

CRISPRは、工業用バイオリアクターにおける細胞ストレスという主要な課題に対処することで、培養肉の生産を変革しています。このツールは、過酷な条件下での細胞の生存率を向上させ、増殖を延ばし、老化を減少させるための正確な遺伝子編集を可能にします。例えば、 TP53や PTENのような遺伝子をノックアウトすることで、 一次細胞系と不死化細胞系の培養期間を100日から200日に延ばし、30日で細胞の豊富さを1,000倍に増加させました。しかし、これらの改変は分化に影響を与える可能性があるため、慎重な最適化が必要です。
記事からの重要な洞察には以下が含まれます:
- バイオリアクターにおけるストレス要因: せん断力、栄養不均衡、酸化ストレスが細胞の生存率を低下させます。
- CRISPR戦略: 遺伝子ノックアウト(TP53, PTEN)および活性化(HIF1A)は、特定のストレス応答をターゲットにしています。
- 検証: 編集された細胞は、性能と分化の可能性を確保するために、ゲノム、プロテオーム、および機能テストを受けます。
-
スケールアップ: バイオリアクター条件への移行には、最適化された培地と機器が関与しており、
Cellbase のようなプラットフォームがカスタマイズされたリソースを提供します。
CRISPRの精度はストレス耐性のある細胞株の開発を可能にしますが、成長と分化のバランスを取ることは、スケーラブルな培養肉の生産において依然として重要です。
遺伝子設計のためのバイオリアクターストレスプロファイルのマッピング
主要なバイオリアクターストレス要因の特定
CRISPR編集を開始する前に、遺伝子設計を導くためにバイオリアクターストレスプロファイルをマッピングすることが重要です。バイオリアクター内のストレス要因は特定の細胞応答を引き起こし、適切な遺伝子ターゲットを選択するために十分に理解する必要があります。
機械的および流体力学的ストレスは最も差し迫った課題の一つです。撹拌槽型バイオリアクターは、細胞膜を損傷し、細胞シグナル伝達経路に干渉する剪断力を生み出します [5][2]. 栄養および代謝ストレスも大きな役割を果たし、しばしば不均一な栄養素の取り込みに起因します。3D足場における栄養素の勾配とアンモニアの蓄積が代謝負荷に寄与します [3][5][6]. さらに、pHの変動や高温は細胞増殖率を低下させ、細胞を早期分化に向かわせることさえあります[3][2].
酸化ストレス、ミトコンドリアストレス、ERストレスを含む他のストレッサーも、細胞の生存率にさらなる挑戦をもたらします。酸化ストレスは、特に血清不使用培地への移行時に深刻になります。天然の抗酸化物質がないため、細胞は活性酸素種に対してより脆弱になります [4]. 細胞レベルでは、ミトコンドリアストレスや小胞体(ER)ストレス が、バイオプロセス条件が最適範囲から逸脱したときに発生します [6]. UCSFの神経変性疾患研究所のXiaoyan Guoはこの動態を強調しています:
"異なる生理的および環境的ストレスの存在下で、細胞は迅速にストレス応答を開始し、細胞の恒常性を再確立します。" [6]
これらのストレス要因を積極的にマッピングすることで、問題が発生したときに反応するのではなく、研究者は正確な遺伝子工学の目標を定義することができます。この体系的なアプローチは、CRISPR戦略がストレス耐性のある細胞株の開発を効果的にターゲットにすることを保証します。
オミクスデータを使用してストレス応答遺伝子を見つける
ストレス環境を特徴付けた後、次のステップはこれらの条件に応答する遺伝子を特定することです。トランスクリプトミクス(RNA-seq)やプロテオミクスのようなツールは、細胞が健康な初期継代状態からストレスのかかった後期継代状態に移行する際の遺伝子発現とタンパク質量の変化を追跡するのに非常に貴重です [1] [6]. しかし、これらの方法は下流の効果を捉える一方で、これらの変化を引き起こす上流の調節因子を特定することにはしばしば失敗します[6].
