

Als je cellen kunt uitbreiden maar ze niet op het juiste moment in het juiste lot kunt veranderen, zal je proces vastlopen bij differentiatie. Dat is hier het kernpunt: synthetische genencircuits geven je binnen-de-cel controle over toewijding, timing, geheugen en afstammingsmix, waar alleen mediaveranderingen vaak heterogene, gedeeltelijk toegewijde populaties achterlaten.
Als ik een kweekvleesdifferentiatie-workflow zou opbouwen, zou ik meteen vier punten uit dit artikel halen:
- Begin met het native netwerk, niet met de constructie. Gebruik snRNA-seq, trajectanalyse, GRN-inferentie en miRNA-profielbepaling om te ontdekken waar cellen vastlopen, afdwalen of in het verkeerde lot vertakken.
-
Stem het type circuit af op het procesprobleem.
Een schakelschakelaar is geschikt voor lock-in, een feedforward- of band-passontwerp is geschikt voor timingcontrole, een logische poort is geschikt voor multi-signaal gating, en miSFITs zijn geschikt voor gegradeerde output. - Ontwerp vanaf dag één voor lage lekkage, weinig ruis en veiligheid. Orthogonale onderdelen, negatieve autoregulatie, iFFLs, cm transgenen en een induceerbare kill- of groeistopmodule maken deel uit van de constructie, niet een bijzaak.
- Valideer vroeg onder schaalrelevante omstandigheden. Een circuit dat werkt in 2D kan verschuiven in 3D, microcarrier of geroerde suspensie vanwege inductorgradiënten, zuurstoflimieten en schuifspanning.
Het artikel maakt ook een praktisch punt dat van belang is voor procesteams: enkelvoudige afstammingscontrole en verhoudingcontrole zijn verschillende taken. Een Tet-On MyoD cassette kan myogene instap bevorderen, maar whole-cut producten vereisen controle over spier-, vet- en ECM-verhoudingen, wat meestal feedback, paracriene signalering en intensievere kloonscreening betekent.
Enkele cijfers verankeren die boodschap:
- Standaard myogene differentiatie kan stagneren met fusie-indexen van ongeveer 50–60%
- Geëngineerde GRNs in iPSCs verhoogden de doel-lijn differentiatie van 52% naar 81%
- Synthetische circuits in gemodificeerde MSCs dreven cardiale differentiatie naar 76%
- Sommige porcine Tet-On-PAX7 lijnen behielden een hoog myogeen potentieel na 40 passages
- Ongeveer 20% van menselijke pluripotente stamcellen kan kanker-gerelateerde mutaties dragen, daarom zijn induceerbare veiligheidsmodules belangrijk
Synthetische Gen Circuit Workflow voor Gekweekt Vlees Differentiatie
Wetenschapper Verhalen: Michael Elowitz, Genetische Circuits in Levende Cellen
Snelle vergelijking
| Onderwerp | Wat het artikel in eenvoudige termen zegt |
|---|---|
| Belangrijkste knelpunt | Differentiatie, geen uitbreiding |
| Waarom circuits helpen | Ze voegen drempelcontrole en celtoestandgeheugen toe binnen de cel |
| Beste eerste stap | Kaart de controle over de oorspronkelijke bestemming met snRNA-seq en gerelateerde tools |
| Belangrijkste circuitopties | Toggle, feedforward, band-pass, logische poorten, miSFITs, CRISPRa/i |
| Myogenese voorbeeld | Tet-On-MyoD om groei te scheiden van terminale differentiatie |
| Adipogenese / ECM controle | miSFITs en bow-tie ontwerpen voor graduele PPARG/BMP4-type output |
| Whole-cut uitdaging | Verhouding controle over spier, vet en bindweefsel |
| Schaalvergrotingsrisico | 2D-gedrag houdt mogelijk niet stand in 3D of bioreactoren |
| Integratiekeuzes | Lentivirus, transposons, CRISPR knock-in, episomale vectoren |
| Reguleringspunt | Geëngineerde lijnen hebben een breder veiligheidspakket nodig; voedselveilige inductoren zoals vanillinezuur worden waar mogelijk geprefereerd boven DOX |
Dus, in eenvoudige termen: dit is niet alleen een artikel over circuitontwerp. Ik zou het lezen als een gids voor het koppelen van circuitarchitectuur, afstammingsbiologie, kloneselectie, bioreactorprestaties en UK/EU-veiligheidsdocumentatie in één differentiatie strategie.
