

สำหรับนักวิจัยในการผลิตเนื้อสัตว์ที่เพาะเลี้ยง การลดการเกิดอะพอพโทซิสเป็นสิ่งสำคัญในการปรับปรุงความมีชีวิตของเซลล์และผลผลิตในเครื่องปฏิกรณ์ชีวภาพ ปัจจัยกดดันเช่น การขาดแคลนสารอาหาร ความไม่สมดุลของออสโมติก และการสะสมของของเสียมักจะกระตุ้นการตายของเซลล์ ทำให้ผลผลิตลดลง ยีนต้านอะพอพโทซิสสามารถบรรเทาความท้าทายเหล่านี้ได้โดยการยืดอายุของเซลล์ในระหว่างการเพาะเลี้ยง นี่คือภาพรวมอย่างรวดเร็วของยีนชั้นนำและบทบาทของพวกมัน:
- BCL-2: ป้องกันการก่อตัวของรูในไมโทคอนเดรีย โดยการบล็อกอะพอพโทซิสตั้งแต่เริ่มต้น มีประสิทธิภาพสำหรับเซลล์ที่ยังไม่แตกต่าง แต่ต้องมีการปรับสมดุลกับโปรตีนที่ส่งเสริมอะพอพโทซิสอย่างระมัดระวัง
- BCL-xL: ปกป้องเซลล์ในระหว่างการแตกต่างและสนับสนุนการเผาผลาญพลังงาน เหมาะสำหรับช่วงที่มีความเครียดสูงในเครื่องปฏิกรณ์ชีวภาพ
- MCL-1: ตอบสนองอย่างรวดเร็วต่อการเปลี่ยนแปลงของสารอาหารและคงที่ในระหว่างการแตกต่าง ทำงานได้ดีเมื่อใช้ร่วมกับยีนอื่นๆ
- BIRC5 (Survivin) : ยับยั้ง caspases เพื่อป้องกันการเกิด apoptosis ในขั้นตอนถัดไป สนับสนุนการแบ่งตัวในเซลล์ที่แบ่งตัวอย่างรวดเร็ว
- XIAP: เป็นตัวยับยั้ง caspase ที่มีประสิทธิภาพภายใต้สภาวะความเครียดสูง เช่น การเพาะเลี้ยงที่มีความหนาแน่นสูง การตรวจสอบสภาวะเหล่านี้ต้องการ การเลือกเซ็นเซอร์สำหรับเครื่องปฏิกรณ์ชีวภาพเนื้อสัตว์เพาะเลี้ยง เพื่อติดตามระดับสารอาหารและการสะสมของของเสียแบบเรียลไทม์
การเปรียบเทียบอย่างรวดเร็ว
| ยีน | บทบาทสำคัญ | ความเสถียรระหว่างการแยกตัว | กรณีการใช้งานที่ดีที่สุด |
|---|---|---|---|
| BCL-2 | บล็อกการตายของเซลล์ในระยะแรก (BAX/BAK) | เสถียร | การรักษาเซลล์ที่ยังไม่แยกตัว |
| BCL-xL | ป้องกันการกระตุ้น caspase, สนับสนุนการเผาผลาญ | เฉพาะระยะ | เซลล์ที่แยกตัวภายใต้ความเครียด |
| MCL-1 | ตอบสนองอย่างรวดเร็วต่อการเปลี่ยนแปลงของสารอาหาร | เสถียร | การอยู่รอดหลายระยะ |
| BIRC5 | ยับยั้ง caspases ที่ปลายน้ำ | ลดลงเมื่อแยกตัว | เซลล์ที่แบ่งตัวอย่างรวดเร็ว |
| XIAP | การยับยั้ง caspase อย่างกว้างขวาง | เสถียร | สภาวะเครื่องปฏิกรณ์ชีวภาพที่มีความเครียดสูง |
1.BCL-2
BCL-2 เป็นยีนต้านการตายของเซลล์ที่ได้รับการวิจัยอย่างดีซึ่งมีบทบาทสำคัญในเส้นทางการตายของเซลล์แบบภายใน (ไมโตคอนเดรีย) เส้นทางนี้เป็นกลไกหลักของการตายของเซลล์ ซึ่งมักถูกกระตุ้นในเซลล์เนื้อสัตว์ที่เพาะเลี้ยงภายใต้ความเครียดของเครื่องปฏิกรณ์ชีวภาพ เช่น การขาดสารอาหารหรือระดับออกซิเจนต่ำ
BCL-2 ทำงานโดยการจับและทำให้โปรตีนที่ส่งเสริมการตายของเซลล์ เช่น BAX และ BAK เป็นกลาง การกระทำนี้ป้องกันการก่อตัวของรูขุมขนในไมโตคอนเดรีย หยุดการปล่อยไซโตโครมซี และหยุดการเกิดการตายของเซลล์ในขั้นตอนต่อไป กลไกนี้มีความสำคัญต่อการยืดอายุการใช้งานของเซลล์ในกระบวนการผลิตเนื้อสัตว์ที่เพาะเลี้ยง ดังที่ Rønning SB et al. อธิบาย:
"อัตราส่วนระหว่าง Bcl-2 และ Bax กำหนดความไวของเซลล์ต่อการเกิดการตายของเซลล์"[5]
นอกเหนือจากบทบาทในไมโตคอนเดรียแล้ว BCL-2 ยังอยู่ในเอนโดพลาสมิกเรติคูลัม (ER) ด้วยHere, it reduces calcium levels and inhibits IP3 receptor-mediated calcium release, mitigating calcium-induced apoptosis – a frequent issue in high-density bioreactor cultures[4]. Managing these scaling challenges is a primary focus for the industry. This dual localisation enables BCL-2 to protect cells from multiple apoptosis triggers.
