

Hücreleri genişletebiliyorsanız ancak onları doğru zamanda doğru kadere yönlendiremiyorsanız, süreciniz farklılaşmada duracaktır. Buradaki ana nokta şudur: sentetik gen devreleri, medya değişikliklerinin genellikle heterojen, kısmen taahhüt edilmiş popülasyonlar bıraktığı yerlerde, bağlılık, zamanlama, hafıza ve soy karışımı üzerinde hücre içi kontrol sağlar.
Eğer bir kültive edilmiş et farklılaşma iş akışı oluşturuyor olsaydım, bu makaleden hemen dört noktayı alırdım:
- Yapıdan değil, yerel ağdan başlayın. Hücrelerin nerede durakladığını, sürüklendiğini veya yanlış kadere yöneldiğini bulmak için snRNA-seq, trajektori analizi, GRN çıkarımı ve miRNA profilini kullanın.
-
Devre türünü süreç sorununa göre eşleştirin.
Bir anahtar düğmesi kilitleme için uygundur, bir ileri besleme veya bant geçiren tasarım zamanlama kontrolü için uygundur, bir mantık kapısı çoklu sinyal geçişi için uygundur ve miSFITs kademeli çıktı için uygundur. - İlk günden itibaren düşük sızıntı, düşük gürültü ve güvenlik için tasarlayın. Ortogonal parçalar, negatif oto-regülasyon, iFFL'ler, cm transgenleri ve indüklenebilir öldürme veya büyüme-durdurma modülü yapının bir parçasıdır, sonradan düşünülmüş bir şey değildir.
- Erken aşamada ölçekle ilgili koşullar altında doğrulayın. 2D'de çalışan bir devre, 3D, mikro taşıyıcı veya karıştırılmış süspansiyon içinde indükleyici gradyanlar, oksijen sınırları ve kesme nedeniyle değişebilir.
Makale ayrıca süreç ekipleri için önemli olan pratik bir noktaya değiniyor: tek soy kontrolü ve oran kontrolü farklı işlerdir. Bir Tet-On MyoD kaseti miyojenik girişe itebilir, ancak bütün kesim ürünleri kas, yağ ve ECM oranlarının, kontrolünü gerektirir ki bu genellikle geri bildirim, parakrin sinyalizasyon ve daha yoğun klon taraması anlamına gelir.
Bu mesajı vurgulayan birkaç rakam:
- Standart miyojenik farklılaşma, yaklaşık %50–60 füzyon indeksleriyle durabilir
- iPSC'lerdeki mühendislik ürünü GRN'ler, hedef soy farklılaşmasını %52'den %81'e çıkardı
- Modifiye edilmiş MSC'lerdeki sentetik devreler, kardiyak farklılaşmayı %76'ya çıkardı
- Bazı domuz Tet-On-PAX7 hatları, 40 pasajdan fazla yüksek miyojenik potansiyelini korudu