Lees verder als je het volledige pad wilt van native netwerk mapping naar constructiekeuze, opschalingscontroles en regelgevingsgeschiktheid.
2. Ontwerpprincipes voor differentiatie-controle circuits
2.1 Kaart het native cel-lot netwerk in kaart voordat je het circuit ontwerpt
Voordat je een circuit ontwerpt, heb je een duidelijk beeld nodig van wat de cel al doet.
Single-nucleus RNA sequencing (snRNA-seq) is een goed startpunt. Het kan quiescente subpopulaties laten zien, inclusief reservecellen gemarkeerd door NOTCH2 en HEYL , en wijzen op paddoelen die de differentiatie kunnen verbeteren [3].
Van daaruit helpen trajectanalyse en genregulerend netwerk (GRN) inferentie om de volgorde van regulatoractivatie in kaart te brengen en te benadrukken waar cellen waarschijnlijk vastlopen. In myogenese loopt de hoofdcascade via MYOD1 en MYOG . In adipogenese zijn de belangrijkste knooppunten PPARG en CEBPA , met fibro-adipogene progenitor (FAP) lot als het belangrijkste vertakkingspunt risico. De onderstaande tabel vat de belangrijkste regulatoren en knelpunten samen.
| Afstamming | Belangrijke meesterregulatoren | Kritieke signaalroutes | Geïdentificeerde knelpunten |
|---|---|---|---|
| Myogeen | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Reservecelvorming (quiescentie) |
| Adipogeen | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Fibro-adipogene progenitor (FAP) lot |
| Pluripotent | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Spontane differentiatie / heterogeniteit |
Een andere nuttige laag is miRNA-expressieprofilering. Endogene miRNA's zoals miR-302a, gekoppeld aan pluripotentie, en miR-375, gekoppeld aan differentiatie, kunnen fungeren als interne classificatoren in sense-and-respond ontwerpen. Dat laat het circuit de werkelijke toestand van de cel lezen in plaats van alleen te vertrouwen op externe inductoren [5].
Die knelpunten zouden de circuitkeuze moeten bepalen. Als het belangrijkste probleem drift is, heeft u mogelijk een lock-in nodig. Als timing het probleem is, kan een puls beter passen. Als lotcontrole afhankelijk is van meer dan één aanwijzing, is multi-input logica meestal logischer.
2.2 Kies de juiste circuitarchitectuur
Dit is waar de afwegingen duidelijk zichtbaar worden. De juiste architectuur hangt af van drie praktische punten: hoe permanent de toewijding moet zijn, hoe nauwkeurig de timing moet worden gecontroleerd en hoeveel genetische lading de cel zonder problemen kan dragen.
Bistabiele tuimelschakelaars passen bij gevallen waar lijncommitment vergrendeld moet blijven. Het belangrijkste probleem is spontane toestandomschakeling veroorzaakt door transcriptionele ruis.
Banddoorfilters passen bij gevallen waarin een transcriptiefactor alleen binnen een bepaald ontwikkelingsvenster nodig is. Het probleem is dat de niveaus van de inductor nauwkeurig moeten worden gecontroleerd, anders verschuift de timing.
Logische poorten voegen specificiteit toe door meer dan één invoer tegelijkertijd te vereisen. Bijvoorbeeld, differentiatie kan alleen worden toegestaan wanneer een exogene inductor aanwezig is en de cel het juiste endogene miRNA-profiel vertoont. Dat helpt het risico op ongewenste toewijzing te verminderen.