The molecular structure of BCL-2, consisting of an eight-alpha-helix bundle and four well-defined BH domains, makes it an e
อย่างไรก็ตาม มีข้อควรระวังที่สำคัญ: การสมดุลระหว่าง BCL-2 และโปรตีนที่กระตุ้นการตายของเซลล์ เช่น BAX ต้องได้รับการจัดการอย่างระมัดระวัง แม้แต่ระดับการแสดงออกของ BCL-2 ที่สูงก็อาจไม่สามารถป้องกันการตายของเซลล์ได้หากสัญญาณการตายของเซลล์มีความแข็งแกร่งเกินไป[2]. การตรวจสอบสมดุลนี้เป็นสิ่งสำคัญสำหรับการบรรลุความมีชีวิตของเซลล์ที่เหมาะสม
sbb-itb-ffee270
2. BCL-xL
BCL-xL, ที่ถูกเข้ารหัสโดยยีน BCL2L1 มีบทบาทสำคัญในครอบครัว BCL-2 โดยการตั้งอยู่ที่เยื่อหุ้มไมโตคอนเดรียภายนอกและป้องกันการตายของเซลล์ โดยทำได้โดยการต่อต้านโปรตีนที่กระตุ้นการตายของเซลล์ เช่น BAX และ BAKนอกจากนี้ มันยังยับยั้ง caspase-3 (CASP3) ที่ถูกตัด ซึ่งมีความสำคัญในการหยุดการตายของเซลล์ กลไกนี้มีคุณค่าอย่างยิ่งใน การเพาะเลี้ยงในไบโอรีแอคเตอร์ที่มีความหนาแน่นสูง , ซึ่งความเครียดทางเมตาบอลิซึมสามารถคุกคามความมีชีวิตของเซลล์ได้
น่าสนใจที่กิจกรรมของ BCL-xL สอดคล้องกับขั้นตอนเฉพาะของการแยกตัว ในบางช่วงการแสดงออกของมันจะเพิ่มขึ้น ในขณะที่โปรตีนต้านการตายอื่น ๆ เช่น BCL-2 และ MCL-1 ยังคงไม่เปลี่ยนแปลง สิ่งนี้เน้นย้ำถึงความสำคัญของมันในการรักษาการอยู่รอดของเซลล์ในระหว่างการแยกตัว ตามที่ระบุไว้ใน Cell Death & Disease:
"BCL-xL/BCL2L1 เป็นโปรตีนต้านการตายที่สำคัญที่ส่งเสริมการอยู่รอดของเซลล์ที่กำลังแยกตัว..." [2]
นอกเหนือจากบทบาทในการตายของเซลล์ BCL-xL ยังสนับสนุนการเผาผลาญพลังงานของเซลล์ มันช่วยเพิ่มทั้งไกลโคไลซิสและการฟอสโฟรีเลชันออกซิเดทีฟ เพื่อให้แน่ใจว่ามีกิจกรรมเมตาบอลิซึมสูงการยับยั้ง BCL-xL ได้แสดงให้เห็นว่าลดการแสดงออกของยีนเมตาบอลิซึมและลดการหายใจของไมโทคอนเดรียทั้งในระดับพื้นฐานและสูงสุด ฟังก์ชันนี้มีความสำคัญอย่างยิ่งสำหรับเซลล์เนื้อสัตว์ที่เพาะเลี้ยงซึ่งพึ่งพาผลผลิตเมตาบอลิซึมที่ยั่งยืน
BCL-xL เข้ากันได้ดีกับกลยุทธ์การแก้ไขยีนที่ใช้กันทั่วไปในการวิจัยเนื้อสัตว์ที่เพาะเลี้ยง เทคนิคเช่นการถ่ายโอน lentiviral ช่วยให้สามารถรวมยีน BCL2L1 ได้อย่างเสถียร ในขณะที่ระบบ CRISPR/Cas9 ที่สามารถเหนี่ยวนำด้วย doxycycline ให้การควบคุมชั่วคราวที่แม่นยำต่อการแสดงออกของมัน [2] [6]. ระดับความแม่นยำนี้มักจะถูกจัดการผ่าน ซอฟต์แวร์ควบคุมกระบวนการชีวภาพ. คุณสมบัติเหล่านี้ทำให้ BCL-xL เป็นผู้สมัครที่แข็งแกร่งสำหรับการปรับปรุงความมีชีวิตของสายเซลล์ในการผลิตเนื้อสัตว์ที่เพาะเลี้ยง
สำหรับขั้นตอนการแยกแยะที่มีความต้องการเมตาบอลิซึมสูง BCL-xL อาจมีประสิทธิภาพมากกว่า BCL-2นักวิจัยสามารถใช้ตัวยับยั้ง WEHI-539 เพื่อทดสอบการพึ่งพา BCL-xL ของสายเซลล์ก่อนที่จะดำเนินการดัดแปลงพันธุกรรมถาวร [2]. นอกจากนี้ การแสดงออกของ BCL-xL ร่วมกับ MCL-1 อาจช่วยปรับปรุงการอยู่รอดของเซลล์ได้มากขึ้น เนื่องจากโปรตีนเหล่านี้ได้รับการสังเกตว่าทำงานร่วมกันในเซลล์บางประเภทที่ต้านทาน [6].
