

培養肉は、従来の肉の味と食感を提供しながら、同じ健康上の懸念を伴わない食品生産の考え方を変革しています。重要な焦点は、より健康的にするために脂肪組成を改善することです。
知っておくべきことは次のとおりです:
- 心血管リスクに関連する飽和脂肪よりも、一価不飽和脂肪酸やオメガ3脂肪酸のような健康的な脂肪が優先されています。
- 経路工学は、細胞レベルでの脂肪生産に影響を与えるために代謝および遺伝的技術を使用します。
- 方法には以下が含まれます:
- CRISPR-Cas9 遺伝子編集 により飽和脂肪の生産を減少させます。
- 酵素の過剰発現 ( e.g. , ステアロイル-CoAデサチュラーゼ) により一価不飽和脂肪を増加させます。
- 成長培地の補充により、遺伝子改変なしでオメガ3含有量を増加させます。
- 課題には、生産の拡大と栄養価を向上させながら風味を維持することが含まれます。
このアプローチは、培養肉の生産者がより健康的で現代の食事ニーズに適した製品を作るのに役立っています。
培養肉と持続可能な細胞農業のための細胞株の工学 #culturedmeat
sbb-itb-ffee270
培養肉における脂肪酸合成の仕組み
脂肪酸合成は、特に飽和脂肪レベルを低下させることを目指す際に、培養肉の脂肪含有量を形成する上で重要な役割を果たします。細胞レベルで脂肪の組成を管理することにより、科学者は結果として得られる肉が飽和脂肪、モノ不飽和脂肪、または多価不飽和脂肪を含むかどうかに影響を与えることができます。これは、脂肪プロファイルに寄与する3つの相互接続された代謝経路を通じて達成されます。それらを分解してみましょう。
脂肪酸合成酵素経路
このプロセスは脂肪酸合成酵素(FAS)経路, から始まり、飽和脂肪の生成を担っています。この経路の中心には、細胞質での脂肪酸合成の最初のステップを触媒する酵素アセチル-CoAカルボキシラーゼ(ACC)があります。この酵素は、培養肉の生産において重要な成熟脂肪細胞のマーカーとしても機能します[5].
興味深いことに、細胞が脂肪酸を生成する方法は種によって異なることがあります。例えば、ウシの細胞は酢酸を使用する傾向がありますが、人間の細胞は脂肪酸合成においてグルコースにより依存しています[1]. これらの違いは、特定のニーズに合わせて経路を調整することの重要性を強調しています。
デサチュラーゼ酵素と一価不飽和脂肪
飽和脂肪が合成されると、デサチュラーゼ酵素が介入し、それらを一価不飽和脂肪酸(MUFAs)に変換します。これらはより健康的と考えられています。例えば、これらの酵素はパルミチン酸やステアリン酸のような飽和脂肪をオレイン酸(C18:1)に変換することができます。オレイン酸はオリーブオイルの健康効果と一般的に関連付けられています[5] .
線維脂肪前駆細胞から得られる培養脂肪は、従来の牛脂と比較してオレイン酸のレベルが高く、パルミチン酸のレベルが低い傾向があります[5]. この組成の変化は培養条件によってさらに影響を受ける可能性があります。例えば、無血清培地の処方を使用することで、従来の血清含有培地と比較して、牛脂肪幹細胞におけるトリグリセリドの蓄積が66%増加することが示されています[1] .
MUFAsを超えて、栄養プロファイルを改善するために多価不飽和脂肪の調整が行われます。
多価不飽和脂肪酸経路
多価不飽和脂肪酸(PUFAs), オメガ3やオメガ6脂肪酸などは、培養肉の栄養価を高める方法を提供します。リノール酸を含むこれらの必須脂肪は人体で生成されず、食物から摂取する必要があります。
しかし、培養肉は通常の肉よりもPUFAレベルが低いことが多いです [5]. これに対処するために、研究者はPPARγ、Gpd1、FABP4などのトリグリセリド合成に関与する遺伝子の発現に注目しています [6][1]. これらの経路をターゲットにすることで、PUFAの含有量を増やし、肉をより栄養価の高いものにすることができます。
さらに、PUFAの組成はメディア補給. によって微調整することができます。 成長培地に特定の脂質を加えることで、科学者は自然の動物組織の脂肪プロファイルを再現したり、遺伝子組み換えなしで栄養価を高めた製品を作成したりすることができます。[3].