- İnsan pluripotent kök hücrelerinin yaklaşık %20'si kanserle bağlantılı mutasyonlar taşıyabilir, bu yüzden indüklenebilir güvenlik modülleri önemlidir
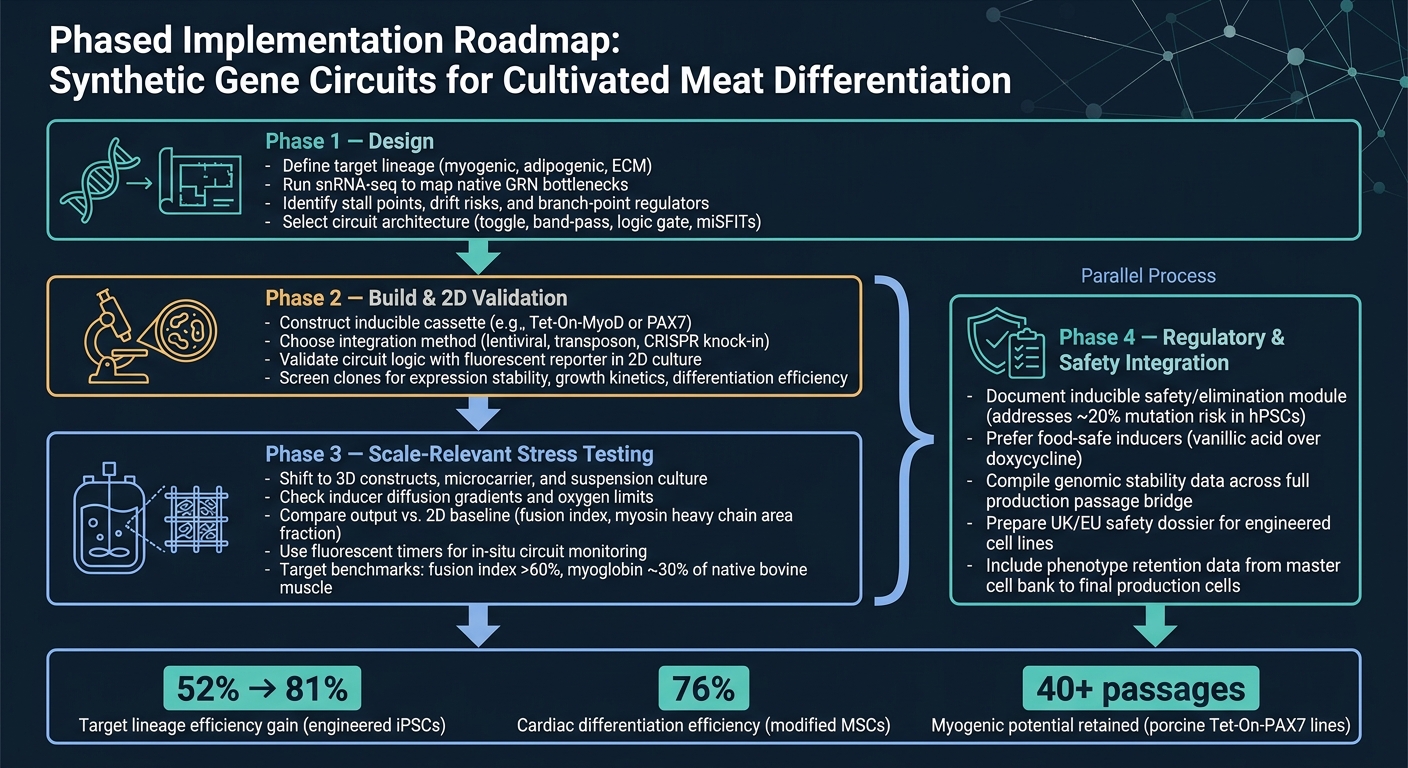
Kültive Edilmiş Et Farklılaşması için Sentetik Gen Devresi İş Akışı
Bilim İnsanı Hikayeleri: Michael Elowitz, Canlı Hücrelerde Genetik Devreler
Hızlı karşılaştırma
| Konu | Makalede söylenenler basit terimlerle |
|---|---|
| Ana darboğaz | Farklılaşma, genişleme değil |
| Neden devreler yardımcı olur | Hücre içinde eşik kontrolü ve hücre durumu hafızası eklerler |
| En iyi ilk adım | Yerel kader kontrolünü snRNA-seq ve ilgili araçlarla haritalayın |
| Ana devre seçenekleri | Geçiş, ileri besleme, bant geçişi, mantık kapıları, miSFITs, CRISPRa/i |
| Miyogenez örneği | Tet-On-MyoD büyümeyi terminal farklılaşmadan ayırmak için |
| Adipogenez / ECM kontrolü | miSFITs ve dereceli PPARG/BMP4 tipi çıktı için papyon tasarımları |
| Tam kesim zorluğu | Kas, yağ ve bağ dokusu arasında oran kontrolü |
| Ölçek büyütme riski | 2D davranış 3D veya biyoreaktörlerde geçerli olmayabilir |
| Entegrasyon seçenekleri | Lentivirüs, transpozonlar, CRISPR knock-in, episomal vektörler |
| Düzenleyici nokta | Mühendislik ürünü hatlar daha geniş bir güvenlik paketi gerektirir; vanilik asit gibi gıda güvenli indükleyiciler mümkün olduğunda DOX yerine tercih edilir |
Yani, basit bir ifadeyle: bu sadece bir devre tasarımı makalesi değil. Bunu, devre mimarisini, soy biyolojisini, klon seçimini, biyoreaktör performansını ve İngiltere/AB güvenlik belgelerini tek bir farklılaştırma stratejisine bağlama rehberi olarak okurdum.
Yerel ağ haritalamasından yapı seçimine, ölçek büyütme kontrollerine ve düzenleyici uyuma kadar tam yolu istiyorsanız okumaya devam edin.
2. Farklılaştırma kontrol devreleri için tasarım ilkeleri
2.1 Devreyi tasarlamadan önce yerel hücre kaderi ağını haritalayın
Bir devre tasarlamadan önce, hücrenin zaten ne yaptığını net bir şekilde anlamanız gerekir.
Tek çekirdekli RNA dizilemesi (snRNA-seq) başlamak için iyi bir yerdir. Bu, NOTCH2 ve HEYL , ile işaretlenmiş yedek hücreler dahil olmak üzere sessiz alt popülasyonları gösterebilir ve farklılaşmayı iyileştirebilecek yol hedeflerine işaret edebilir [3].