プール型CRISPRノックアウトスクリーニングはこのギャップを埋めます。大規模な細胞集団において、数千の遺伝子を体系的に破壊することにより、これらのスクリーニングは、ストレス下で成長の優位性をもたらす遺伝子の変異を明らかにし、重要な調節ハブを発見します。例えば、TP53やPTEN のような遺伝子を標的にすることで、長期間の培養ストレスによって引き起こされる分子老化のシグネチャーを逆転させることが示されています。これにより、後期継代の細胞が初期継代の野生型細胞に類似したトランスクリプトームプロファイルを維持することが可能になります。
階層的クラスタリングを使用することで、研究者は時間の経過に伴う発現変化に基づいて遺伝子をグループ化し、細胞周期の進行やタンパク質合成のようなプロセスに関連するモジュールを分離することができます。これらのプロセスは通常、バイオリアクターによって誘発される老化が進行するにつれて低下します。経路エンリッチメント分析(gprofiler2のようなツールを使用)と組み合わせることで、これらのモジュールはTGFβシグナル伝達や軟骨分化などの特定の生物学的経路に関連付けることができ、細胞の拡大を積極的に制限する可能性があります [1] .
以下の表は、包括的なストレスマップを構築するための各手法の貢献を示しています。
| 手法 | 主な用途 | 主要な出力 |
|---|---|---|
| トランスクリプトミクス (RNA-seq) | mRNA発現変化の測定 | ストレスを受けた細胞と受けていない細胞間の差次的に発現した遺伝子 (DEGs)[1] |
| プロテオミクス | タンパク質の存在量の測定 | 特定のストレッサーにマッピングされた翻訳出力[6] |
| プール型CRISPRスクリーン | 機能的遺伝子の摂動 | 上流の調節ハブとフィットネスに重要な遺伝子[1][6] |
| PCA & 階層クラスタリング | データの視覚化とグループ化 | 細胞状態の変化と共調整されたストレス応答経路[1] |
sbb-itb-ffee270
CRISPRを用いた細胞株エンジニアリング-Cas9 - 成功を最大化するためのヒントとコツ
ストレス耐性細胞株をエンジニアリングするためのCRISPR戦略
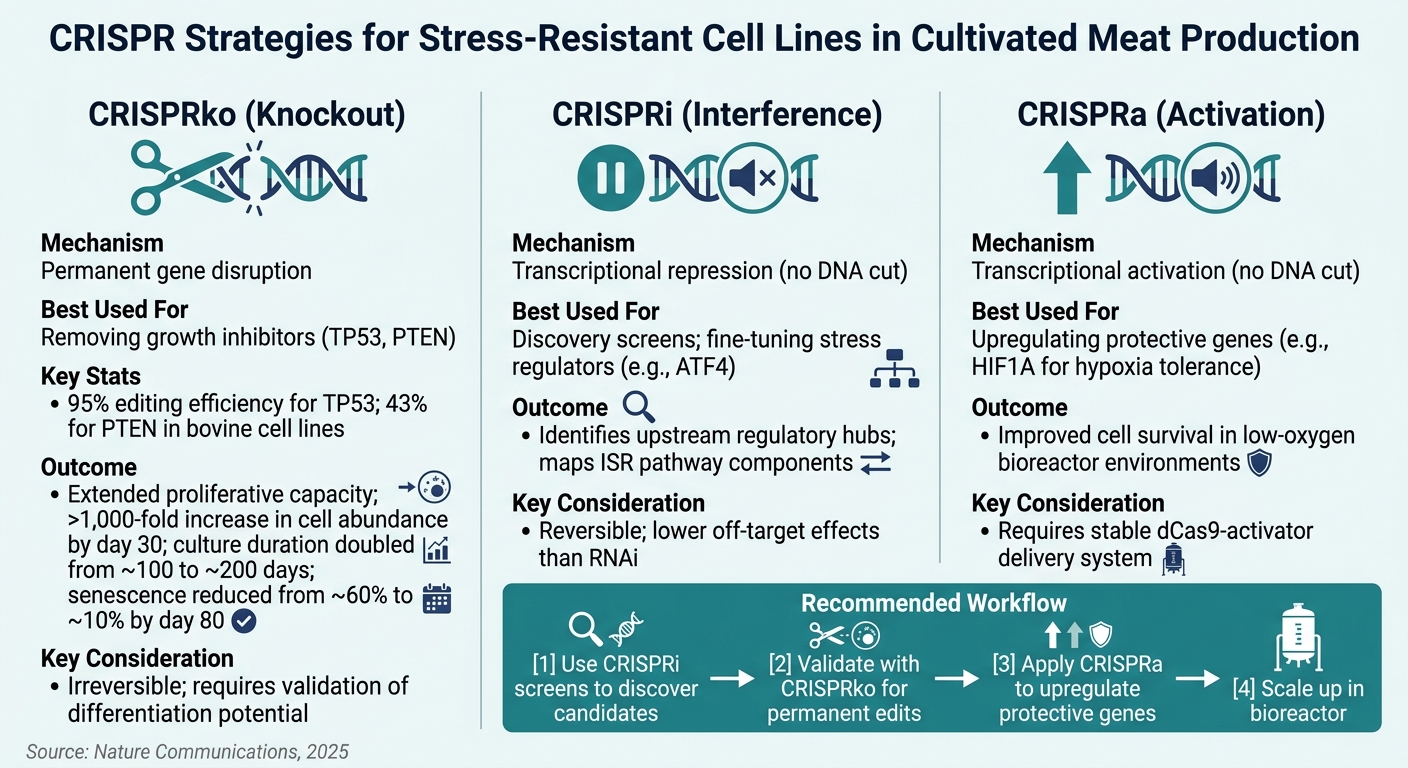
培養肉におけるストレス耐性細胞株のためのCRISPR技術