| 経路/酵素 | 主な機能 | 脂肪組成への影響 |
|---|---|---|
| 脂肪酸合成酵素 (FAS) | 長鎖飽和脂肪酸を生成 | 飽和脂肪含有量を増加させる (e.g. , パルミチン酸) |
| アセチル-CoAカルボキシラーゼ (ACC) | 脂肪酸合成の律速段階 | 全体的な脂質蓄積レベルに不可欠 |
| デサチュラーゼ酵素 | 飽和結合を二重結合に変換 | オレイン酸のような一価不飽和脂肪酸 (MUFA) を増加 |
| PPARγシグナリング | 脂肪生成遺伝子の発現を調節 | 脂質貯蔵の成熟と体積を制御 |
遺伝子および代謝工学による脂肪プロファイルの改善方法
脂肪酸がどのように合成されるかを理解することで、遺伝子および代謝工学を使用して培養肉の脂肪組成を改善する機会が開かれました。これらのアプローチは、飽和脂肪酸のレベルを下げると同時に、より健康的な脂肪酸を増やし、現代の食事の好みに合わせた栄養プロファイルを調整することを目的としています。
CRISPR-Cas9による標的遺伝子編集
CRISPR-Cas9技術は、科学者がDNAを正確に編集することで脂肪の組成を調整することを可能にします。この方法は、他の種からの外来DNAを導入することなく、飽和脂肪を生成する責任のある遺伝子を標的にして無効化することができます [7].
"CRISPRは遺伝子編集ツールであり、分子のハサミとして考えることができます。そして、そのハサミをゲノムの特定の位置に導き、DNAに正確な切り込みを入れることができます。" - Dawn Cayabyab, Ph.D. 学生、UC Davis [7]
2025年6月、南京農業大学, のShijie Ding、Chunbao Li、Guanghong Zhouを含む研究者たちは、培養肉生産におけるCRISPR/Cas9の可能性を実証しました。豚の衛星細胞においてCDKN2A遺伝子をノックアウトすることで、細胞の老化に対処し、筋肉前駆細胞の再生可能な供給源を作り出しました。これらの改変された細胞は、18回以上の継代にわたって90%以上の生存率で安定した成長を維持しました。3D食用足場, を使用して、チームは肉のような構造物を開発することに成功し、CRISPRによるスケーラビリティと遺伝子最適化の可能性を示しました [8].
この精密な編集ツールにより、研究者は飽和脂肪レベルが低い細胞を直接選択することが可能になります。さらに、酵素発現を修正することで、脂肪プロファイルを改善する別の方法が提供されます。
ステアロイル-CoAデサチュラーゼ(SCD)の過剰発現
脂肪組成を改善する別の方法は、ステアロイル-CoAデサチュラーゼ(SCD)の活性を高めることです。この酵素は、ステアリン酸のような飽和脂肪酸を、オレイン酸のような一価不飽和脂肪酸に変換します[2]. SCDの発現を増加させることで、脂質プロファイルを一価不飽和脂肪にシフトさせることができ、これらは健康的と広く認識されています。
このアプローチは、特に血清フリーの培地システムと組み合わせると効果的です。研究によると、これらのシステムは従来の血清含有培地と比較してトリグリセリドの蓄積を66%増加させることができます[9]. その結果、栄養の推奨に沿った、より健康的な脂肪組成の培養肉が得られます。
オメガ3強化のための培養基補充
遺伝子改変を超えて、培養基を調整することで脂肪酸プロファイルをさらに向上させることができます。例えば、リノレン酸のような不飽和脂肪酸を培養基に補充することで、細胞の生存率を損なうことなく細胞内脂質レベルを向上させます[4].
慎重に設計された脂肪酸の混合物は、天然の牛肉の脂肪プロファイルを再現することができます。この方法は、培養基中の総脂質濃度を400 µMまでサポートします - パルミチン酸のような飽和脂肪の毒性閾値をはるかに超えています。リノレン酸のような不飽和脂肪酸は細胞によく耐えられ、非毒性レベルは200 µMまで達しますが、パルミチン酸の毒性は約40 µMです[4].