Oradan, yörünge analizi ve gen düzenleyici ağ (GRN) çıkarımı, düzenleyici aktivasyon sırasını haritalamaya yardımcı olur ve hücrelerin en olası duraklama noktalarını vurgular. Miyogenezde, ana kaskad MYOD1 ve MYOG üzerinden geçer. Adipogenezde, ana düğümler PPARG ve CEBPA, fibro-adipojenik progenitör (FAP) kaderi ana dal noktası riski olarak hareket eder. Aşağıdaki tablo ana düzenleyicileri ve darboğazları özetlemektedir.
| Soy hattı | Ana düzenleyici ustalar | Kritik sinyal yolları | Belirlenen darboğazlar |
|---|---|---|---|
| Miyojenik | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Rezerv hücre oluşumu (uyuşukluk) |
| Adipojenik | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Fibro-adipojenik progenitör (FAP) kaderi |
| Pluripotent | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Spontan farklılaşma / heterojenlik |
Başka bir faydalı katman miRNA ifade profillemesi. Endojen miRNA'lar, örneğin miR-302a, pluripotensiye bağlı ve miR-375, farklılaşmaya bağlı, algıla-ve-yanıtla tasarımlarında iç sınıflandırıcılar olarak işlev görebilir.Bu, devrenin hücrenin gerçek durumunu okumasına olanak tanır, yalnızca harici indükleyicilere bağlı kalmak yerine [5].
Bu darboğazlar devre seçimini yönlendirmelidir. Ana sorun kayma ise, bir kilitleme gerekebilir. Zamanlama sorun ise, bir darbe daha uygun olabilir. Kader kontrolü birden fazla ipucuna bağlıysa, çok girişli mantık genellikle daha mantıklıdır.
2.2 Doğru devre mimarisini seçin
Bu, ödünleşimlerin açıkça ortaya çıktığı yerdir. Doğru mimari, üç pratik noktaya bağlıdır: taahhüdün ne kadar kalıcı olması gerektiği, zamanlamanın ne kadar sıkı kontrol edilmesi gerektiği ve hücrenin sorunsuz taşıyabileceği genetik yük miktarı.
Bistable toggle switches, soy hattı taahhüdünün kilitli kalması gerektiği durumlara uygundur. Ana sorun, transkripsiyonel gürültü nedeniyle meydana gelen kendiliğinden durum değişiklikleridir.
Bant geçiren filtreler , bir transkripsiyon faktörünün yalnızca belirli bir gelişimsel pencerede gerektiği durumlara uygundur. Buradaki püf noktası, indükleyici seviyelerinin sıkı bir şekilde kontrol edilmesi gerektiğidir, aksi takdirde zamanlama kayabilir.
Mantık kapıları , aynı anda birden fazla girdi gerektirerek özgüllük ekler. Örneğin, farklılaşmaya yalnızca bir eksojen indükleyici mevcut olduğunda ve hücre doğru endojen miRNA profilini gösterdiğinde izin verilebilir. Bu, hedef dışı bağlılık riskini azaltmaya yardımcı olur.
Aşağıdaki tablo, ana mimarileri ve bunlarla birlikte gelen ödünleşimleri özetlemektedir.
| Mimari | Tersine Çevrilebilirlik | Zamansal hassasiyet | Entegrasyon karmaşıklığı | Birincil kullanım durumu | Ana risk |
|---|---|---|---|---|---|
| İki kararlı anahtar | Düşük (kilitlendikten sonra) | Orta | Orta | Kalıcı soy hattı taahhüdü | Gürültü nedeniyle kendiliğinden geçiş |
| Bant geçiren filtre | Yüksek (konsantrasyona bağlı) | Yüksek | Yüksek | Geçici gelişim aşamaları | Sıkı indükleyici kontrolü gerektirir |
| Mantık kapısı (VE/VEYA/DEĞİL) | Değişken | Orta | Orta–yüksek | Hücre tipi spesifik aktivasyon | KAPALI durumlarda sızıntı | Papyon / çoklu giriş | Yüksek | Orta | Orta | Çoklu sinyal entegrasyonu | Endojen miRNA kararlılığına dayanır |
| miSFITs | Yüksek | Orta | Düşük–orta | Kademeli çıktı ayarı | Zayıf ayarlanmışsa dar dinamik aralık |
"Fonksiyonelliği korurken hesaplama katmanlarının sayısını en aza indirerek, bu strateji gen devresi mühendisliğinde ölçeklenebilirlik engellerini ele alır." - Nature Communications [9]
Her eklenen düzenleyici katman yükü artırır ve hücresel kaynakları kullanır. Pratikte, iki tasarım aynı işi yapıyorsa, ölçek önemli olduğunda genellikle daha basit olanı tercih edilir.
