10]. Ook downstreamprocessen zoals oogsten hebben aandacht nodig - enzymatische methoden met trypsine kunnen het oppervlakteproteoom van ontworpen cellen veranderen [14], mogelijk de voordelen van ribosoomengineering tenietdoen.
| Schaalvergrotingsfactor | Belangrijkste knelpunt | Relevantie voor ribosoomengineering |
|---|---|---|
| Nutriëntendiffusie | 200 μm penetratielimiet [5] | Kan cellen met hoge eiwitsynthese-eisen in 3D-weefsels uithongeren |
| Genetische stabiliteit | Spontane mutaties [13] | Zou de efficiëntie van de geoptimaliseerde translatie na verloop van tijd kunnen aantasten |
| Schuifspanning | Roertank-turbulentie [13] | Risico's op verstoring van geoptimaliseerde cellulaire routes |
| Oogstmethode | Proteolytische schade door trypsine [14] | Kan het proteoom veranderen en verbeteringen in eiwitkwaliteit maskeren |
Het oplossen van deze opschalingsuitdagingen is essentieel voor het vertalen van ribosoomengineering van het laboratorium naar commerciële productie.Elke strategie moet grondig worden getest om betrouwbare eiwitopbrengsten, stabiliteit en veiligheid onder industriële omstandigheden te garanderen.
Conclusie: Het Pleidooi voor Ribosoomengineering in Gekweekt Vlees
Het produceren van 1 kg eiwit in een 5.000 L bioreactor vereist een verbazingwekkende 8 biljoen spiercellen [5]. Dit benadrukt de enorme uitdaging van het opschalen van de productie van gekweekt vlees. Ribosoomengineering biedt een oplossing door de eiwitproductie van individuele cellen te verbeteren, in plaats van simpelweg het aantal cellen te verhogen.
Timing is cruciaal bij het toepassen van ribosoomengineering. Het verbeteren van de translatie op het verkeerde moment kan de myogenese verstoren, wat mogelijk de productie van belangrijke contractiele eiwitten zoals MyHC beïnvloedt [5]. Het bereiken van de juiste balans tussen translatie en myogenese is net zo belangrijk als de engineering zelf.
"Om hoogwaardige CBM en de productie ervan met hoge opbrengst te bereiken, moet het moleculaire aspect grondig worden geïnspecteerd om goede laboratoriumpraktijken voor commerciële productie te bereiken." - Asim Azhar et al., Frontiers in Food Science and Technology [5]
Verschillende technieken hebben al veelbelovende resultaten laten zien bij het verhogen van de output van recombinant eiwit, zoals het overexpressen van translatie-initiatie factoren (eIF3i en eIF3c), codonoptimalisatie en het richten op mRNA-modificaties [15]. Deze methoden moeten echter met zorg worden toegepast om problemen zoals metabole belasting, proteostase stress en langdurige genetische instabiliteit te vermijden. Hoewel moleculaire optimalisatie essentieel is, kan het niet volledig de uitdagingen aanpakken zoals beperkingen in nutriëntendiffusie, gevoeligheid voor schuifspanning en verstoring van het proteoom tijdens de oogst.Deze hindernissen vereisen gelijktijdige vooruitgang in bioprocesontwerp.
De potentiële milieuvoordelen van gekweekt vlees zijn enorm. Het zou de uitstoot van broeikasgassen met 78%–96% kunnen verminderen, het landgebruik met 99% kunnen verminderen en het watergebruik met 82%–96% kunnen verlagen vergeleken met traditionele veeteelt [12]. Het bereiken van deze voordelen op schaal hangt af van het overbruggen van de kloof tussen de huidige productiviteit van celculturen en economische haalbaarheid. Ribosoomengineering is een krachtig hulpmiddel om deze kloof te helpen dichten, maar het moet deel uitmaken van een bredere, geïntegreerde aanpak die moleculaire biologie, bioprocesinnovaties en uitgebreide multi-omics monitoring omvat. Alleen door deze inspanningen te combineren kan de volledige belofte van gekweekt vlees worden gerealiseerd.
Hoe Cellbase Ondersteunt Onderzoek naar Gekweekt Vlees
Het bevorderen van moleculaire optimalisatie naar grootschalige productie in gekweekt vlees vereist nauwkeurige hulpmiddelen en materialen in elke fase.
Voor teams die werken aan cel-lijn optimalisatie, vereenvoudigt
Bij het opschalen van de productie biedt
Veelgestelde vragen
Welke ribosoom-engineeringbenadering is het meest veelbelovend voor gekweekte vleescellijnen?
Onderzoek naar ribosoom-engineering voor gekweekt vlees is gericht op het verbeteren van eiwitbiosynthese en het beïnvloeden van celbeslissingen. Een veelbelovende benadering is ribosoompool-engineering, die ribosomaal RNA-operons modificeert om de translatie-efficiëntie te verbeteren. Hulpmiddelen zoals iSAT en RISE bieden platforms voor in vitro ribosoom-evolutie, waardoor de ontwikkeling van ribosomen met verbeterde functionaliteit mogelijk wordt. Bovendien spelen platforms zoals
Hoe kunnen hogere translatiepercentages worden verhoogd zonder misgevouwen eiwitten of celstress te veroorzaken?
Om translatiepercentages te verbeteren zonder eiwitmisvouwing of cellulaire stress te veroorzaken, richten onderzoekers zich op het verfijnen van het translatieproces in plaats van het overal te versnellen. Enkele belangrijke benaderingen zijn:
- Gebruik van langzaam vertalende codons: Deze helpen de snelheid van translatie af te stemmen op het natuurlijke proces van eiwitvouwing, waardoor een juiste structuurvorming wordt gegarandeerd.
- Vermindering van vrije vouwenergie in de 5' coderingsregio: Deze aanpassing kan de efficiëntie van eiwitproductie verbeteren terwijl de cellulaire gezondheid behouden blijft.
Andere technieken omvatten lage inductieregimes, temperatuurdalingen, en geavanceerde synthetische hulpmiddelen zoals SINEUP RNA's. Deze strategieën maken hogere eiwitopbrengsten mogelijk zonder de cel te overbelasten.
Voor degenen die werken met gespecialiseerde materialen, kunnen bronnen zoals
Welke veranderingen zijn nodig in bioreactoren om ribosoom-gemodificeerd spierweefsel te ondersteunen dat dikker is dan 200 µm?
Om spierweefsel dikker dan 200 µm te laten groeien, moeten bioreactoren uitdagingen overwinnen met betrekking tot de diffusie van voedingsstoffen, zuurstof en pH - factoren die cruciaal zijn voor het overleven van cellen in driedimensionale structuren. Roerstoftankbioreactoren vereisen nauwkeurige aanpassingen om uniforme omstandigheden te handhaven terwijl de schuifspanning die cellen kan beschadigen, wordt verminderd. In veel gevallen spelen perfusie-gebaseerde systemen een sleutelrol bij het creëren van stabiele omgevingen, vooral in dicht opeengepakte weefsels. Voor degenen die werken met gespecialiseerde bioreactoren en materialen, biedt