"n-3多価不飽和脂肪酸(PUFAs)などの健康促進脂肪酸の組み込みは、これらの製品の栄養価を高めるための潜在的な戦略を表しています。" - Waris Mehmood et al., Aarhus University [4]
3D培養システム用のバイオマテリアル, (スフェロイドなど)と組み合わせると、培地の補充はさらに影響力を増します。この組み合わせは、2D単層培養と比較してトリグリセリドの蓄積を最大34%増加させることが示されています[9]. ただし、最終製品に「魚のような」風味を生じさせないように、オメガ3のレベルを慎重に管理する必要があります[4].
異なる経路工学アプローチの比較
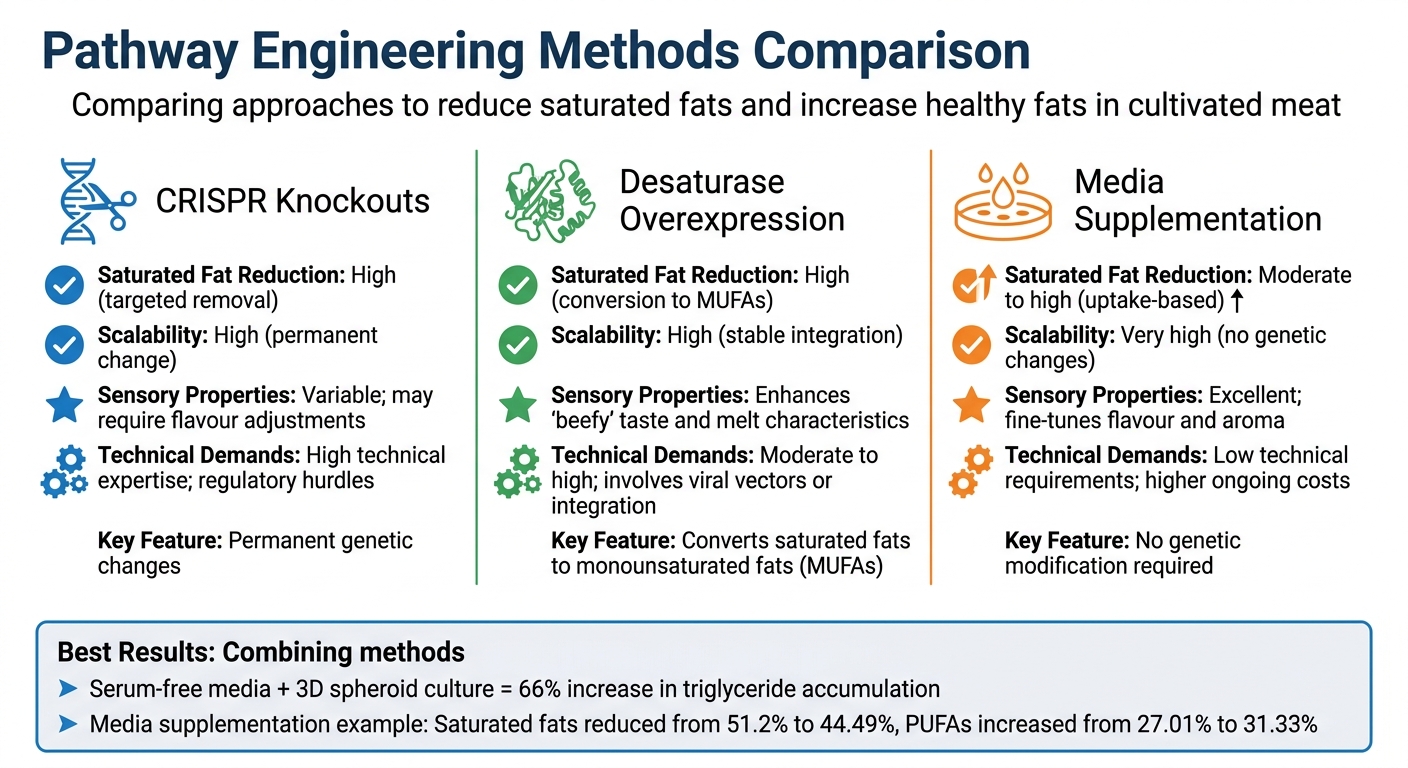
培養肉における健康的な脂肪プロファイルのための経路工学手法
このセクションでは、以前に議論された技術を基に、さまざまな経路工学手法の強みとトレードオフを掘り下げます。各アプローチは培養肉の脂肪プロファイルを改善するための独自の利点を提供し、選択は主に生産目標、技術的リソース、および栄養目標に依存します。
まずはCRISPRベースの遺伝子ノックアウト. から始めましょう。これらは恒久的な遺伝的変化を生み出し、一度実施されると非常にスケーラブルです。しかし、厳しい規制要件や高度な技術的専門知識の必要性などの課題も伴います。一方で、デサチュラーゼの過剰発現, 、特にSCD酵素を含むものはバランスを取ります。この方法は、飽和脂肪をより健康的な一価不飽和脂肪酸(MUFAs)に継続的に変換する安定した細胞株を確立し、継続的な外部入力の必要性を排除します。
次に、メディア補充, があります。これはそのシンプルさと迅速な適用で際立っています。2026年の研究ではその効果が実証されました:オリーブオイルと大豆レシチンを脂肪生成誘導剤として使用することで、培養された豚肉の飽和脂肪酸を51.2%から44.49%に減少させ、同時に多価不飽和脂肪酸を27.01%から31.33%に増加させました[10]. シンプルで効果的ではありますが、メディア補充には継続的なコストが伴い、慎重な財務計画が必要です。高度な3Dスフェロイドシステム, と組み合わせることで、この方法はトリグリセリドの蓄積をさらに促進することができます。
