

השתקת אפיגנטיקה משנה את הדרך בה אנו ניגשים לייצור בשר מתורבת. עבור אנשי מקצוע ב-R&D, היא מציעה דרך לשלוט בביטוי גנים מבלי לשנות את ה-DNA לצמיתות, תוך התמודדות עם אתגרים מרכזיים כמו שגשוג תאים, התמיינות ובקרת איכות. הנה מה שאתם צריכים לדעת:
- מה זה: דיכוי פעילות גנים באמצעות מתילציה של DNA, שינוי היסטונים או התערבות RNA - שיטות הפיכות ומדויקות שמשאירות את רצף הגנים ללא שינוי.
- למה זה חשוב: מאריך את חיי התאים, משפר התמיינות תאי שריר ומשפר את יכולת ההרחבה תוך הימנעות מסיכונים כמו אונקוגנזה משינויים גנטיים קבועים.
- כלים מרכזיים: מערכות CRISPR-dCas9 (כמו KRAB או DNMT3A) ועורכים מבוססי TALE משיגים יעילות השתקה גבוהה, עם חלק מההשפעות הנמשכות מעל 300 ימים.
- אתגרים: אספקת הכלים הללו בקנה מידה רחב, במיוחד בביו-ריאקטורים, והתאמת גישות לנתיבים ספציפיים למינים נותרות מכשולים.
עבור מהנדסי ביופרוסס ומדעני תרביות תאים, המיקוד הוא בשליטה מדויקת על התנהגות התאים לשיפור הפרודוקטיביות ואיכות המוצר. השתקה אפיגנטית עשויה להיות המפתח להתגברות על החסמים בייצור בשר מתורבת.
מנגנונים מרכזיים של השתקה אפיגנטית בתאי בעלי חיים
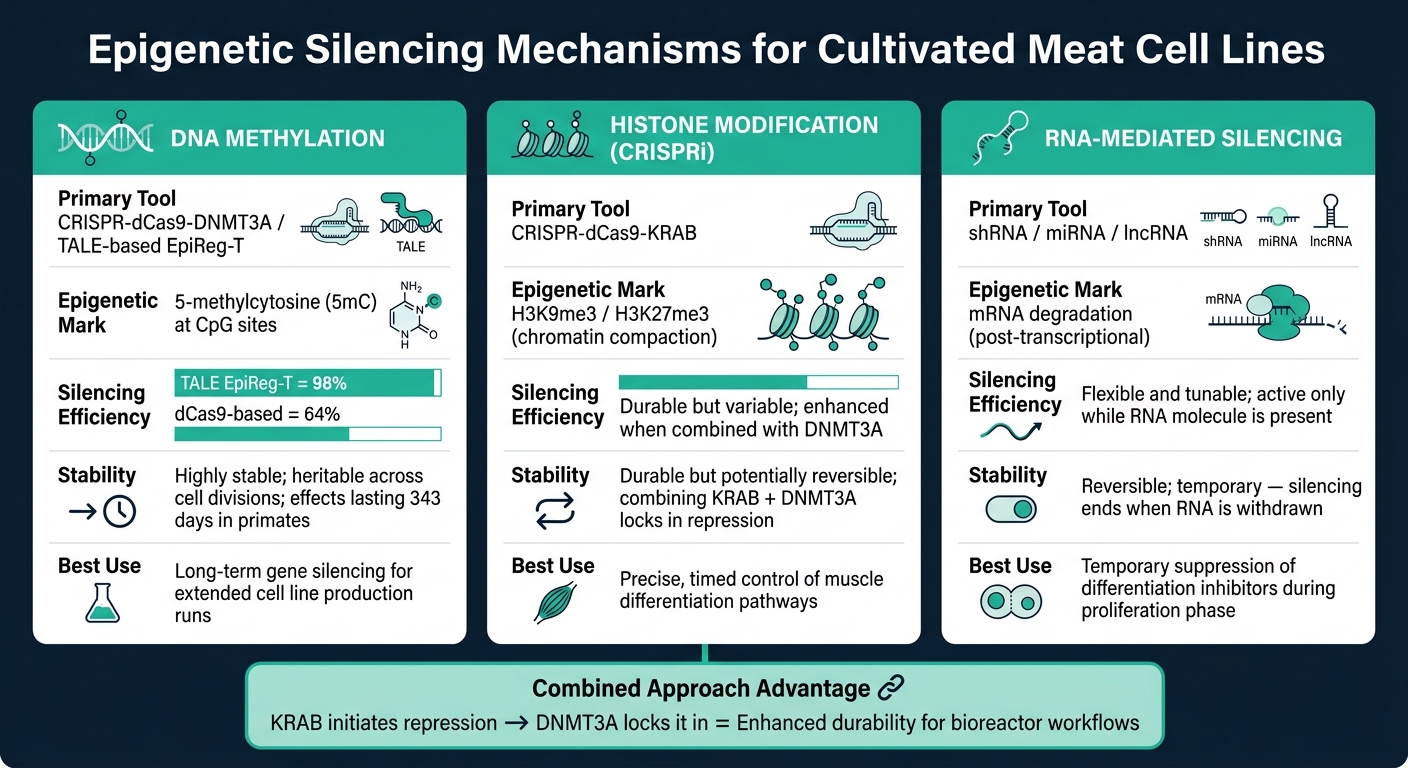
כלי השתקה אפיגנטיים לבשר מתורבת: מנגנונים, יעילות & יציבות