3. MCL-1
MCL-1 (Myeloid Cell Leukaemia-1) มีบทบาทสำคัญในการควบคุมเส้นทางการตายของเซลล์แบบภายใน พบที่เยื่อหุ้มไมโตคอนเดรียด้านนอก มันป้องกันการตายของเซลล์โดยการจับและกักเก็บโปรตีนที่ส่งเสริมการตายของเซลล์ BAX และ BAK หยุดการรวมตัวและการซึมผ่านของเยื่อหุ้ม การกระทำนี้จะบล็อกการปล่อยไซโตโครม c หยุดกระบวนการตายของเซลล์ก่อนที่จะถึงขั้นตอนการดำเนินการ [8] . นอกจากนี้ MCL-1 ยังจับกับโปรตีน BH3-only เช่น Bim, PUMA และ NOXA ด้วยความสัมพันธ์สูง [8]. เช่นเดียวกับ BCL-2 และ BCL-xL, MCL-1 มีความสำคัญในการต่อต้านสัญญาณการตายของเซลล์ โดยเฉพาะในช่วงที่มีความเครียดในไบโอรีแอคเตอร์
หนึ่งในคุณสมบัติที่โดดเด่นของ MCL-1 คือครึ่งชีวิตที่สั้น ทำให้การแสดงออกของมันตอบสนองต่อความพร้อมของสารอาหารและสัญญาณเมตาบอลิกได้อย่างรวดเร็ว โดยเฉพาะผ่านทางเส้นทาง AMPK/mTOR การศึกษาชี้ให้เห็นว่าการลดการบริโภคแคลอรี่ลง 25% สามารถลดการแปล MCL-1 ได้ประมาณ 39% ± 10% [7]. ความไวนี้มีความสำคัญอย่างยิ่งสำหรับการผลิตเนื้อสัตว์ที่เพาะเลี้ยง ซึ่งการเปลี่ยนแปลงในองค์ประกอบของสื่อการเจริญเติบโตหรือการขาดแคลนสารอาหารในระหว่างการเพาะเลี้ยงแบบแขวนลอยขนาดใหญ่ (ซึ่งต้องมีการวางแผนการผลิตอย่างรอบคอบการวางแผนขนาดการผลิต) สามารถลดระดับ MCL-1 ลงได้อย่างมากการลดลงดังกล่าวส่งผลกระทบต่อความมีชีวิตของเซลล์ ทำให้การปรับปรุงใน IVCC (ความเข้มข้นของเซลล์ที่มีชีวิตรวม) ที่ได้จากกลยุทธ์ต่อต้านการตายของเซลล์ลดลง เพื่อบรรเทาปัญหานี้ สูตรอาหารที่ปราศจากเซรั่ม ที่สนับสนุนกิจกรรม mTORC1 ที่แข็งแกร่งเป็นสิ่งจำเป็น [7] .
คุณสมบัติที่น่าสังเกตอีกประการหนึ่งของ MCL-1 คือความเสถียรในระหว่างการแยกแยะ ในโมเดลต้นกำเนิดตับอ่อน การแสดงออกของ MCL-1 ยังคงที่ตลอดโปรโตคอลการแยกแยะ 17 วัน ซึ่งแตกต่างจาก BCL-xL ที่แสดงการเปลี่ยนแปลงตามขั้นตอน [2]. ความเสถียรนี้ทำให้ MCL-1 มีประโยชน์อย่างยิ่งสำหรับการประยุกต์ใช้เนื้อสัตว์ที่เพาะเลี้ยง ซึ่งเซลล์จำเป็นต้องอยู่รอดในหลายขั้นตอนของการเจริญเติบโตโดยไม่ต้องการการแทรกแซงที่ต้องการเวลาอย่างแม่นยำ&
เครื่องมือแก้ไขยีนสามารถใช้ในการปรับเปลี่ยน MCL-1 เช่นเดียวกับยีนต่อต้านการตายของเซลล์อื่น ๆ ทำให้เป็นเป้าหมายที่หลากหลายสำหรับการวิศวกรรมสายเซลล์
เมื่อใช้ร่วมกับยีนต้านการตายของเซลล์อื่น ๆ MCL-1 มอบประโยชน์เพิ่มเติม ตัวอย่างเช่น การจับคู่ MCL-1 กับ BCL-xL ได้แสดงผลเสริมฤทธิ์ - การยับยั้งโปรตีนทั้งสองพร้อมกันลด EC50 ของยารอดชีวิตจากประมาณ 10 μM เหลือน้อยกว่า 20 nM [6]. วิธีการนี้สามารถปรับปรุงการรอดชีวิตของเซลล์ได้อย่างมีนัยสำคัญในช่วงที่มีความเครียดสูงของการผลิตเนื้อสัตว์ที่เพาะเลี้ยง
4. BIRC5 (Survivin)
BIRC5, มักเรียกว่า Survivin เป็นสมาชิกของครอบครัวโปรตีน Inhibitor of Apoptosis (IAP) [2]. แตกต่างจากโปรตีนครอบครัว BCL-2 ซึ่งทำงานที่เยื่อหุ้มไมโตคอนเดรียเพื่อป้องกันการเริ่มต้นของการตายของเซลล์ BIRC5 ทำงานในขั้นตอนที่ลึกกว่า มันบล็อก caspases ที่รับผิดชอบในการดำเนินการตายของเซลล์ ทำหน้าที่เป็นแนวป้องกันสุดท้ายต่อการตายของเซลล์ที่โปรแกรมไว้ [10].