メソッド比較表
| メソッド | 飽和脂肪削減 | スケーラビリティ | 感覚特性 | 技術的要求 |
|---|---|---|---|---|
| CRISPRノックアウト | 高い(ターゲット除去) | 高い(永久的な変化) | 変動する; 味の調整が必要な場合あり | 高い技術的専門知識; 規制上のハードル |
| デサチュラーゼ過剰発現 | 高い(MUFAへの変換) | 高い(安定した統合) | 「ビーフィー」な味と溶解特性を強化 | 中程度から高い; ウイルスベクターまたは統合を含む |
| メディア補給 | 中程度から高い(取り込みベース) | 非常に高い(遺伝的変化なし) | E |
低い技術要件; 継続的なコストが高い |
この比較から、最良の結果はしばしば方法を組み合わせることで得られることが明らかです。例えば、無血清培地と3Dスフェロイド培養を組み合わせることで、従来の技術と比較してそれぞれ66%および34%のトリグリセリド蓄積の増加が示されています[9]. この層状アプローチにより、研究者は遺伝的および環境的要因の両方を洗練し、消費者に訴求し健康基準を満たす最適化された脂肪プロファイルを持つ培養肉を作成できます。
経路工学のための機器と材料
培養肉における健康的な脂肪プロファイルを作成するには、一般的な供給業者からは通常入手できない特殊なツールと生物材料が必要です。この分野は大きな成長を遂げており、2025年までに140社以上が27億ポンド以上を投資すると予測されています[12].
この作業の主要なリソースには以下が含まれます:
- 細胞株: 例として、豚脂肪幹細胞, 牛筋衛星細胞、および水牛脂肪細胞[11].
- 無血清培地の配合: スケーラブルな生産に不可欠[4].
- 脂肪酸: オレイン酸、リノール酸、リノレン酸、ステアリン酸、パルミチン酸など、脂肪プロファイルを微調整するため[4].
- バイオリアクター: 撹拌槽、エアリフト、充填床、または灌流システムなどのオプション[12].
- 3Dスフェロイド培養システム: 細胞の成熟を促進するために使用[12].
- 分析ツール: RT-qPCR、フローサイトメトリー、Agilent BioTek Cytation 5のような高解像度イメージングシステムを含む[4].