שיפור הביצועים של קווי תאים לבשר מתורבת תלוי במידה רבה בשליטה מדויקת על מנגנונים אפיגנטיים. להלן סקירה של השיטות העיקריות המשמשות בתאי בעלי חיים.
השתקה מבוססת מתילציה של DNA
מתילציה של DNA כוללת הוספת קבוצות מתיל לאתרי CpG, תהליך המונע על ידי DNA מתילטרנספראזות (DNMTs). כאשר זה מתרחש באזורים מקדמי גנים, זה מונע ממכונות השעתוק לגשת לגנים, ובכך מכבה אותם [6]. ההשתקה הזו ניתנת להעברה תורשתית, כאשר DNMT1 שומר על דפוס המתילציה לאורך חלוקות תאים [7].
כלי מתקדם אחד, CRISPR-dCas9-DNMT3A, משלב את החלבון dCas9 הלא פעיל קטליטית עם האנזים DNMT3A כדי להנחות מתילציה למיקומים גנומיים ספציפיים. שיטה זו משיגה יעילות השתקה גבוהה ללא חיתוך ה-DNA. גישה מעודנת יותר, רגולטורים אפיגנטיים מבוססי TALE (EpiReg-T), הראתה 98% יעילות השתקה בעכברים, בהשוואה ל-64% במערכות מבוססות dCas9 קודמות [5]. במחקרים עם פרימטים לא אנושיים, מנה אחת של מערכת זו שמרה על השתקת גנים למשך עד 343 ימים [5].
לאחר הקמת מתילציה של DNA, שינויים בהיסטונים מספקים שכבה משנית ודינמית של רגולציית גנים.
שינויי היסטונים ו-CRISPRi
שינויים בהיסטונים משנים את מבנה הכרומטין על ידי מיקוד לחלבוני היסטון, מה שהופך גנים לנגישים יותר או פחות. סימנים כמו H3K9me3 ו-H3K27me3 מקפלים את הכרומטין, מונעים מגורמי שעתוק גישה ל-DNA [6].