ในวัฒนธรรมการแขวนลอย, ปัจจัยกดดันเช่น การขาดแคลนสารอาหาร, การสะสมของเสียจากการเผาผลาญ, และความเครียดจากแรงเฉือนทางกล สามารถกระตุ้นการตายของเซลล์แบบ apoptosis ได้ โดยการยับยั้งกิจกรรมของ caspase ในขั้นตอนนี้, การแสดงออกมากเกินไปของ BIRC5 ช่วยยืดอายุการใช้งานและประสิทธิภาพของเซลล์ ซึ่งส่งผลให้เกิดการปรับปรุงใน ค่าเชิงบูรณาการของความเข้มข้นของเซลล์ที่มีชีวิต - ซึ่งเป็นตัวชี้วัดสำคัญสำหรับการเพิ่มประสิทธิภาพการเพาะเลี้ยงเซลล์ [9] . Eric Baek, นักวิจัยที่ KAIST, อธิบายว่า:
"การปรับปรุงค่าเชิงบูรณาการของความเข้มข้นของเซลล์ที่มีชีวิตโดยการเอาชนะการตายของเซลล์, โดยเฉพาะ apoptosis, เป็นหนึ่งในกลยุทธ์ที่ใช้กันอย่างแพร่หลายที่สุดสำหรับการผลิตโปรตีนบำบัด [และเซลล์] อย่างมีประสิทธิภาพ" [9]
การแทรกแซงในขั้นตอนปลายน้ำนี้แสดงให้เห็นว่าสามารถเพิ่มผลผลิตของไบโอรีแอคเตอร์ในสายเซลล์เนื้อสัตว์ที่เพาะเลี้ยง รวมถึง เซลล์ดาวเทียมสุกร และ ไมโอบลาสต์ของโค.
กลยุทธ์ที่มีประสิทธิภาพที่สุดเกี่ยวข้องกับ การวิศวกรรมแบบผสมผสาน, การจับคู่ BIRC5 กับสารป้องกันไมโทคอนเดรีย เช่น BCL-2 หรือ BCL-xL ศาสตราจารย์ Michael Betenbaugh จาก มหาวิทยาลัยจอห์นฮอปกินส์ เน้นย้ำถึงวิธีการนี้:
"กลยุทธ์ที่บล็อกการตายของเซลล์ในหลายจุดตามลำดับอาจจำกัดการขยายสัญญาณการตายของเซลล์แบบอะพอพโทซิสเหล่านี้" [10]
โดยการรวมการยับยั้งแคสเปสของ BIRC5 เข้ากับการป้องกันไมโทคอนเดรียในขั้นต้น นักวิจัยสามารถสร้างการป้องกันหลายชั้นต่อการตายของเซลล์แบบอะพอพโทซิสได้
BIRC5 ยังผสานเข้ากับกระบวนการแก้ไขยีนได้อย่างราบรื่นCRISPR/Cas9 เป็นวิธีการชั้นนำในการสร้างเซลล์ไลน์ที่มีการแสดงออกมากเกินไป [9], แม้ว่านิวคลีเอสสังกะสีจะเป็นทางเลือกที่แม่นยำ siRNA สามารถใช้สำหรับการตรวจสอบเส้นทางก่อนที่จะผูกพันกับการรวมตัวทางพันธุกรรม [9].
5. XIAP
XIAP (X-linked inhibitor of apoptosis) ได้รับการยอมรับว่าเป็นตัวยับยั้ง caspase ที่มีประสิทธิภาพมากที่สุดในตระกูล IAP (inhibitor of apoptosis protein) ร่วมกับยีนเช่น BCL-2 และ MCL-1, XIAP มีบทบาทสำคัญในการกำหนดเป้าหมายการตายของเซลล์ในระยะการดำเนินการ ดังที่ได้เน้นใน Genes & Development :
"XIAP ถือว่าเป็นตัวยับยั้ง caspase ที่มีประสิทธิภาพมากที่สุดในหลอดทดลอง" [12]