機器と材料の検索 Cellbase
培養肉の研究者にとって、これらの専門的な材料の調達は
一次細胞株や成長因子のような敏感な生物材料は、輸送中の生存性を維持するためにコールドチェーン物流で取り扱われます。さらに、研究者は「Cell Ag Experts」に相談して、特にラボベースの機器から製造規模のバイオリアクターへのスケールアップ時に技術的なアドバイスを受けることができます。プラットフォームの「何でも聞いてください」機能は、この移行期間中に貴重なサポートを提供します[12]. これらのツールとサービスは、調達を迅速化し、複数の一般的なサプライヤーをナビゲートする際のリスクを軽減します。
経路工学ワークフローの設定
効率的な経路工学ワークフローを確立するには、材料の互換性とプロセス制御に細心の注意を払う必要があります。例えば、足場は37°Cの培養条件、滅菌、および調理プロセスに耐える必要があります [12]. グルコース、乳酸、アンモニウムレベルのリアルタイムセンサーは、正確な代謝制御を維持するために重要です[12].
結論と今後の方向性
経路工学は、培養肉における脂肪プロファイルの微調整においてエキサイティングな可能性を開いています。無血清培地の最適化や高度な3D培養システムのような技術を活用することで、研究者は従来の家畜農業では再現できない栄養精度を達成できるようになりました。
最も有望なブレークスルーのいくつかは、複数の戦略を組み合わせることから生まれています。例えば、FaTTy豚細胞株は、遺伝子編集を必要とせずにMUFAプロファイルを改善する方法を示しています[2]. 同様に、オーフス大学のマーティン・クローイヤー・ラスムッセンは、2025年12月に、400 µMで慎重にバランスを取った脂肪酸ミックスに分化したウシ衛星細胞を曝露することで、細胞の生存率を維持しながら最も高い脂質滴の蓄積が得られることを示しました[4] .
しかし、特に生産のスケールアップに関しては課題が残っています。3D培養では、酸素や栄養素の勾配のような物質輸送の制限が、密集した組織の中心部での細胞死を引き起こす可能性があります[1]. 実用的な解決策は、細胞拡張のための高密度バイオリアクターを使用し、その後に専門的な3D分化段階を行う二段階のバイオプロセシングにあります[1]. さらに、オメガ3脂肪酸で製品を強化することは有望ですが、高濃度で魚のようなオフフレーバーのリスクを避けるために慎重な調整が必要です[4].
血清を含まない培地への移行は、進展のもう一つの重要な領域です。倫理的および環境的な利点を超えて、血清を含まない配合は、細胞増殖と脂質蓄積の両方を向上させるのに効果的であることが証明されています[1]. これらの進歩は、培養肉の生産方法を変革しています。
最終的に、この分野での成功は、特定の製品目標を達成するために、細胞タイプ、培養システム、および培地配合の適切な組み合わせを選択することに依存しています。目標が飽和脂肪酸レベルを下げること、オメガ3の含有量を増やすこと、またはリアルな霜降りを作り出すことであれ、ここで概説されている経路工学戦略は、栄養的に最適化された次世代の培養肉を作り出すための強固な基盤を提供します。これらの開発は、培養肉産業にとってより健康的で商業的に実現可能な未来を示しています。
よくある質問
培養肉の飽和脂肪を最も効果的に減少させる経路工学の方法はどれですか?
培養肉の飽和脂肪を減少させる効果的な方法の一つは、無血清培地. を使用することです。この技術は筋衛星細胞における脂質蓄積を微調整し、脂肪酸プロファイルをより制御できるようにします。その結果、最終製品の飽和脂肪含有量を減少させるのに役立ちます。これらの進歩は、培養肉のより健康的な脂肪プロファイルを作り出す上で重要な役割を果たします。
細胞のDNAを変えずにオメガ3レベルをどうやって増やせますか?
培養肉のオメガ3レベルを増やすために、培地に微細藻類由来のオメガ3脂肪酸を追加することができます。この方法は、細胞のDNAを変えることなく、その栄養プロファイルを改善します。
健康的な脂肪プロファイルは、培養肉の風味、香り、または口当たりに影響を与えますか?
健康的な脂肪プロファイルは、培養肉の風味、香り、そして食感に影響を与えると予想されます。脂肪はこれらの感覚的な特性を形作る重要な要素です。良いニュースは、培養脂肪がその化学的構成と感覚的属性の両方で伝統的な脂肪を非常に近く模倣できることを既に示していることです。これは、健康上の利益を提供しつつ、人々が愛する味を維持するバランスを取ることを意味します。