De onderstaande tabel schetst de belangrijkste architecturen en de afwegingen die ermee gepaard gaan.
| Architectuur | Omkeerbaarheid | Tijdelijke precisie | Integratiecomplexiteit | Primaire gebruiksgeval | Belangrijkste risico |
|---|---|---|---|---|---|
| Bistabiele schakelaar | Laag (eenmaal vergrendeld) | Gemiddeld | Gemiddeld | Permanente afstammingsverbintenis | Spontane omschakeling door ruis |
| Banddoorlaatfilter | Hoog (concentratie-afhankelijk) | Hoog | Hoog | Voorbijgaande ontwikkelingsstadia | Vereist strakke inductorcontrole |
| Logische poort (EN/OF/NIET) | Variabel | Gemiddeld | Gemiddeld–hoog | Celtype-specifieke activering | Lekkage in UIT-staten |
| Vlinderdas / multi-input | Hoog | Gemiddeld | Gemiddeld | Multi-signaal integratie | Vertrouwt op endogene miRNA-stabiliteit |
| miSFITs | Hoog | Gemiddeld | Laag–gemiddeld | Geleidelijke output afstemming | Nauwe dynamische bereik als slecht afgestemd |
"Door het minimaliseren van het aantal computationele lagen terwijl de functionaliteit behouden blijft, pakt deze strategie schaalbaarheidsbarrières aan in genencircuitengineering." - Nature Communications [9]
Elke toegevoegde regelgevende laag verhoogt de belasting en put uit de cellulaire bronnen. In de praktijk, als twee ontwerpen hetzelfde werk doen, is de eenvoudigere meestal de betere keuze wanneer schaal belangrijk is.
Zodra de architectuur is vastgesteld, is de volgende taak om ervoor te zorgen dat deze standhoudt onder lage lekkage, ruisonderdrukking en fail-safe controle.
2.3 Bouw voor betrouwbaarheid, lage lekkage en veiligheid
Een circuit moet stabiel blijven gedurende een uitgebreide cultuur. Een korte uitbarsting van goede prestaties is niet genoeg voor productiegebruik.
Orthogonale onderdelen zijn de eerste verdediging. Promotoren, transcriptiefactoren en regelgevende elementen die niet kruisreageren met de native machinerie helpen om off-target effecten te beperken en verminderen de kans dat endogene signalen het circuit aanzetten of uitschakelen. Aangepaste promotoren met hoge strakheid zoals PCREm zijn gebruikt om de basale lekkage in induceerbare zoogdiersystemen te verminderen [6] .
Negatieve autoregulatie is ook de moeite waard om toe te voegen waar mogelijk. Het is een van de beter bekende motieven voor het verminderen van transcriptionele ruis en het produceren van een meer lineaire respons op de inductorconcentratie [6] . Incoherente feedforward-lussen (iFFLs) kunnen een extra controlelaag toevoegen door stochastische fluctuaties te filteren, zodat cellen reageren op aanhoudende signalen in plaats van op korte ruispieken.
Codon-aangepaste ( cm) versies van synthetische transcriptiefactoren maken karakterisering ook gemakkelijker. Ze laten je circuitgestuurde expressie scheiden van endogene genomische (g) expressie tijdens validatie [1]. Dat mag als een klein detail klinken, maar het bespaart tijd wanneer je probeert uit te zoeken of een uitlezing afkomstig is van het circuit of het gastheer-genoom.
Veiligheidsmodules zijn vereist. Ongeveer 20% van de menselijke pluripotente stamcellen dragen kanker-geassocieerde mutaties [7] . Dus, als een circuit in een stamcel-afgeleide lijn wordt geplaatst, moet het een induceerbare groeistop- of eliminatiemodule bevatten. Vanillinezuur is hier een nuttige inductor om te prioriteren omdat het een goedgekeurde voedseladditief is, wat helpt bij het gebruik ervan als een circuittrigger in gekweekte vleescellijnen [1] .
"Synthetische biologie biedt ingenieurs de middelen om circuits in te zetten om eenvoudig en nauwkeurig de expressie van meerdere genen af te stemmen om... mogelijke nadelige off-target bijwerkingen te elimineren." - npj Systems Biology and Applications [6]
Deze keuzes stellen de lijnspecifieke circuits in Sectie 3 in.
sbb-itb-ffee270
3. Circuitstrategieën voor myogene, adipogene en ratio-gecontroleerde differentiatie
3.1 Myogene circuits die expansie scheiden van terminale differentiatie
Zodra de circuitarchitectuur is ingesteld, is de volgende taak lijnspecifieke inzet. Voor myogenese is het hoofdprobleem eenvoudig te stellen maar moeilijk uit te voeren: cellen moeten eerst prolifereren en dan overschakelen naar differentiatie wanneer daarom wordt gevraagd, zonder te vroeg in beide richtingen af te wijken.