הפרעה באמצעות CRISPR (CRISPRi) משתמשת ב-dCas9 שמחובר לדומיין מדכא KRAB. קומפלקס זה מכוון למקדמי גנים ספציפיים, שם הוא מגייס חלבונים מדכאים שמניחים סימני היסטון מעכבים. מחקר בכבשים הדגיש את H3K27me3 כאות דיכוי מרכזי במהלך התפתחות השרירים, בעוד שמגברים פעילים מקושרים לגנים המקדמים ביצועי צמיחה מעולים [8]. על ידי הבנת מצבי ההיסטונים המווסתים את התמיינות השרירים בבעלי חיים, מדענים יכולים לכוון את התנהגות התאים בדיוק.
"עריכת אפיגנטיקה היא אסטרטגיה מבטיחה לשינוי ביטוי גנים תוך הימנעות משינויים קבועים ופוטנציאל גנוטוקסיות של טכנולוגיות עריכת גנום." - Nature Biotechnology [5]
שינויים בהיסטונים הם לעיתים קרובות דינמיים יותר מאשר מתילציה של DNA, ודורשים התערבויות מתמשכות או מתוזמנות כדי לשמור על השפעותיהם. שילוב KRAB עם DNMT3A במבנה יחיד יכול לשפר את העמידות: סימני היסטונים מתחילים דיכוי, בעוד שהמתילציה נועלת אותו במקום [5].
בנוסף לשיטות מבוססות DNA אלו, השתקה בתיווך RNA מציעה חלופה גמישה וזמנית.
השתקה בתיווך RNA
השתקה בתיווך RNA מתמקדת בהפחתת רמות ה-mRNA ישירות. מיקרו-רנ"א (miRNAs) ו-רנ"א קצר בצורת סיכת ראש (shRNAs) נקשרים לרצפי mRNA משלימים, מה שמוביל לפירוקם לפני התרגום [6] . בינתיים, רנ"א ארוך לא מקודד (lncRNAs) פועלים מוקדם יותר על ידי גיוס קומפלקסים המשנים כרומטין לאזורים גנומיים ספציפיים [6] .
ליישומים של בשר מתורבת, השתקה בתיווך RNA מציעה יתרון משמעותי: הפיכות וגמישות. ההשתקה נשארת פעילה רק כל עוד מולקולת ה-RNA נוכחת, מה שהופך אותה לאידיאלית להתערבויות זמניות. לדוגמה, ניתן לדכא מעכבי התמיינות במהלך שלב התרבות, ואז להסירם כדי לאפשר התפתחות שריר תקינה.עם זאת, שמירה על אספקה רציפה של מולקולות RNA יכולה להוסיף מורכבות כאשר מגדילים קווי תאים לגידול בביו-ריאקטור.
הטבלה למטה מסכמת את המאפיינים המרכזיים של המנגנונים הללו:
| מנגנון | כלי ראשי | סימון אפיגנטי | יציבות |
|---|---|---|---|
| מתילציה של DNA | CRISPR-dCas9-DNMT3A | 5-מתילציטוזין (5mC) | יציב מאוד; תורשתי בין חלוקות [5][7] |
| דיכוי היסטונים (CRISPRi) | CRISPR-dCas9-KRAB | H3K9me3 / H3K27me3 | עמיד אך פוטנציאלית הפיך [5][8] |
| הפרעת RNA | shRNA / miRNA | פירוק mRNA | הפיך וניתן לכוונון [6] |
sbb-itb-ffee270
גנים ומסלולים יעד לשיפור ביצועי קווי תאים
בהתבסס על דיונים קודמים על מנגנונים אפיגנטיים, בחירת גני היעד הנכונים היא קריטית לשיפור ביצועי קווי התאים.ההצלחה של התערבויות אלו תלויה לא רק בזיהוי המטרות אלא גם בבחירת שיטות ההשתקה המתאימות. מחקרים זיהו סט ליבה של מטרות גנים אשר, כאשר הם מדוכאים, משפרים היבטים מרכזיים של קווי תאי בשר מתורבת, כולל פרוליפרציה, התמיינות ויציבות מטבולית.