ストレス耐性のための主要な遺伝子と経路
詳細なストレスマップを手にしたら、次のステップは編集対象の遺伝子を特定することです。ターゲットの選択は、細胞のパフォーマンスに影響を与える主なストレッサーに依存します。
複製老化は、細胞の増殖を制限するため、培養肉の生産における主要な障害です。この分野の細胞源の約25%は間葉系幹細胞(MSCs)であり、繰り返し継代後に不可逆的な成長停止に直面します[1]. p53腫瘍抑制タンパク質をコードする遺伝子であるTP53, をノックアウトすることは、この問題に直接対処します。ウシMSCsの研究では、TP53ノックアウトが細胞の増殖能力を大幅に拡張し、未編集の系統の限界をはるかに超えて分裂できることが示されています[1]. 同様に、PTENをノックアウトすることで、PI3K/AKT/mTOR経路が強化され、ストレス耐性が向上します[1].
代謝およびミトコンドリアのストレスに対処するために、統合ストレス応答 (ISR) は重要な経路です。転写因子ATF4はミトコンドリアストレス応答を調整する中心的な役割を果たし、CRISPRスクリーンはその上流の調節因子をマッピングするのに役立っています [6]. カリフォルニア大学サンフランシスコ校のXiaoyan GuoとMartin Kampmannは次のように説明しています:
"転写または翻訳レポーターに基づく偏りのない遺伝子スクリーニングは、特定のストレス応答の調節因子を特定するための強力なアプローチです。" [6]
TGFβ経路も注目に値し、特にウシMSCの拡大において重要です。CRISPRスクリーンは、TGFβ駆動の軟骨分化が細胞増殖を抑制することを示しています。この経路を抑制することは、細胞を未分化で拡張可能な状態に維持するのに役立ちます[1] . 3D足場の密集したコアでよく見られる低酸素条件下では、CRISPRaを使用して HIF1Aを活性化することで、低酸素環境での細胞の生存率が向上します。これらの修正により、細胞は 産業規模のバイオリアクターの動的条件下で繁栄する能力を備えます.
しかし、細胞増殖を最大化する編集、例えばTP53ノックアウトは、細胞が筋肉や脂肪組織に分化する能力を低下させる可能性があることに注意することが重要です。成長と分化の可能性の間のこのトレードオフは、エンジニアリング戦略を設計する際に慎重にバランスを取る必要があります[1].