Een Tet-On-MyoD cassette is een van de meest directe manieren om dit te doen. In deze opstelling prolifereren cellen onder standaardomstandigheden wanneer doxycycline (DOX) afwezig is. Voeg DOX toe, en het circuit stuurt myogene toewijding aan.Onderzoekers van meerdere Chinese instellingen gebruikten deze aanpak in kippenembryonale fibroblasten en rapporteerden efficiënte myotubevorming na inductie [4] .
Een bistabiele tuimelschakelaar biedt strakkere controle over de toestand. Systemen opgebouwd uit wederzijds remmende repressors zoals E-KRAB en Pip-KRAB kunnen een korte DOX-puls omzetten in een stabiel myogeen programma [6] . In de praktijk betekent dit dat de gedifferentieerde toestand na inductie vergrendeld blijft in plaats van te vervagen zodra het signaal wordt verwijderd. Het toevoegen van negatieve autoregulatie helpt ook om stochastische ruis te verminderen en lekkende differentiatie tijdens de uitbreidingsfase te beperken [6].
Dit is belangrijk omdat standaard in vitro myogene differentiatie vaak halverwege stagneert. Fusie-indices zijn meestal slechts rond de 50–60%, wat een grote niet-fuserende reservepopulatie achterlaat [3]. Circuit-gestuurde MyoD-activatie kan de toewijding verbeteren, maar toewijding alleen garandeert geen uniforme fusie. Wanneer MyoD-circuits worden gecombineerd met MEK-, NOTCH- en RXR-modulatie, wordt bijna volledige fusie in 2D-cultuur mogelijk [3]. Het circuit controleert de timing van de lijninvoer; de kleine moleculen zorgen voor een schonere en meer uniforme celcyclus-exit.
3.2 Adipogene en fibrogene controle voor samenstelling en structuur
Spier alleen is niet genoeg. Zodra myogene controle is ingesteld, is het volgende probleem de samenstelling: hoeveel vet wordt gevormd, hoeveel ECM wordt afgezet en wanneer die programma's worden geactiveerd. Hier is binaire AAN/UIT-controle vaak te grof. Wat teams meestal nodig hebben, is een graduele output, vooral rond het adipogene-fibrogene vertakkingspunt.
miSFITs bieden een praktische manier om expressie in stappen af te stemmen.Door gemuteerde miRNA-doelsites te plaatsen - bijvoorbeeld, sites voor miR-17 - in de 3′UTR van outputgenen zoals PPARG of BMP4, kunnen onderzoekers expressieniveaus selecteren uit een variantbibliotheek [5]. Dat maakt adipocyteninductie meer als een dimmer dan een lichtschakelaar. In plaats van cellen in een alles-of-niets-reactie te duwen, kunnen teams adipogenese zorgvuldiger titreren [5].
Fibroblasten zijn hier ook niet alleen toeschouwers. Ze leveren ECM-eiwitten die de textuur vormen [10]. Dat maakt fibrogene controle onderdeel van productontwerp, niet slechts een bijzaak. Circuits kunnen helpen bij het beheren van de verschuiving tussen fibrogene en adipogene toestanden, en in gekweekt gevogelte kan directe PPARG-activering in fibroblasten nodig zijn om betekenisvolle vetafzetting te genereren [10].
Een bow-tie architectuur past goed bij dit probleem omdat het sensing en output gescheiden houdt. De sensinglaag leest de huidige toestand van de cel, terwijl de outputlaag PPARG, CEBPA of andere lijnregulatoren aanpast. Die scheiding helpt voorkomen dat adipogene of bindweefselprogramma's worden geactiveerd voordat cellen het juiste ontwikkelingsstadium hebben bereikt.
3.3 Multi-lijnverhouding controle en feedback sensing
Verhoudingscontroleschakelingen behandelen een ander probleem: niet of differentiatie plaatsvindt, maar of de uiteindelijke populatiemix blijft waar deze zou moeten zijn. Voor whole-cut producten is het net zo belangrijk om spier, vet en ECM in de juiste verhoudingen te krijgen als om een enkele lijn te laten differentiëren.