Mimari belirlendikten sonra, bir sonraki iş düşük sızıntı, gürültü bastırma ve emniyetli kontrol altında dayanmasını sağlamaktır.
2.3 Güvenilirlik, düşük sızıntı ve güvenlik için inşa et
Bir devre uzun süreli kültür boyunca kararlı kalmalıdır. Üretim kullanımı için kısa süreli iyi performans yeterli değildir.
Ortogonal parçalar ilk savunmadır. Yerel makine ile çapraz etkileşime girmeyen promotörler, transkripsiyon faktörleri ve düzenleyici elemanlar, hedef dışı etkileri sınırlamaya yardımcı olur ve endojen sinyallerin devreyi açma veya kapatma olasılığını azaltır.Yüksek sıkılıkta modifiye edilmiş promotörler, PCREm gibi, indüklenebilir memeli sistemlerinde bazal sızıntıyı azaltmak için kullanılmıştır [6] .
Negatif oto-regülasyon mümkün olduğunda eklenmeye değerdir. Transkripsiyonel gürültüyü azaltmak ve indükleyici konsantrasyonuna daha doğrusal bir yanıt üretmek için bilinen daha iyi motiflerden biridir [6]. Uyumsuz ileri besleme döngüleri (iFFLs), hücrelerin kısa süreli gürültü dalgalanmaları yerine sürdürülebilir sinyallere yanıt vermesini sağlayarak stokastik dalgalanmaları filtreleyerek başka bir kontrol katmanı ekleyebilir. Kodon-modifiye edilmiş ( cm) sentetik transkripsiyon faktörlerinin versiyonları da karakterizasyonu kolaylaştırır. Devre kaynaklı ifadeyi, doğrulama sırasında endojen genomik (g) ifadeden ayırmanıza olanak tanır [1]. Bu küçük bir detay gibi görünebilir, ancak bir okumanın devreden mi yoksa ana genomdan mı geldiğini anlamaya çalışırken zaman kazandırır.Güvenlik modülleri gereklidir. İnsan pluripotent kök hücrelerinin yaklaşık %20'si kanserle ilişkili mutasyonlar taşır [7]. Bu nedenle, bir devre kök hücre türevli bir hatta giriyorsa, indüklenebilir bir büyüme-durdurma veya yok etme modülü içermelidir. Vanilik asit burada öncelik verilmesi gereken faydalı bir indükleyicidir çünkü lisanslı bir gıda katkı maddesidir ve bu da onu kültive edilmiş et hücre hatlarında devre tetikleyicisi olarak kullanma durumunu destekler [1] .
"Sentetik biyoloji, mühendislerin devreleri kolayca ve hassas bir şekilde birden fazla genin ifadesini ayarlamak için kullanmalarına... potansiyel olumsuz hedef dışı yan etkileri ortadan kaldırmalarına olanak tanır." - npj Systems Biology and Applications [6]
Bu seçimler, Bölüm 3'te soy-spesifik devreleri kurar.
sbb-itb-ffee270
3. Miyojenik, adipogenik ve oran kontrollü farklılaşma için devre stratejileri
3.1 Genişlemeyi terminal farklılaşmadan ayıran miyojenik devreler
Devre mimarisi belirlendikten sonra, bir sonraki iş soy-spesifik dağıtımdır. Miyogenez için ana problem basitçe ifade edilebilir ancak uygulaması zordur: hücrelerin önce çoğalması, ardından istenildiğinde farklılaşmaya geçmesi gerekir, her iki yönde de çok erken kaymadan.
Bir Tet-On-MyoD kaseti bunu yapmanın en doğrudan yollarından biridir. Bu kurulumda, hücreler doksisiklin (DOX) yokken standart koşullar altında çoğalır. DOX ekleyin ve devre miyojenik bağlılığı yönlendirir.Çin'deki çeşitli kurumlarda araştırmacılar bu yaklaşımı tavuk embriyonik fibroblastlarında kullandı ve indüksiyondan sonra etkili miyotüp oluşumu bildirdiler [4].
Bir iki kararlı geçiş anahtarı daha sıkı durum kontrolü sunar. E-KRAB ve Pip-KRAB gibi karşılıklı olarak engelleyici baskılayıcılardan oluşturulan sistemler, kısa bir DOX darbesini kararlı bir miyojenik programa dönüştürebilir [6]. Pratikte bu, farklılaşmış durumun sinyal kaldırıldıktan sonra kaybolmak yerine kilitli kalması anlamına gelir. Negatif oto-regülasyon eklemek de stokastik gürültüyü azaltmaya ve genişleme aşamasında sızıntı farklılaşmasını sınırlamaya yardımcı olur [6].