| ストレスファクター | 主要遺伝子ターゲット | CRISPR戦略 | 結果 |
|---|---|---|---|
| 複製老化 | TP53 | ノックアウト | 増殖能力の延長; 細胞量の増加 |
| 栄養/成長ストレス | PTEN | ノックアウト | PI3K/AKT/mTORシグナル伝達の強化; 生存率の向上 |
| ミトコンドリアストレス | ATF4 | CRISPRi / レポーター | 上流の調節経路の特定 |
| 低酸素 | HIF1A | CRISPRa(活性化) | 低酸素バイオリアクター環境での生存率の向上 |
| 軟骨形成ドリフト | TGFβ経路 | ノックアウト / 抑制 | ウシMSCにおける未分化で増殖的な状態の維持 |
主要な遺伝子が特定されたら、適切なCRISPR技術を選択することが次の重要なステップとなります。
CRISPR編集技術の比較
CRISPR法の選択は、遺伝子改変の精度と永続性を決定します。各アプローチには、永久的な変更、可逆的な調整、または探索的スクリーニングが目標であるかどうかに応じて、独自の強みがあります。
CRISPRノックアウト(CRISPRko)は、遺伝子を永久に無効化するための主要な方法です。TP53やPTEN, のように、完全な機能喪失が必要なターゲットに最適です。検証研究では、CRISPRkoがウシ細胞株においてTP53で95%、PTENで43%の編集効率を達成することが示されています[1]. これらの変動は、大規模な編集を進める前にターゲット特異的な効率をテストすることの重要性を強調しています。
CRISPR干渉(CRISPRi)は、遺伝子抑制を可逆的に行うことができ、発見段階に最適です。それはまた、RNAiと比較してオフターゲット効果を減少させます[6]. 一方、CRISPR活性化(CRISPRa)は、低酸素耐性に関与する保護遺伝子(HIF1A)や抗酸化防御を過剰発現させることによって、ストレス耐性を強化します。
技術の簡単な比較はこちらです:
| 技術 | メカニズム | 最適な用途 | 重要な考慮事項 |
|---|---|---|---|
| CRISPRko | 永久的な遺伝子破壊 | 成長阻害因子の除去 (TP53, PTEN) | 不可逆的;分化の可能性の検証が必要 |
| CRISPRi | 転写抑制(DNA切断なし) | 探索スクリーン;調整因子の微調整 | 可逆的;RNAiよりもオフターゲット効果が低い |
| CRISPRa | 転写活性化(DNA切断なし) | 保護遺伝子のアップレギュレーション (HIF1A) | 安定したdCas9-アクティベーターのデリバリーシステムが必要 |
ターゲットの特定を始めたばかりのチームにとって、プールされたCRISPRiスクリーニングは、大規模にストレス耐性遺伝子を発見するためのコスト効率の良い方法を提供します。Once promising candidates are validated, CRISPRko can be used for permanent edits suited to production. These approaches complement each other, and using them in sequence is increasingly seen as a best practice in the field [1][6].
培養肉研究に特化したCRISPR試薬やバイオリアクター用品の調達には、
CRISPR編集された細胞株の実装と検証
CRISPR編集の設計と提供
ターゲット遺伝子を特定したら、次のステップはCRISPR編集の設計と提供です。効果的な遺伝子破壊を確実にするために、重要なエクソンをターゲットとするシングルガイドRNA(sgRNA)の作成に焦点を当てます。このアプローチは、遺伝子を完全にノックアウトする可能性を高め、切り詰められた部分的に機能するタンパク質を生成するのを防ぎます。デュアルガイドRNA戦略を使用することで、ノックアウト効率を大幅に向上させ、約55%から95%以上に引き上げることができます[8].
選択するデリバリー方法は、特定の細胞タイプに依存します。培養肉細胞株の場合、事前に組み立てられたCas9リボヌクレオプロテイン(RNP)が最適な選択肢であることが多いです。これらのRNPは一過性であり、デリバリー後に迅速に分解されるため、オフターゲット効果を最小限に抑え、プラスミドDNAの統合リスクを回避します[8]. プールされたスクリーンやトランスフェクションが困難な一次細胞株が関与する場合、レンチウイルス導入は信頼性のある代替手段です。レンチウイルスシステムを使用する場合、研究者は通常、感染の多重度(MOI)を約0.3に低く保ち、複数の統合を避けることで、下流の分析を複雑にしないようにします[1].
最適な結果を得るために、細胞が対数増殖期にあり、トランスフェクション前に70–90%のコンフルエンスであることを確認してください。配送後、個々のクローンを限定希釈法や蛍光活性化セルソーティング(FACS)などの方法を使用して分離し、明確で曖昧さのない検証を行います。最後に、ゲノム、プロテオーム、および機能レベルで編集を確認し、成功を確認する必要があります。
編集された細胞株のスクリーニングと検証
編集された細胞株をバイオリアクター条件 に移行する際には、徹底的な検証が不可欠です。このプロセスには、ゲノム、プロテオーム、および機能の3つのレベルでのスクリーニングが含まれます。これらのステップを省略すると、生産条件下で失敗する可能性のある細胞株を選択するリスクが高まります。
ゲノムレベルでは、T7E1やサーベイヤーのようなミスマッチアッセイを使用して初期スクリーニングを行うことができ、細胞プール内の編集頻度を迅速に推定できます。正確な確認のために、サンガーシーケンシングまたは次世代シーケンシング(NGS)を使用して、二アレル破壊性インデルを持つクローンを特定します。[7][8]. プロテオミクスの検証は、通常ウェスタンブロット分析を使用して行われ、ターゲットタンパク質の完全な欠如を確認します。例えば、2025年に行われた研究では、TP53をノックアウトすることで、競争的スクリーニングの30日目に細胞の増加が1,000倍以上になり、培養期間が100日から約200日に倍増したことが示されました。[1].