XIAP ใช้กลไกที่แตกต่างกันสองแบบในการยับยั้งการตายของเซลล์ ประการแรก โดเมน BIR2 และบริเวณลิงเกอร์ของมันจะบล็อก caspases-3 และ -7 ที่เป็นตัวกระทำลำดับที่สอง โดเมน BIR3 ของมันยับยั้ง caspase-9 ซึ่งมีผลในการหยุดเส้นทางการตายของเซลล์แบบไมโทคอนเดรียภายใน นอกจากนี้ โดเมน RING ที่ปลาย C ของมันยังช่วยในการยูบิควิติเนชันและการสลายโปรตีโอโซมของ caspases เป้าหมาย [11]. โดยการแทรกแซงในเส้นทางการตายของเซลล์ทั้งภายในและภายนอก XIAP พิสูจน์ได้ว่ามีประสิทธิภาพสูงในการจัดการกับตัวกระตุ้นการตายของเซลล์ เช่น การขาดแคลนสารอาหาร ผลพลอยได้จากการเผาผลาญ และความเครียดทางกล - ปัจจัยที่พบได้บ่อยใน ระบบการผลิตเนื้อสัตว์ที่เพาะเลี้ยง. การทำงานของมันยังได้รับการเสริมด้วยการอนุรักษ์ที่แข็งแกร่งข้ามสายพันธุ์
ตัวอย่างเช่น XIAP ของมนุษย์มีความเหมือนกันของโปรตีน 87.7% กับ Bos taurus (โค) และ 89.5% กับ Mus musculus (หนู) [11] . ความคล้ายคลึงกันสูงนี้ทำให้การวิจัยจากระบบโมเดลสัตว์เลี้ยงลูกด้วยนมสามารถนำไปใช้กับสายเซลล์ที่ใช้ในการผลิตเนื้อสัตว์เพาะเลี้ยงได้อย่างน่าเชื่อถือ
XIAP สามารถถูกควบคุมโดยใช้เครื่องมือเช่น shRNA, antisense oligonucleotides, หรือ CRISPR/Cas9 [11]. ภายใต้ความเครียดที่รุนแรง โดเมน RING ของมันอาจกระตุ้นการ ubiquitination ตัวเอง [12], ในขณะที่ตัวยับยั้งภายในเช่น SMAC/DIABLO และ HTRA2 สามารถแทนที่ XIAP จาก caspases [11][13]. การค้นพบเหล่านี้ทำให้ XIAP เป็นเป้าหมายที่น่าสนใจสำหรับวิธีการแก้ไขยีนที่มุ่งเน้นการเพิ่มประสิทธิภาพสายเซลล์สำหรับการพัฒนาเนื้อสัตว์เพาะเลี้ยง
การเปรียบเทียบยีนต้านการตายของเซลล์อย่างรวดเร็ว
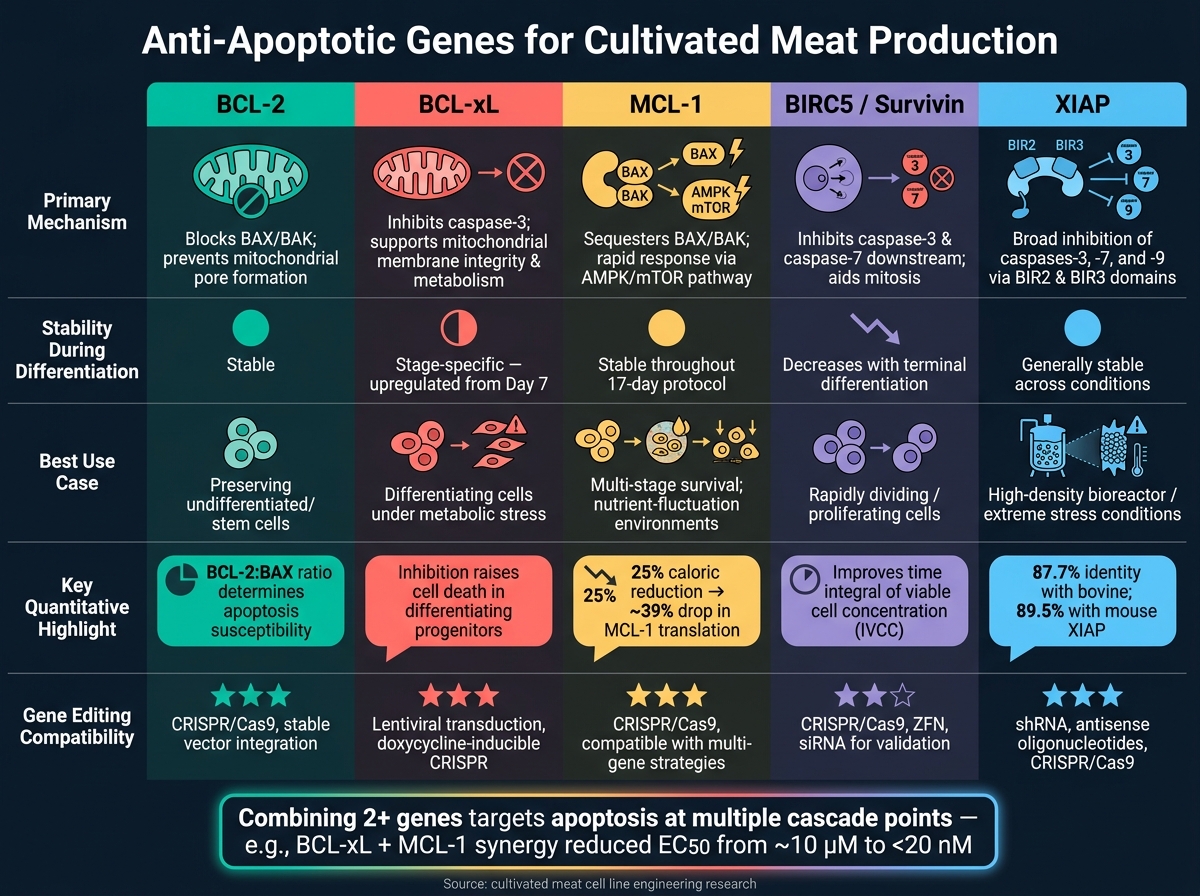
ยีนต้านการตายของเซลล์สำหรับเนื้อสัตว์เพาะเลี้ยง: การเปรียบเทียบแบบเคียงข้างกัน
เมื่อทำงานเกี่ยวกับการผลิตเนื้อสัตว์เพาะเลี้ยง การเข้าใจว่ายีนต้านการตายของเซลล์ต่างๆ ทำงานอย่างไรสามารถช่วยปรับแต่งการวิศวกรรมสายเซลล์ได้ แต่ละยีนมีกลไกเฉพาะตัว พฤติกรรมระหว่างการแยกตัว และการประยุกต์ใช้ที่เป็นไปได้ ตารางด้านล่างสรุปความแตกต่างเหล่านี้ ทำให้ง่ายต่อการตัดสินใจว่ายีนใด - หรือการรวมกันของยีน - อาจทำงานได้ดีที่สุดสำหรับความต้องการของคุณ
| ยีน | กลไกหลัก | ความเสถียรของการแสดงออก | ผลกระทบต่อความมีชีวิตที่รายงาน | ความเข้ากันได้กับการแก้ไข |
|---|---|---|---|---|
| BCL-2 | บล็อก BAX/BAK ที่ส่งเสริมการตายของเซลล์และรับประกันการอยู่รอดของเซลล์ที่ยังไม่แตกต่าง[2] | ค่อนข้างคงที่ในระหว่างการแยกแยะ[2] | จำเป็นสำหรับการรักษากลุ่มเซลล์ต้นกำเนิดเริ่มต้น[2] | ความเข้ากันได้สูงกับเครื่องมือแก้ไข |
| BCL-xL | ยับยั้ง caspase-3 ที่ถูกตัด; รักษาความสมบูรณ์ของเยื่อหุ้มไมโตคอนเดรียและการเผาผลาญ[2] | เพิ่มขึ้นตั้งแต่วันที่ 7 ของการแยกแยะ[2] | จำเป็นสำหรับการสนับสนุน progenitors ที่แตกต่าง; การยับยั้งของมันเพิ่มการตายของเซลล์ [2] | ความเข้ากันได้สูงกับเครื่องมือแก้ไข |