Bu önemlidir çünkü standart in vitro miyojenik farklılaşma genellikle yarı yolda durur. Füzyon indeksleri genellikle sadece %50–60 civarındadır, bu da büyük bir birleşmeyen yedek popülasyon bırakır [3]. Devre odaklı MyoD aktivasyonu bağlılığı artırabilir, ancak bağlılık tek başına uniform füzyonu garanti etmez. MyoD devreleri MEK, NOTCH ve RXR modülasyonu ile birleştirildiğinde, 2D kültürde neredeyse tam füzyon mümkün hale gelir [3]. Devre, soy girişinin zamanlamasını kontrol eder; küçük moleküller daha temiz ve daha uniform bir hücre döngüsü çıkışı sağlar.
3.2 Kompozisyon ve yapı için adipogenik ve fibrogenik kontrol
Yalnızca kas yeterli değildir. Miyojenik kontrol sağlandığında, bir sonraki mesele kompozisyondur: ne kadar yağ oluşur, ne kadar ECM birikir ve bu programlar ne zaman devreye girer. Burada, ikili AÇIK/KAPALI kontrol genellikle çok keskindir. Takımların genellikle ihtiyaç duyduğu şey, özellikle adipogenik-fibrogenik dal noktasında, kademeli çıktıdır.
miSFITs, ifadeyi adım adım ayarlamak için pratik bir yol sunar.Mutasyona uğramış miRNA hedef bölgelerini - örneğin, miR-17 için bölgeler - PPARG veya BMP4 gibi çıktı genlerinin 3′UTR'sine yerleştirerek, araştırmacılar varyant bir kütüphaneden ifade seviyelerini seçebilirler [5]. Bu, adiposit indüksiyonunu bir ışık anahtarından ziyade bir dimmer anahtarına benzetir. Hücreleri tamamen ya da hiç tepki vermeye zorlamak yerine, ekipler adipogenezi daha dikkatli bir şekilde ayarlayabilir [5].
Fibroblastlar burada sadece seyirci değildir. Doku yapısını şekillendiren ECM proteinlerini sağlarlar [10]. Bu, fibrogenik kontrolü sadece bir yan mesele değil, ürün tasarımının bir parçası yapar. Devreler, fibrogenik ve adipogenik durumlar arasındaki geçişi yönetmeye yardımcı olabilir ve kültive edilmiş kümes hayvanlarında, fibroblastlarda doğrudan PPARG aktivasyonu anlamlı yağ birikimi sağlamak için gerekli olabilir [10].
Bir papyon mimarisi bu soruna iyi uyar çünkü algılama ve çıktıyı ayrı tutar. Algılama katmanı hücrenin mevcut durumunu okurken, çıktı katmanı PPARG, CEBPA veya diğer soy düzenleyicilerini ayarlar. Bu ayrım, hücreler doğru gelişim aşamasına ulaşmadan önce adipogenik veya bağ dokusu programlarının açılmasını engellemeye yardımcı olur.
3.3 Çoklu soy oranı kontrolü ve geri bildirim algılama
Oran kontrol devreleri farklı bir sorunla ilgilenir: farklılaşmanın olup olmadığı değil, nihai popülasyon karışımının olması gerektiği yerde kalıp kalmadığı. Tüm kesim ürünleri için kas, yağ ve ECM'nin doğru oranlarda elde edilmesi, herhangi bir soyun farklılaşmasını sağlamak kadar önemlidir.
Bu sistemler geri bildirim kontrolünü hücrelerin içine inşa eder. Duruma özgü promotörler, belirli bir soya bağlı kalmış hücrelerle sınırlı sinyal protein ifadesini kısıtlar.Sentetik parakrin modüller, ardından, taahhüt edilmiş miyojenik hücrelerin, yakın hücrelerde adipogenik taahhüdü baskılayan bir inhibitör sinyal salmasına izin verir. Mantık, Delta-Notch gelişim sistemlerindeki lateral inhibisyona benzer [1][6]. Dallanmanın daha karmaşık hale geldiği yerlerde, çok girişli kapılar, hücre dışı ipuçlarını içsel durum sinyalleriyle birleştirebilir [9] .
miSFIT'ler de bu aşamada çalışır. BMP4 veya diğer morfogenlerin çıktı gücünü ayarlayarak, ekipler yukarı akış karar mantığını yeniden yazmadan soy dengesi değiştirebilir. Mühendislik ürünü gen düzenleyici ağlarda, bu tür bir kontrol, hedef soy farklılaşma verimliliğini kontrollerde %52'den mühendislik ürünü iPSC'lerde %81'e çıkardı [2]. Modifiye edilmiş mezenkimal kök hücrelerde, sentetik devreler kardiyak farklılaşma verimliliğini %76'ya çıkardı [2].