Deze systemen bouwen feedbackcontrole in de cellen zelf. Toestand-specifieke promotoren beperken de expressie van signaaleiwitten tot cellen die zich al aan een bepaalde lijn hebben gecommitteerd.Synthetische paracriene modules laten vervolgens toe dat toegewijde myogene cellen een remmend signaal afgeven dat de adipogene toewijding in nabijgelegen cellen onderdrukt. De logica lijkt op laterale inhibitie in Delta-Notch ontwikkelingssystemen [1] [6]. Waar vertakking ingewikkelder wordt, kunnen multi-input poorten extracellulaire signalen combineren met interne toestandsignalen [9].
miSFITs werken ook in dit stadium. Door de uitvoersterkte van BMP4 of andere morfogenen af te stemmen, kunnen teams de lijnbalans verschuiven zonder de upstream beslissingslogica te herschrijven. In ontworpen genregulerende netwerken verhoogde dit type controle de efficiëntie van doel-lijn differentiatie van 52% in controles tot 81% in ontworpen iPSCs [2]. In gemodificeerde mesenchymale stamcellen dreven synthetische circuits de efficiëntie van cardiale differentiatie naar 76% [2].
De onderstaande tabel vergelijkt single-lineage en ratio-control benaderingen met behulp van de punten die het meest van belang zijn in de productie.
| Kenmerk | Enkelvoudige afstammingscircuit | Verhoudingsregelcircuit |
|---|---|---|
| Complexiteit | Laag; meestal een enkele induceerbare promotor en regelaar [4] | Hoog; vereist logische poorten en paracriene signalering [6][9] |
| Monitoring last | Laag; volgt meestal een enkele reporter [4] | Hoog; vereist het volgen van meerdere afstammingsmarkers [5] |
| Robuustheid | Gemiddeld; gevoelig voor heterogeniteit en reservecelvorming [3] | Hoog; gebruikt feedback en laterale inhibitie om populaties in balans te houden [1] |
| Productiewaarde | Hoog voor biomassa; beperkt voor complexe weefselstructuur [10] | Essentieel voor whole-cut producten die spier, vet en ECM nodig hebben [4] |
Verhouding-controle circuits voegen een zwaardere validatielast toe. Maar hun ingebouwde feedback is moeilijk te evenaren met alleen procescontrole, wat meer druk legt op kloneselectie en procestesten.
4. Van constructie tot proces: validatie, opschaling en regelgevingsgeschiktheid
4.1 Integratiestrategie en kloneselectie voor stabiele prestaties
Na het ontwerp van het circuit begint het moeilijke deel: het circuit in cellen krijgen op een manier die stabiel blijft tijdens productie-schaal passages.
Lentivirale levering is vaak efficiënt en geeft snel stabiele integranten. Maar integratie is willekeurig. Dat betekent meer regelgevende aandacht, plus een risico dat de expressie na verloop van tijd afneemt door silencing. Transposonsystemen zoals PiggyBac en Sleeping Beauty zitten in het midden. Ze kunnen prestaties behouden over vele passages, maar je moet nog steeds screenen op kopieaantal en insertieplaatsen. Doornroosje, bijvoorbeeld, is gebruikt om runder satellietcellen stabiel te immortaleren door TERT en CDK4 te overexpressen, waarbij lijnen hun myogene potentieel behouden na meer dan 40 passages [10] . CRISPR knock-in biedt de strakste controle over waar het construct landt en de meest precieze genomische instelling, hoewel het selecteren van klonen langzamer is en de doorvoer lager.
| Integratiemethode | Invoegcontrole | Stabiliteit | Schaalbaarheid | Regelgevende Overwegingen |
|---|---|---|---|---|
| Lentivirale Levering | Laag (willekeurige integratie) | Hoog, maar gevoelig voor stillegging | Hoog | Grotere regelgevende controle vanwege willekeurige insertie en virale resten |
| Transposons (PiggyBac/SB) | Gemiddeld | Hoog over vele passages | Hoog | Vereist screening voor kopieaantal en insertieplaatsen |
| CRISPR Knock-in | Hoog (plaats-specifiek) | Zeer hoog | Gemiddeld | Gunstig; vermindert het risico op verstoring van endogene genen |
| Episomale Vectoren | Geen (extrachromosomaal) | Laag; kan verloren gaan tijdens deling | Laag | Lagere integratiedrempel, maar ongeschikt voor langdurige expansie |
Kloon screening moet meer doen dan alleen bevestigen dat het construct aanwezig is.Het moet expressiedrift, insertieprofiel, groeikinetiek, differentiatie-efficiëntie en fenotypebehoud bij een hoog passagegetal volgen. snRNA-seq is hier nuttig omdat het klonen kan uitsluiten die verrijkt zijn voor Pax7⁺/Ki-67⁻ reservecellen - cellen die de celcyclus verlaten zonder te differentiëren - voordat opschaling plaatsvindt [3]. Porcine EPSCs met een Tet-On-PAX7 circuit behielden hoge spierdifferentiatie in 3D microcarrier en suspensiecultuur over 40 passages [8].