| MCL-1 | ปรับสัญญาณ pro-apoptotic เป็นส่วนหนึ่งของครอบครัว BCL-2 [2] | การแสดงออกคงที่ในระหว่างการระบุสายพันธุ์ [2] | ให้ประโยชน์การอยู่รอดที่กว้างขวางแต่ขาดผลกระทบเฉพาะขั้นตอนเช่น BCL-xL [2] | ความเข้ากันได้สูงกับเครื่องมือแก้ไข |
| BIRC5 (Survivin) | บล็อก caspase-3 และ caspase-7; ช่วยในการแยกโครโมโซมระหว่างไมโทซิส | สูงในเซลล์ที่เพิ่มจำนวน; ลดลงเมื่อมีการแยกแยะขั้นสุดท้าย | สนับสนุนการอยู่รอดและการเพิ่มจำนวนในเซลล์ที่แบ่งตัวอย่างรวดเร็ว | เข้ากันได้กับทั้ง shRNA knockdown และ CRISPR editing |
| XIAP | ยับยั้ง caspases หลายตัว ให้การป้องกันการตายของเซลล์อย่างกว้างขวาง | โดยทั่วไปมีความเสถียรในสภาวะต่างๆ | มีประสิทธิภาพเป็นพิเศษภายใต้ความเครียด เช่น สภาวะ bioreactor ความหนาแน่นสูง | มีความเข้ากันได้สูงกับเครื่องมือแก้ไข |
BCL-xL โดดเด่นในบทบาทคู่ในการส่งเสริมการอยู่รอดของเซลล์และสนับสนุนกิจกรรมการเผาผลาญ โดยเฉพาะในช่วงการแยกแยะที่สำคัญเมื่อโปรตีน pro-apoptotic เช่น BAK ลดลงตามธรรมชาติBCL-2, ในทางกลับกัน เหมาะสำหรับการรักษาเซลล์ที่ยังไม่แยกแยะ ในขณะที่ XIAP ให้การปกป้องที่กว้างขวาง โดยเฉพาะในสภาพแวดล้อมที่มีความเครียด เช่น วัฒนธรรมที่มีความหนาแน่นสูง
ไม่มียีนใดที่ทำงานได้ดีที่สุดในทุกสถานการณ์ ตัวอย่างเช่น BIRC5 มีประโยชน์อย่างยิ่งในสถานการณ์ที่ต้องการการแบ่งเซลล์อย่างรวดเร็ว ในทางปฏิบัติ การรวมยีนสองตัวหรือมากกว่ามักจะให้การปกป้องที่มีประสิทธิภาพที่สุด โดยตอบสนองต่อการกระตุ้นการตายของเซลล์หลายประเภทพร้อมกัน
ผลการวิจัยเหล่านี้เป็นพื้นฐานสำหรับการรวมยีนเหล่านี้เข้ากับกลยุทธ์การวิศวกรรมสายเซลล์สำหรับการผลิตเนื้อสัตว์เพาะเลี้ยง ซึ่งรวมถึงการเลือก วัตถุดิบเนื้อสัตว์เพาะเลี้ยง ที่เหมาะสมเพื่อให้แน่ใจว่าสามารถขยายขนาดได้
การใช้ยีนเหล่านี้ในการวิศวกรรมสายเซลล์เนื้อสัตว์เพาะเลี้ยง
เพื่อปรับปรุงความมีชีวิตของเซลล์ในการผลิตเนื้อสัตว์เพาะเลี้ยง การรวมยีนสำคัญอย่างมีกลยุทธ์เป็นสิ่งสำคัญการระบุยีนต้านการตายของเซลล์เพียงอย่างเดียวนั้นไม่เพียงพอ - การนำยีนเหล่านี้ไปใช้ในสายเซลล์อย่างมีประสิทธิภาพคือสิ่งที่ทำให้เกิดความแตกต่าง สองกลยุทธ์หลักที่ใช้กันทั่วไปคือ: การแสดงออกมากเกินไปของยีนต้านการตายของเซลล์ เช่น BCL-2, BCL-xL, และ MCL-1 เพื่อเพิ่มการอยู่รอดของเซลล์ หรือ การลบยีนที่ส่งเสริมการตายของเซลล์ เช่น BAX, BAK, และ BOK เพื่อกำจัดตัวขับเคลื่อนการตายของเซลล์ การรวมวิธีการเหล่านี้มักส่งผลให้สายเซลล์เหมาะสมยิ่งขึ้นสำหรับการผลิตขนาดใหญ่ [1].