פרוליפרציה והאלמותיות
שיפור יכולת הפרוליפרציה כרוך לעיתים קרובות במיקוד גנים כמו CDKN2A ו-TP53. CDKN2A מקודד את p16^INK4A ו-p14^ARF, חלבונים שמגבילים את מחזור התא ומניעים הזדקנות. השתקת CDKN2A מונעת עצירת G1/S, ומאפשרת התרחבות תאית חזקה. לדוגמה, תאים חזיריים עם השתקת CDKN2A שמרו על תכונותיהם המיוגניות לאורך 18–30 מעברים, בעוד שתאים מסוג פראי איבדו תכונות אלו עד מעבר 10.יתרה מזאת, CDKN2A דלדול גרם לעלייה פי ~194 בביטוי של PAX7 במעבר 20, גורם קריטי לזהות תאי גזע שריר [9] .
"מיקוד בלוקוס הגן CDKN2A הוא חיוני למניעת הזדקנות או להשריית אלמוות תאי." - Food Materials Research [9]
TP53 הוא יעד מפתח נוסף. מסך CRISPR של 600 גנים בתאי גזע מזנכימליים של בקר זיהה את TP53 כיעד היעיל ביותר לשיפור פרוליפרציה. נוקאאוט של TP53 הביא לעלייה פי 1,000 בשפע התאים במשך 30 ימים, עם ביצועים עקביים בהתרחבות לטווח ארוך [1] . בנוסף, השתקת PTEN, רגולטור שלילי של מסלול PI3K/AKT/mTOR, מגבירה את שיעורי הכפלת התאים ואת פעילות mTOR.עם זאת, גישה זו דורשת ניטור קפדני, שכן היא עשויה להפחית את יעילות ההבחנה [1].
ההתקדמויות הללו בפרוליפרציה מכינות את הקרקע לאופטימיזציה של ההבחנה, השלב הקריטי הבא.
שליטה בהבחנה
איזון בין התרחבות תאים ליצירת רקמות הוא אתגר מורכב בייצור בשר מתורבת. מטרה שנחקרה היטב היא מיוסטטין (MSTN), רגולטור שלילי של מיוגנזה. השתקת MSTN משפרת את היווצרות סיבי השריר, בדומה לתכונת "שרירים כפולים" בגזעי בקר מסוימים [4] . כאשר משולב עם הפעלת MYOD1 וטכניקות מתקדמות כמו הדפסת תלת-ממד ביולוגית בעיבוד אור דיגיטלי (DLP) על הידרוג'לים עם דפוסי חריצים, יישור ותפקוד תאי שריר משתפרים משמעותית באמצעות פונקציונליזציה של פני השטח [4] .
היבט קריטי נוסף הוא ניהול רגולטורים של פלוריפוטנטיות כמו SOX2 ו-OCT4. השתקה הפיכה של SOX2 באמצעות פלטפורמות CRISPR/dCas9-KRAB משיגה עד 85% דיכוי תוך 72 שעות, כאשר הביטוי הבסיסי מתאושש לכ-90% לאחר הסרת המבנה העריכתי [3] . הפיכות זו מאפשרת דיכוי מבוקר במהלך התרחבות תאים ושחרור בזמן לתמיכה בהתפתחות רקמות תקינה.
מסלולי לחץ ומטבוליזם
שמירה על איכות התאים לאורך מחזורי ייצור ממושכים כוללת התמודדות עם אתגרי לחץ ומטבוליזם. TP53 ממלא תפקיד כפול כמדכא גידולים וחיישן לחץ. בתנאי תרבית, הוא יכול להפעיל הזדקנות מוקדמת, אפילו ללא נזק גנומי משמעותי [1]. על ידי השתקת TP53, התאים שומרים על פרופילי ביטוי הגנים של תאים במעבר מוקדם, ושומרים על פונקציות קריטיות כמו סינתזת חלבונים ושכפול DNA [1].