Aşağıdaki tablo, üretimde en önemli olan noktaları kullanarak tek soy ve oran kontrolü yaklaşımlarını karşılaştırır.
| Özellik | Tek soy devresi | Oran kontrol devresi |
|---|---|---|
| Karmaşıklık | Düşük; tipik olarak tek bir indüklenebilir promotör ve düzenleyici [4] | Yüksek; mantık kapıları ve parakrin sinyalizasyon gerektirir [6][9] |
| İzleme yükü | Düşük; genellikle tek bir raporlayıcıyı takip eder [4] | Yüksek; birden fazla soy belirtecinin izlenmesini gerektirir [5] |
| Dayanıklılık | Orta; heterojenlik ve rezerv hücre oluşumuna eğilimli [3] | Yüksek; popülasyonları dengelemek için geri bildirim ve lateral inhibisyon kullanır [1] |
| Üretim değeri | Biyokütle için yüksek; karmaşık doku yapısı için sınırlı [10] | Kas, yağ ve ECM gerektiren bütün kesim ürünleri için gereklidir [4] |
Oran-kontrol devreleri daha ağır bir doğrulama yükü ekler.Ancak, yerleşik geri bildirimleri yalnızca süreç kontrolü ile eşleştirmek zordur, bu da klon seçimi ve süreç testine daha fazla baskı uygular.
4. Yapıdan sürece: doğrulama, ölçek büyütme ve düzenleyici uyum
4.1 Kararlı performans için entegrasyon stratejisi ve klon seçimi
Devre tasarımından sonra zor kısım başlar: bu devreyi üretim ölçeğinde pasajlama boyunca kararlı kalacak şekilde hücrelere yerleştirmek.
Lentiviral teslimat genellikle verimlidir ve hızlı bir şekilde kararlı entegrasyonlar sağlar. Ancak entegrasyon rastgeledir. Bu, daha fazla düzenleyici dikkat gerektirir ve susturma nedeniyle ifadenin zamanla düşme riski taşır. Transpozon sistemleri gibi PiggyBac ve Sleeping Beauty ortada yer alır. Birçok pasaj boyunca performansı koruyabilirler, ancak yine de kopya sayısı ve yerleştirme yerleri için tarama yapmanız gerekir. Uyuyan Güzel, örneğin, TERT ve CDK4'ü aşırı eksprese ederek sığır uydu hücrelerini kararlı bir şekilde ölümsüzleştirmek için kullanılmıştır ve hatlar 40 geçişin ötesinde miyojenik potansiyelini korur [10]. CRISPR knock-in yapının nereye yerleştiği üzerinde en sıkı kontrolü ve en hassas genomik ayarı sağlar, ancak klon seçimi daha yavaş ve verim daha düşüktür.
| Entegrasyon Yöntemi | Yerleştirme Kontrolü | Stabilite | Ölçeklenebilirlik | Düzenleyici Hususlar |
|---|---|---|---|---|
| Lentiviral Teslimat | Düşük (rastgele entegrasyon) | Yüksek, ancak susturulmaya eğilimli | Yüksek | Rastgele yerleştirme ve viral kalıntılar nedeniyle daha fazla düzenleyici inceleme gerektirir |
| Transpozonlar (PiggyBac/SB) | Orta | Birçok pasaj boyunca yüksek | Yüksek | Kopya sayısı ve yerleştirme yerleri için tarama gerektirir |
| CRISPR Knock-in | Yüksek (yer-spesifik) | Çok yüksek | Orta | Olumlu; endojen genleri bozma riskini azaltır |
| Episomal Vektörler | Yok (ekstrakromozomal) | Düşük; bölünme sırasında kaybolabilir | Düşük | Daha düşük entegrasyon engeli, ancak uzun vadeli genişleme için uygun değil |
Klon taraması, yapının mevcut olduğunu doğrulamanın ötesinde daha fazlasını yapmalıdır.İfade kaymasını, yerleştirme profilini, büyüme kinetiğini, farklılaşma verimliliğini ve yüksek pasaj numarasında fenotip korunmasını izlemelidir. snRNA-seq burada faydalıdır çünkü ölçek büyütmeden önce farklılaşmadan hücre döngüsünden çıkan Pax7⁺/Ki-67⁻ rezerv hücreleri ile zenginleştirilmiş klonları dışlayabilir [3]. Tet-On-PAX7 devresine sahip domuz EPSC'leri, 3D mikro taşıyıcı ve süspansiyon kültüründe 40'tan fazla pasaj boyunca yüksek kas farklılaşmasını korudu [8].