4.2 Hoe circuitgedrag verandert in 3D, microcarrier en bioreactorcultuur
Zodra je een kloon hebt, is de volgende test of deze zich buiten 2D op dezelfde manier gedraagt. In veel gevallen is dat niet zo. Prestaties in 2D worden zelden schoon overgedragen naar suspensie-, microcarrier- of scaffoldcultuur omdat diffusiegradiënten, zuurstoflimieten en schuif allemaal de circuitoutput verschuiven.
Een van de eerste controles is de diffusie van de inductor. In geroerde-tank bioreactoren moeten kleine-molecuul inductoren gelijkmatig de cellen bereiken. In de praktijk kunnen er gradiënten ontstaan, vooral in dichte microcarrier culturen en binnenin aggregaten of scaffoldkernen. Suspensie cultuur is meestal geschikter voor grootschalig gekweekt vlees omdat het een hogere celdichtheid ondersteunt en strakkere procescontrole biedt.
Het monitoren van de celtoestand wordt ook moeilijker naarmate het systeem opschaalt. Fluorescente reporter signalen die gemakkelijk te lezen zijn met microscopie in 2D kunnen moeilijk te onderscheiden worden in ondoorzichtige 3D constructies. Fluorescente timers - sondes die emissie verschuiven van blauw naar rood naarmate een eiwit rijpt - kunnen real-time circuit-activatiegegevens geven in situ [1]. De verstandige route is gefaseerde validatie: eerst in 2D, dan in 3D constructies, en vervolgens onder de uiteindelijke bioreactoromstandigheden [3] [8].
4.3 Karakterisering, voedselveiligheidsdocumentatie en UK/EU overwegingen
Na procestesten moet karakterisering aantonen dat circuitfunctie, fenotype en veiligheid nog steeds standhouden. De kerngegevensstapel moet flowcytometrie, qPCR met cm-sequenties, RNA-seq tijdreeksen en functionele uitlezingen zoals myosine zware keten gebiedsfractie en myoglobine-expressie bevatten [1]. Geoptimaliseerde serumvrije differentiatiemedia hebben aangetoond dat myoglobine-expressie tot ongeveer 30% van de niveaus in native runderspier kan worden gebracht [3]. Dat geeft teams een duidelijke benchmark in plaats van een vaag doel.
U moet ook de eiwit-, aminozuur- en vetprofielen documenteren, samen met sensorische eigenschappen [10][3].
Vanuit een regelgevend oogpunt maken het VK en de EU een duidelijk onderscheid tussen spontaan geïmmortaliseerde (niet-GMO) cellijnen en genetisch gemodificeerde lijnen. Voor de laatste zijn bredere veiligheidsdossiers nodig [10][3]. Stabiliteitspakketten moeten fenotypebehoud en genomische stabiliteit aantonen over de volledige productieketen - van de mastercelbank tot de uiteindelijke productiecel - en traceerbaarheidsrecords moeten rekening houden met elke passage daartussen [10]. Als het circuit afhankelijk is van een chemische inductor, is een voedselveilige of gelicentieerde toevoeging zoals vanillinezuur te verkiezen boven doxycycline [1].
Routine genomische surveillance is een must, en een induceerbare zelfmoord- of eliminatieschakelaar moet worden gedocumenteerd als een kernmaatregel voor risicobeheersing [7]. De functie ervan moet ook worden opgenomen in het veiligheidsdossier, vooral omdat de regels voor gekweekt vlees in het VK en de EU vorm blijven krijgen.