הטבלה למטה מסכמת את מטרות הגנים העיקריות ותפקידיהן הפונקציונליים:
| גן מטרה | מסלול | השפעת השתקה | הקשר מינים |
|---|---|---|---|
| CDKN2A | דיכוי מחזור התא | מונע הזדקנות; ~194× PAX7 עלייה בביטוי במעבר 20 [9] | חזיר |
| TP53 | תגובה ללחץ / מדכא גידולים | עלייה פי 1,000 בכמות התאים במשך 30 ימים; התרחבות עקבית לטווח ארוך [1] | בקר |
| PTEN | PI3K/AKT/mTOR | מגביר את קצב הכפלה; משפר את פעילות mTOR [1] | בקר |
| MSTN | ויסות מיוגנזה | משפר את היווצרות סיבי השריר ויעילות ההתמיינות [4] | בקר |
| SOX2 | שמירה על פלוריפוטנטיות | מנהל את המעבר ממצב גזע להתמיינות; דיכוי של 85% תוך 72 שעות [3] | רבים |
גישה מבטיחה שצוברת תאוצה היא מיקוד מרובה, הכוללת השתקה של מספר גנים בו זמנית.לדוגמה, שילוב של דיכוי CDKN2A עם הפעלת GATA4 הראה השפעות סינרגטיות העולות על התערבויות בודדות [9] [10] . אסטרטגיה ברמת המערכות הזו מדגישה את החשיבות של פלטפורמות מתמחות כמו
כלים אפיגנטיים ושיטות מסירה
כדי לנצל מטרות גנים ספציפיות, חוקרים מסתמכים על כלים אפיגנטיים מתמחים ומערכות מסירה יעילות.
פלטפורמות אפיגנטיות סינתטיות
זיהוי מטרות הגנים הנכונות הוא רק חלק מהמשוואה - הכלים המשמשים להשתקת גנים אלו הם קריטיים באותה מידה. שני מערכות מתוכנתות בולטות ברלוונטיות שלהן למחקר בבשר מתורבת: CRISPRoff ו-רגולטורים אפיגנטיים מבוססי TALE (EpiReg-T).
CRISPRoff משתמשת בשלד dCas9 בשילוב עם תחומי KRAB ו-DNMT3A/3L כדי ליצור סימנים מדכאים תורשתיים, כגון מתילציה של DNA ו-H3K9me3, מבלי להכניס שברים ב-DNA. גישה זו מבטיחה השתקה מתמשכת של גנים, מה שהופך אותה לשימושית במיוחד לשמירה על קווי תאים לאורך תקופות ממושכות - גורם מפתח בהתמודדות עם אתגרי ההרחבה והעקביות בייצור בשר מתורבת. לעומת זאת, EpiReg-T מבוסס TALE הראה יעילות השתקה עליונה, והגיע ל-98% בהשוואה ל-64% שנראו במערכות דומות מבוססות dCas9 [5] .
מחקר מכריע שפורסם ב-Nature Biotechnology באוקטובר 2025 הדגיש את הפוטנציאל של עורכים מבוססי TALE. חוקרים, כולל אלו מ-Epigenic Therapeutics ומה-האקדמיה הסינית למדעים, הראו כי מנה אחת של EpiReg-T שנמסרה באמצעות ליפיד ננו-חלקיקים (LNPs) השתקה את הגן PCSK9 בקופים עם יעילות של מעל 90% למשך 343 ימים . זה הושג עם השפעות מינימליות מחוץ למטרה, כפי שאושר באמצעות ניתוחים מולטי-אומיים [5]. תוצאות כאלה מציבות את המערכות מבוססות TALE בנפרד כאשר עמידות ועוצמה הן קריטיות.