4.2 Devre davranışının 3D, mikro taşıyıcı ve biyoreaktör kültüründe nasıl değiştiği
Bir klon elde ettiğinizde, bir sonraki test, 2D dışında aynı şekilde davranıp davranmadığıdır. Çoğu durumda, davranmaz. 2D'deki performans, süspansiyon, mikro taşıyıcı veya iskele kültürüne temiz bir şekilde nadiren taşınır çünkü difüzyon gradyanları, oksijen sınırları ve kesme kuvveti devre çıktısını değiştirir.
İlk kontrollerden biri indükleyici difüzyonudur. Karıştırmalı tank biyoreaktörlerinde, küçük moleküllü indükleyicilerin hücrelere eşit şekilde ulaşması gerekir. Pratikte, özellikle yoğun mikrokültürlerde ve agregatlar veya iskelet çekirdekleri içinde gradyanlar oluşabilir. Süspansiyon kültürü genellikle büyük ölçekli yetiştirilen et için daha uygun bir seçenektir çünkü daha yüksek hücre yoğunluğunu destekler ve daha sıkı süreç kontrolü sağlar.
Sistem büyüdükçe hücre durumunu izlemek de zorlaşır. 2D'de mikroskopi ile kolayca okunabilen floresan raporlayıcı sinyaller, opak 3D yapılar içinde çözülmesi zor hale gelebilir. Floresan zamanlayıcılar - bir proteinin olgunlaşmasıyla emisyonu mavi renkten kırmızıya kaydıran problar - gerçek zamanlı devre aktivasyon verilerini in situ [1]. sağlayabilir.Mantıklı yol, aşamalı doğrulamadır: önce 2D'de, sonra 3D yapılarında, ardından nihai biyoreaktör koşulları altında [3][8].
4.3 Karakterizasyon, gıda güvenliği belgeleri ve Birleşik Krallık/AB değerlendirmeleri
Proses testinden sonra, karakterizasyon devre fonksiyonunun, fenotipin ve güvenliğin hala geçerli olduğunu göstermelidir. Temel veri yığını, akış sitometrisi, cm dizileri ile qPCR, RNA-seq zaman kursları ve miyozin ağır zincir alan fraksiyonu ve miyoglobin ekspresyonu gibi fonksiyonel çıktıları içermelidir [1]. Optimizasyonlu serumsuz farklılaşma medyalarının, yerli sığır kasında bulunan seviyelerin yaklaşık %30'una kadar miyoglobin ekspresyonunu getirdiği gösterilmiştir [3]. Bu, ekiplere belirsiz bir hedef yerine net bir kıyaslama sağlar.
Protein, amino asit ve yağ profillerini, duyusal özelliklerle birlikte belgelemeniz de gerekiyor [10][3].
Düzenleyici açıdan bakıldığında, Birleşik Krallık ve AB, kendiliğinden ölümsüzleşmiş (GDO olmayan) hücre hatları ile genetik olarak mühendislik yapılmış hatlar arasında net bir çizgi çizer. İkincisi, daha geniş güvenlik dosyalarına ihtiyaç duyar [10][3]. Stabilite paketleri, ana hücre bankasından nihai üretim hücrelerine kadar - tam üretim köprüsü boyunca - fenotip korunumu ve genomik stabiliteyi göstermeli ve izlenebilirlik kayıtları aradaki her geçişi hesaba katmalıdır [10]. Eğer devre kimyasal bir indükleyiciye bağlıysa, vanilik asit gibi gıda güvenli veya lisanslı bir katkı maddesi, doksisikline tercih edilir [1].
Rutin genomik gözetim bir zorunluluktur ve indüklenebilir bir intihar veya eliminasyon anahtarı, temel bir risk kontrol önlemi olarak belgelenmelidir [7]. Fonksiyonu, özellikle İngiltere ve AB'nin kültive edilmiş et için kuralları şekillendirmeye devam ederken, güvenlik dosyasına da dahil edilmelidir.
5. Pratik yol haritası ve sonuç
5.1 Kültive edilmiş et ekipleri için aşamalı bir uygulama yol haritası
Konseptten üretime en temiz yol, aşamalı bir iş akışıdır.
Aşama 1 tasarımdır. Hedef soy hattını tanımlayarak başlayın, ardından bir devre mimarisi seçmeden önce ana darboğazları doğrulamak için snRNA-seq kullanın. Bu adım önemlidir çünkü bir devre yalnızca gerçekten tanımladığınız kısıtlamaları çözebilir.
Aşama 2 inşa ve 2D doğrulamadır. Yapıyı inşa edin ve devrenin 2D'de, basit bir raporlayıcı okuması kullanarak, amaçlandığı gibi davrandığını kontrol edin.Bu aşamada, hedef basittir: daha zor ve maliyetli modellere geçmeden önce mantığın çalıştığını doğrulamak.