機能的な検証も同様に重要です。代謝の生存率と増殖率は、Alamar Blueアッセイを使用して評価でき、200日以上にわたる集団倍加時間(PDT)の追跡は、複製老化を克服した細胞株を特定するのに役立ちます。[1]. 低酸素またはミトコンドリアストレスに耐えるように設計された細胞株に対して、FACSベースのレポーターアッセイは、低酸素または栄養制限条件下で細胞が正しく反応することを確認できます[6]. さらに、TP53またはPTENノックアウトを持つ細胞株は、分化能力を保持する能力についてテストする必要があります。CD29やCD44などの間葉系幹細胞(MSC)マーカーのフローサイトメトリーは、これらの細胞が幹細胞性を維持していることを確認できます[1].
| 検証レベル | 方法 | 目的 |
|---|---|---|
| ゲノム | サンガーシーケンシング / NGS | 二アレル破壊的インデルの確認[7][8] |
| プロテオミクス | ウエスタンブロット | ターゲットタンパク質の完全な欠如を確認[7][8] |
| 表現型 | フローサイトメトリー (CD29/CD44) | MSCマーカーと幹細胞性の保持を確認[1] |
| 機能的 | アラマーブルー / PDTトラッキング | 成長動態と代謝健康を評価[1] |
| ストレス | FACSベースのレポーターアッセイ | 困難な条件下でのストレス応答行動をテストする[6] |
編集された細胞株をスケールアップする前に、STRプロファイリングを実施して細胞の同一性を確認し、マイコプラズマ検査を行って汚染を排除する[7]. ノックアウト細胞株の作成には通常約3か月かかり、ワークフローの特定のステップを繰り返す可能性があります。
スケールアップ: ストレス耐性細胞株を生産に移行する
編集された細胞株をバイオリアクター条件に移行する
一度検証された編集細胞株は、ラボスケールの接着培養から、撹拌タンクバイオリアクター、エアリフトリアクター、または回転壁容器のような懸濁系に移行する必要があります。これらはすべて、工業規模の培養肉生産をサポートすることができます[2].
ウシ間葉系幹細胞(bMSC)のような接着依存性細胞の場合、ラミニン-511でコーティングされたマイクロキャリアを使用することで、懸濁培養への実用的な経路を提供します[3]. この移行中、CD29やCD44のようなMSCマーカーを監視して、細胞が分化能力を保持していることを確認することが重要です[1].
スケールアップの重要なステップには、培地の再配合が含まれます。血清ベースの培地は、化学的に定義された血清フリーの配合に置き換えられ、脂質、非必須アミノ酸、抗酸化物質が強化され、大規模条件下での細胞の生存率を維持します[4]. 特に、TP53およびPTENノックアウトを持つCRISPR編集された細胞株は、この移行により適しています。Nature Communications(2025年)に発表された研究では、これらの編集によりbMSCの増殖寿命が約100日から200日以上に延び、80日目までに老化が約60%からわずか10%に減少したことが示されました[1].
「TP53およびPTENのノックアウトは、増殖率を大幅に向上させ、老化を遅らせました。" - Nature Communications [1]
移行期間中、Alamar BlueアッセイやqRT-PCRのようなツールは、細胞の生存率を追跡し、遺伝子改変の安定性を確保するために不可欠です。これらのCRISPR編集されたウシ細胞株は、倍加率が平均12%改善し、50日目には50%の増加に達するものもあります[1]. 細胞がバイオリアクター条件で安定した性能を示したら、スケールアップに必要な専門機器の調達に焦点を移すことができます。
スケールアップのための機器と材料の調達
生産レベルのバイオリアクター運転へのスケールアップは、調達において重大な課題をもたらします。細胞の適応を確認した後、必要な材料と機器の取得が優先事項となります。使い捨て撹拌槽バイオリアクター、検証済みのマイクロキャリア、無血清培地成分、クローンの継続的なモニタリング用FACSシステムのようなアイテムは非常に専門的であり、一般的な実験室の供給業者からは入手できないことが多いです。
培養肉産業向けに特別に設計された
結論
CRISPR技術は、研究ツールから培養肉生産における細胞株を工学的に操作する実用的な方法へと移行しました。TP53やPTENのような主要な調節因子を標的にすることで、研究者は細胞増殖を大幅に延長し、通常の培養期間を実質的に倍増させました [1]. この進展は、培養肉のスケーラブルな生産の限界を押し広げます.