5. Praktische routekaart en conclusie
5.1 Een gefaseerde implementatieroutekaart voor teams die gekweekt vlees produceren
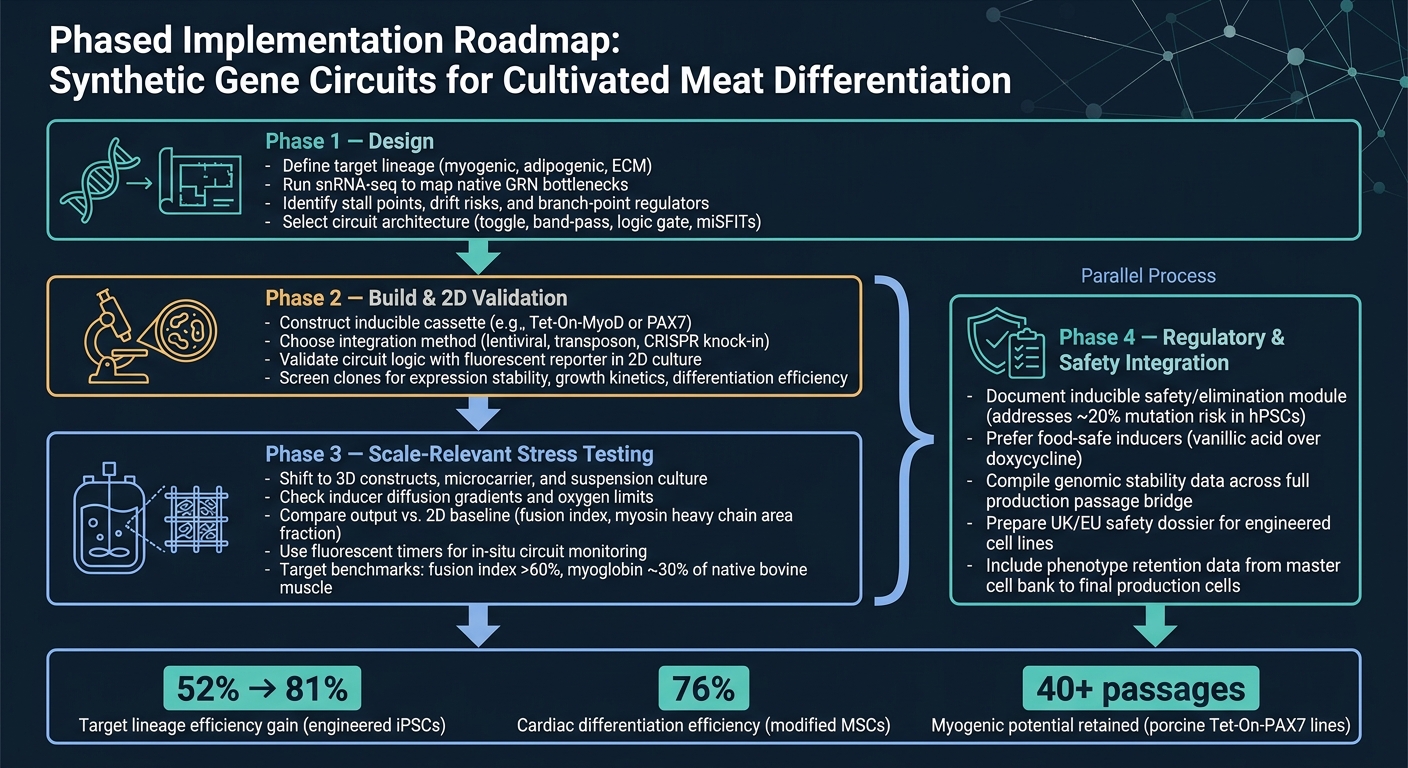
De schoonste route van concept tot productie is een gefaseerde workflow.
Fase 1 is ontwerp. Begin met het definiëren van de doelstam, gebruik vervolgens snRNA-seq om de belangrijkste knelpunten te bevestigen voordat je een circuitarchitectuur kiest. Die stap is belangrijk omdat een circuit alleen de beperkingen kan oplossen die je daadwerkelijk hebt geïdentificeerd.
Fase 2 is bouwen en 2D-validatie. Bouw het construct en controleer of het circuit zich gedraagt zoals bedoeld in 2D, met behulp van een eenvoudige reporter-uitlezing.In dit stadium is het doel eenvoudig: bevestig dat de logica werkt voordat u overgaat naar moeilijkere en duurdere modellen.