Aşama 3, ölçekle ilgili stres testidir. 3D sistemlere ve biyoreaktörle ilgili koşullara geçin, ardından çıktıyı 2D temel çizgisiyle karşılaştırın. Bu, birçok tasarımın zayıf noktalarını göstermeye başladığı yerdir, özellikle kütle transferi, kesme ve matris etkileri devreye girdiğinde.
Aşama 4, düzenleyici ve güvenlik entegrasyonudur ve Aşama 3 ile paralel olarak yürütülmelidir. Güvenlik ve düzenleyici çalışmaların sona kadar beklememesi gerekir. Ölçek büyütme ile birlikte, herhangi bir indüklenebilir güvenlik modülü için belgeleri de içerecek şekilde yürütün.
5.2 Cellbase aracılığıyla etkinleştirici araç ve malzemelerin temini
İş akışı belirlendikten sonra, temin genellikle hızı sınırlayan adım haline gelir.
- hücre hatları
- serumsuz ve kimyasal olarak tanımlanmış ortamlar
- iskeletler
- biyoreaktör bileşenleri
- sensörler
- analitik ekipmanlar
Her aşamada uyumlu malzemelere güvenilir erişim, devre davranışının ölçekle ilgili koşullar altında ne kadar hızlı karakterize edilebileceği üzerinde doğrudan bir etkiye sahiptir.
5.3 Anahtar çıkarımlar
Sentetik gen devreleri, kültürlenmiş et ekiplerine, yalnızca ortam protokollerinin eşleşemeyeceği zamanlama, eşik ve soy dengesi üzerinde programlanabilir kontrol sağlar. Mimari seçimi, geri döndürülebilirlik, sızıntı ve güvenliği şekillendirir.İndüklenebilir sistemler genellikle tercih edilir çünkü koşullu kontrol sağlarlar ve daha düşük metabolik yük sunarlar [6].
"Sentetik biyoloji araç seti, ayarlanabilir gen ekspresyonuna sahip hücre hatları oluşturmak için kullanılabilir ve bu, PAT ve hesaplamalı modelleme ile birleştirildiğinde, kapalı döngü kontrol sistemlerinin optimal ürün verimi ve kalitesi sağlamasına olanak tanır." - npj Systems Biology and Applications [6]
Başarılı uygulama sadece bir biyoloji sorunu değildir. Devre mühendisliği, biyoproses tasarımı, düzenleyici dokümantasyon ve tedarik arasında sıkı bir bağlantıya bağlıdır.
SSS
Sentetik gen devreleri farklılaşma tutarlılığını nasıl artırır?
Sentetik gen devreleri, hücre davranışı ve soy bağlılığı üzerinde programlanmış kontrol sağladıkları için farklılaşmayı daha tutarlı hale getirebilir.Pratikte, bu, gen ve transkripsiyon faktörü ifadesini sıkı bir zamanlama ile ayarlamak için modüler mantık işlemlerini kullanmak anlamına gelir.
Bu zamanlama önemlidir. Hücrelerin, karışık veya istenmeyen kaderlere sürüklenmek yerine, tanımlanmış durum değişikliklerinden doğru sırayla geçmesine yardımcı olur. Ayrıca, hedef dışı farklılaşmayı azaltır ve kültür boyunca gürültüyü düşürür.
Getirisi basittir: kültive edilmiş et üretimi için daha uniform, kararlı ve olgun hücre popülasyonları.
Hangi devre tasarımı miyojenik veya adipogenik kontrol için uygundur?
Kültive edilmiş et araştırmalarında, aynı tavuk fibroblastları her iki soy hattına da yönlendirilebilir. Miyogenez bir dizi indüksiyon protokolünü takip ederken, adipogenez hücrelerin tavuk serumu veya yağ asitleri gibi girdilere maruz bırakılmasıyla etkinleştirilebilir.
Buradan, bu hücre kaderleri, tanımlanmış yağ ve kolajen oranlarına sahip et yapıları oluşturmak için 3D hidrojel iskeleler içinde adım adım kontrol edilebilir.
Gen devreleri neden 3D kültürde genellikle farklı davranır?
3D kültürde, gen devreleri genellikle farklı davranır çünkü hücreler, 2D monolayerlerde basitçe var olmayan fiziksel ve yapısal girdilerle başa çıkmaktadır. Bu girdiler arasında mekanik gerilim, kayma stresi, matris sertliği ve yerel hücre yoğunluğu bulunur.
Bu ipuçları, Notch gibi sinyal yollarını değiştirebilir. Ayrıca, sentetik devrelerin kuvveti algılama ve hücre-hücre yapışması ve doku morfogenezini içeren aşağı akış tepkilerini koordine etme şeklini de değiştirebilirler.