เครื่องมือแก้ไขยีนสมัยใหม่เช่น CRISPR/Cas9 ช่วยให้สามารถแก้ไขพร้อมกันได้ เช่น การลบ Bak1, Bax, และ Bok ในขั้นตอนเดียว ทางเลือกอื่นเช่น ZFNs หรือ การรบกวน RNA สามารถใช้เพื่อลดกิจกรรมของ caspases ชั่วคราว (e.g . caspases-3, -7, -8, and -9). สำหรับกลยุทธ์การแสดงออกเกิน, โปรโมเตอร์สังเคราะห์ช่วยให้มั่นใจถึงระดับการแสดงออกที่สม่ำเสมอและสูงของยีนเช่น BCL-2 ในระหว่างการขยายขนาด, ซึ่งมีความสำคัญต่อการรักษาประสิทธิภาพของเซลล์ใน ระบบการเพาะเลี้ยงแบบ fed-batch หรือแบบต่อเนื่อง . วิธีการรวมกันเหล่านี้เสริมสร้างการพัฒนาเซลล์ไลน์สำหรับการประยุกต์ใช้เนื้อสัตว์ที่เพาะเลี้ยง
การดัดแปลงพันธุกรรมดังกล่าวมีผลกระทบโดยตรงต่อการปรับปรุง ความเข้มข้นของเซลล์ที่มีชีวิตรวม (IVCC), ซึ่งเป็นตัวชี้วัดสำคัญในการผลิตเนื้อสัตว์ที่เพาะเลี้ยง การตายของเซลล์เด่นชัดที่สุดในช่วงห้าวันแรกของการแยกแยะ, ทำให้การแทรกแซงในช่วงต้นด้วยยีนเช่น BCL-2 หรือ BCL-xL เป็นสิ่งจำเป็น งานวิจัยที่ตีพิมพ์ใน Cell Death & Disease เน้นว่า BCL-xL การแสดงออกเพิ่มขึ้นเมื่อเซลล์แยกแยะ, บ่งชี้ว่า progenitors ที่มีความเป็นผู้ใหญ่มากขึ้นพึ่งพาบทบาทการป้องกันของมันอย่างมาก [2] . โดยการตรวจสอบระดับการแสดงออกของยีนตระกูล BCL-2 ตลอดช่วงการเจริญเติบโต การแทรกแซงสามารถกำหนดเวลาได้อย่างแม่นยำเพื่อให้ได้ผลสูงสุด.
"โดยการสร้างสายเซลล์ที่มีความเสถียรซึ่งแสดงออกมากเกินไปของยีนต้านการตายของเซลล์หรือยีนที่ลดการตายของเซลล์ ผลผลิตสุดท้ายสามารถเพิ่มขึ้นได้เนื่องจากเซลล์มีความทนทานต่อความเครียดจากสิ่งแวดล้อมมากขึ้น" - Gyun Min Lee et al. [1]
สำหรับการผลิตที่ใช้ไบโอรีแอคเตอร์ เซลล์ต้องได้รับการออกแบบให้ทนต่อ ความเครียดจากความเข้มข้นสูง และ การขาดสารอาหาร. ก่อนการขยายขนาด จำเป็นต้องตรวจสอบการแก้ไขทางพันธุกรรมโดยใช้เครื่องมือเช่น Western blot หรือ FACS สำหรับนักวิจัยที่ต้องการสายเซลล์เฉพาะหรือวัสดุทางพันธุกรรมที่ปรับแต่งสำหรับสภาพแวดล้อมไบโอรีแอคเตอร์ความหนาแน่นสูง แพลตฟอร์มเช่น
บทสรุป
การเลือกยีนต้านการตายของเซลล์สำหรับสายเซลล์เนื้อสัตว์เพาะเลี้ยงต้องใช้วิธีการที่ปรับแต่งเฉพาะ ยีนเช่น BCL-2 , BCL-xL, และ MCL-1 แต่ละตัวมีบทบาทเฉพาะในการปกป้องเซลล์ แต่ความสำเร็จของพวกมันขึ้นอยู่กับปัจจัยต่างๆ เช่น ประเภทของเซลล์ ระยะการพัฒนา และความเครียดเฉพาะที่พบในระหว่างการผลิต ดังที่เน้นในงานวิจัย:
"ความสมดุลระหว่างสมาชิกที่ต้านการตายของเซลล์และสมาชิกที่ส่งเสริมการตายของเซลล์จะเป็นตัวกำหนดว่าเซลล์จะมีชีวิตอยู่หรือตาย" [2]
นอกเหนือจากการอยู่รอด การวิศวกรรมต้านการตายของเซลล์ยังช่วยรักษาการทำงานของเมตาบอลิซึมอีกด้วย ตัวอย่างเช่น โปรตีนเช่น BCL-xL มีความสัมพันธ์ใกล้ชิดกับการรักษากระบวนการไกลโคไลซิสและการฟอสโฟรีเลชันออกซิเดทีฟ อย่างไรก็ตาม การแทรกแซงที่ดำเนินการไม่ดีอาจทำให้กระบวนการที่สำคัญเหล่านี้เสียหายได้ [2]. การรับรองว่าเซลล์ไลน์ที่ถูกออกแบบยังคงรักษาอัตลักษณ์และกิจกรรมเมตาบอลิกที่ตั้งใจไว้ตลอดการผลิตเป็นขั้นตอนที่สำคัญ แม้ว่าบางครั้งจะถูกมองข้าม ข้อมูลเชิงลึกเหล่านี้กำลังกำหนดอนาคตของการออกแบบเซลล์ไลน์