Fase 3 is schaalrelevante stresstests. Schakel over naar 3D-systemen en bioreactor-relevante omstandigheden, en vergelijk vervolgens de output met de 2D-baseline. Dit is waar veel ontwerpen hun zwakke punten beginnen te vertonen, vooral wanneer massatransfer, schuifspanning en matrixeffecten een rol gaan spelen.
Fase 4 is integratie van regelgeving en veiligheid, en het moet parallel lopen met Fase 3. Veiligheids- en regelgevingswerkzaamheden moeten niet tot het einde wachten. Voer het uit naast opschaling, inclusief documentatie voor elk induceerbaar veiligheidsmodule.
5.2 Het verkrijgen van hulpmiddelen en materialen via Cellbase
Zodra de workflow is ingesteld, wordt sourcing vaak de beperkende factor.
- cellijnen
- serumvrije en chemisch gedefinieerde media
- steigers
- bioreactorcomponenten
- sensoren
- analytische apparatuur
Betrouwbare toegang tot compatibele materialen in elke fase heeft een direct effect op hoe snel circuitgedrag kan worden gekarakteriseerd onder schaalrelevante omstandigheden.
5.3 Belangrijkste punten
Synthetische genencircuits geven teams voor gekweekt vlees programmeerbare controle over timing, drempels en lijnbalans die alleen-media-protocollen niet kunnen evenaren. De keuze van de architectuur bepaalt omkeerbaarheid, lekkage en veiligheid.Induceerbare systemen worden meestal geprefereerd omdat ze voorwaardelijke controle en een lagere metabolische belasting bieden [6].
"De synthetische biologie toolkit kan worden gebruikt om cellijnen met afstembare genexpressie op te zetten, die, in combinatie met PAT en computationele modellering, gesloten-lus controlesystemen kunnen mogelijk maken om optimale productopbrengst en kwaliteit te leveren." - npj Systems Biology and Applications [6]
Succesvolle implementatie is niet alleen een biologisch probleem. Het hangt af van nauwe koppeling tussen circuit engineering, bioprocesontwerp, regelgevende documentatie en inkoop.
Veelgestelde vragen
Hoe verbeteren synthetische genencircuits de consistentie van differentiatie?
Synthetische genencircuits kunnen differentiatie consistenter maken omdat ze je geprogrammeerde controle geven over celgedrag en lijncommitment.In de praktijk betekent dit het gebruik van modulaire logische operaties om de expressie van genen en transcriptiefactoren met strakke timing af te stemmen.
Die timing is belangrijk. Het helpt cellen om door gedefinieerde toestandveranderingen in de juiste volgorde te gaan, in plaats van af te drijven naar gemengde of ongewenste toestanden. Het vermindert ook off-target differentiatie en vermindert ruis in de cultuur.
Het resultaat is eenvoudig: meer uniforme, stabiele en volwassen celpopulaties voor de productie van gekweekt vlees.
Welk circuitontwerp is geschikt voor myogene of adipogene controle?
In onderzoek naar gekweekt vlees kunnen dezelfde kipfibroblasten in beide afstammingen worden geduwd. Myogenese volgt een reeks inductieprotocollen, terwijl adipogenese kan worden geactiveerd door de cellen bloot te stellen aan inputs zoals kipserum of vetzuren.
Vanaf daar kunnen deze celbestemmingen stap voor stap worden gecontroleerd binnen 3D-hydrogelstructuren om vleesstructuren te bouwen met gedefinieerde vet- en collageenverhoudingen.
Waarom gedragen genencircuits zich vaak anders in 3D-cultuur?
In 3D-cultuur , gedragen genencircuits zich vaak anders omdat cellen te maken hebben met fysieke en structurele inputs die simpelweg niet bestaan in 2D-monolagen. Deze inputs omvatten mechanische spanning, schuifspanning, matrixstijfheid en lokale celdichtheid.
Deze signalen kunnen signaalroutes zoals Notch verschuiven. Ze kunnen ook veranderen hoe synthetische circuits kracht detecteren en coördineren downstream reacties, inclusief cel-cel adhesie en weefselmorfogenese.