しかし、編集された細胞株から本格的な生産に至るまでの過程では、各ステップでの徹底的な検証が必要です。工学的に操作された細胞が筋肉や脂肪組織に分化する能力を維持することは、急速な増殖を達成することと同様に重要です。これがなければ、どんなに急成長する細胞株でも商業的な実行可能性を欠くことになります[1]. これは、改善された増殖が意味のある生産成果に結びつくことを確認するための厳格な検証プロセスの必要性を強調しています。
Nature Communicationsはこのアプローチを強化し、次のように述べています:
"これらの発見は、ウシ幹細胞の特性を最適化するためのCRISPRスクリーニングの有用性を示しており、将来的によりスケーラブルな培養肉生産への道を提供します。" [1]
これらの進展にもかかわらず、調達のような実際的な課題が進捗を妨げることがあります。sgRNAライブラリー、使い捨てバイオリアクター、無血清培地の一般的なサプライヤーへの依存は、しばしば互換性の問題や遅延を引き起こします。
適切な材料の入手可能性は、遺伝子工学そのものと同じくらい重要です。Nature Communicationsによると、培養肉は従来の肉に代わる有望な選択肢を提供しますが、スケーラビリティとコスト効率は依然として大きな課題です。CRISPRベースのエンジニアリングは、規律あるバイオプロセス設計と
FAQs
CRISPRターゲットを選択する前にどのバイオリアクターストレスをプロファイルすべきですか?
培養肉生産においてストレス耐性のある細胞株を開発するためにCRISPRターゲットを選択する際、細胞の成長と生存に影響を与える主要なバイオリアクターストレスを評価することが重要です。これらのストレスには以下が含まれます:
- せん断応力: バイオリアクター内の細胞は、混合や曝気からの機械的な力にさらされることがよくあります。長時間のせん断応力は細胞膜を損傷し、成長を妨げる可能性があります。
- 酸素レベル: 最適な酸素濃度を維持することが重要です。酸素が少なすぎるとエネルギー生産が制限され、過剰な酸素は酸化ストレスを引き起こす可能性があります。
- 栄養素の利用可能性 : 細胞は一貫した栄養供給を必要とします。バランスの崩れや枯渇は増殖と生産性を妨げる可能性があります。
- pHの変動: 細胞は狭いpH範囲内で最適に機能します。逸脱は代謝プロセスや酵素活性を妨げる可能性があります。
- 温度の変動: わずかな温度変化でも細胞機能に影響を与え、ストレスや生存率の低下を引き起こす可能性があります。
- 廃棄物の蓄積: 代謝副産物が効率的に除去されない場合、有毒になり細胞の成長を阻害する可能性があります。
これらのストレス要因を徹底的に理解することで、研究者は重要なストレス応答経路を特定できます。この知識により、CRISPRを使用した標的遺伝子改変が可能になり、細胞株の耐性が向上し、バイオリアクター条件でのより堅牢な性能が保証されます。
成長を早める編集と筋肉と脂肪の分化をどのようにバランスさせますか?
培養肉の生産において急速な成長と筋肉と脂肪の分化をバランスさせるには、遺伝子と培養条件の慎重な管理が必要です。CRISPR技術はここで中心的な役割を果たし、TP53やPTENなどの遺伝子の標的修正を可能にします。. これらの調整により、細胞の増殖を促進しつつ、筋肉と脂肪組織に分化する能力を保持することができます。
培養条件の微調整と遺伝子発現の調整も、望ましいバランスを達成するために同様に重要です。
バイオリアクターのスケールアップ前に必要な最小限の検証は何ですか?
バイオリアクターに移行する前に、遺伝子改変された細胞株が安定した望ましい特性、例えば成長率の向上、ストレス耐性、分化能力を維持していることを確認することが重要です。この検証プロセスでは、遺伝的安定性を評価し、バイオプロセス条件下での一貫した性能を確保する必要があります。マルチオミクス解析とストレス応答プロファイリングからのデータがこの評価の鍵となります。ハイスループットCRISPRスクリーニングを使用することで、細胞増殖と寿命を向上させる遺伝子編集を特定し、これらの細胞株をスケーラブルな培養肉生産により適したものにすることができます。