วิธีการหลายยีนใหม่กำลังเกิดขึ้น ซึ่งรวมการแสดงออกเกินของยีนป้องกันกับการเคาะออกด้วย CRISPR ของยีนโปร-อะพอพโทติก เช่น BAX , BAK1, และ BOK เพื่อสร้างเซลล์ไลน์ที่แข็งแกร่งขึ้นสำหรับการใช้งานในอุตสาหกรรม [1]. เครื่องมือสำหรับการวิเคราะห์เมตาบอลิก เช่น การทดสอบพลังงานชีวภาพ กำลังกลายเป็นสิ่งจำเป็นเพื่อยืนยันว่าการดัดแปลงทางพันธุกรรมเหล่านี้ช่วยเพิ่มประสิทธิภาพโดยรวมของเซลล์ สำหรับนักวิจัยที่จัดหา เซลล์ไลน์จากสุกร, วัสดุพันธุกรรม หรืออุปกรณ์ไบโอรีแอคเตอร์
คำถามที่พบบ่อย
ฉันควรเริ่มต้นด้วยยีนต้านการตายแบบใดสำหรับสายเซลล์ของฉัน?
BCL-2 มักถูกแนะนำให้เป็นจุดเริ่มต้นเมื่อทำงานกับสายเซลล์ ยีนต้านการตายที่ได้รับการวิจัยอย่างดีนี้เป็นที่รู้จักในด้านความสามารถในการปรับปรุงการอยู่รอดของเซลล์ ทำให้เป็นตัวเลือกยอดนิยมในการวิจัยเนื้อสัตว์ที่เพาะเลี้ยง ฟังก์ชันของมันในการสนับสนุนความมีชีวิตของเซลล์ทำให้เป็นตัวเลือกที่ใช้งานได้จริงสำหรับการทดลองในระยะเริ่มต้น
การแสดงออกของยีนต้านการตายแบบเกินหรือการปิดยีนที่ส่งเสริมการตายแบบใดดีกว่ากัน?
ในการผลิตเนื้อสัตว์ที่เพาะเลี้ยง การเพิ่มการแสดงออกของยีนต้านการตาย เช่น สมาชิกของตระกูล BCL-2 อย่าง BCL-xL, มักจะให้ผลลัพธ์ที่ดีกว่าการปิดยีนที่ส่งเสริมการตาย กลยุทธ์นี้สนับสนุนทั้งการอยู่รอดและการเพิ่มจำนวนของเซลล์ ซึ่งเป็นปัจจัยสำคัญสำหรับการขยายการผลิต ในขณะที่ยังคงรักษาระบบการควบคุมตามธรรมชาติของเซลล์ไว้
โดยการเพิ่มกิจกรรมของยีนต้านการตายของเซลล์ เซลล์จะมีความต้านทานต่อการตายของเซลล์มากขึ้น โดยเฉพาะในสภาวะที่เครียด ซึ่งทำให้เป็นวิธีที่น่าเชื่อถือและปลอดภัยมากขึ้นในการรักษาความมีชีวิตของเซลล์ในระหว่างกระบวนการเพาะเลี้ยง
ฉันจะยืนยันได้อย่างไรว่าการแก้ไขยีนต้านการตายของเซลล์ช่วยปรับปรุง IVCC ในเครื่องปฏิกรณ์ชีวภาพของฉัน?
เพื่อพิจารณาว่าการแก้ไขยีนต้านการตายของเซลล์ช่วยเพิ่ม in vitro ความมีชีวิตและการเพิ่มจำนวนของเซลล์ (IVCC) หรือไม่ คุณจะต้องใช้วิธีการที่เป็นระบบ:
- ประเมินอัตราการมีชีวิตและการเพิ่มจำนวน: ใช้วิธีการเช่นการนับเซลล์หรือโฟลไซโตเมทรีเพื่อวัดอัตราเหล่านี้ทั้งก่อนและหลังการแก้ไขยีน
- ตรวจสอบการแสดงออกของยีน: เทคนิคเช่น qPCR หรือ Western blotting สามารถยืนยันการแสดงออกของยีนที่ต้องการได้สำเร็จ
- ตรวจสอบตัวบ่งชี้การตายของเซลล์: ตรวจสอบตัวบ่งชี้เช่นกิจกรรมของ caspase เพื่อให้แน่ใจว่าการแก้ไขลดการตายของเซลล์ได้อย่างมีประสิทธิภาพ
สำหรับการประเมินผลที่สมบูรณ์ จำเป็นต้องทดสอบความเสถียรและการเพิ่มจำนวนของเซลล์ที่แก้ไขในระยะยาวในเครื่องปฏิกรณ์ชีวภาพ เพื่อให้แน่ใจว่าการปรับปรุงยังคงอยู่ตลอดหลายรอบการเพาะเลี้ยง