אתגרי מסירה
מסירת הכלים הללו לתאי בעלי חיים - במיוחד בקנה מידה - נותרת אתגר טכני משמעותי. בעוד שעורכי אפיגנטיקה נמנעים מהסיכונים של שבירות DNA דו-גדיליות, הם עדיין דורשים מנגנון מסירה אמין. ליפיד ננו-חלקיקים (LNPs) הפכו לאפשרות הלא-ויראלית המובילה.הם מספקים באופן זמני mRNA המקודד את העורך האפיגנטי, ומאפשרים גישה של "פגע וברח" שמבססת השתקת גנים מתמשכת ללא אינטגרציה של DNA [5]. הטבע הזמני הזה חשוב במיוחד לבשר מתורבת, שבו חששות רגולטוריים סביב שינויים גנטיים נשארים נושא מרכזי.
עם זאת, היעילות של LNP יכולה להשתנות באופן משמעותי בהתאם לסוג התא. אופטימיזציה של פורמולציות עבור תאי מיוסאטליט ראשוניים של בקר או חזיר, במיוחד בהגדרות בקנה מידה של ביוריאקטור, היא עדיין תחום של מחקר פעיל. שיטות משלוח שעובדות היטב בניסויים בקנה מידה קטן לעיתים קרובות נכשלות בביצועים עקביים ב-ביוריאקטורים עם טנק מעורב. פתרון אתגרי המשלוח הללו חיוני לקידום המחקר ולהגדלת הייצור, תהליך הנתמך יותר ויותר על ידי פלטפורמות מתמחות.
כיצד Cellbase תומך באפיגנטיקה R&D
קווי תאים שעברו שינוי אפיגנטי דורשים ריאגנטים מאומתים בדיוק. חוקרים זקוקים לגישה לקווי תאים מאופיינים היטב שמתאימים לשינויים אפיגנטיים, פורמולציות מדיה מוגדרות ששומרות על יציבות אפיגנטית, וכלים אנליטיים שמסוגלים לאשר השתקת גנים ברמת הכרומטין. ספקי מעבדה כלליים לעיתים קרובות חסרים את המומחיות להבטיח תאימות ליישומי בשר מתורבת.
מה המשמעות של השתקה אפיגנטית עבור עיבוד ביולוגי של בשר מתורבת
שיפורים מדידים בקווי תאים
השתקה אפיגנטית מציעה יתרונות מעשיים שהופכים ליותר ויותר ברורים, במיוחד בהארכת תוחלת החיים היצרנית של קווי תאים. על ידי שימוש באסטרטגיה זמנית של "פגע וברח", חוקרים יכולים לדכא באופן זמני גנים האחראים להזדקנות מבלי לשנות לצמיתות את הגנום [2]. גישה זו הראתה הצלחה ב-תאי מיוסאטליטים של בקר ו-תאי מיוסאטליטים של חזיר, מאפשרת הכפלות תאים רבות יותר באופן משמעותי ומטפלת בצווארי בקבוק נפוצים בעיבוד ביולוגי.חשוב לציין, ששיטה זו הפיכה - ברגע שהמבנה מוסר, הביטוי הגנטי כמעט חוזר לרמות הבסיס [3]. שליטה הפיכה זו אידיאלית עבור תהליכי עבודה בביו-ריאקטור, שכן היא מבטיחה שהתאים ימשיכו להתרבות במהלך שלב ההתרחבות ומאפשרת להפעיל את ההתמיינות בזמן המתאים. הרחבת תאים משופרת מתורגמת ישירות להתמיינות רקמות יעילה יותר ולשיפור איכות המוצר.
יצירת רקמות ואיכות המוצר
ההישגים בהתרבות התאים יוצרים את הבסיס ליצירת רקמות טובה יותר. התמיינות מבוקרת היא המקום שבו השתקה אפיגנטית משפיעה ישירות על איכות המוצר הסופי. לדוגמה, בתכנות מחדש של תאי בקר, השתקת סמני פלוריפוטנטיות כמו OCT4, SOX2, ו-NANOG מקלה על המעבר לשושלת המיוגנית.תהליך זה מוביל ליצירת מיוטיוביות מוארכות ורב-גרעיניות עד יום 30 של פרוטוקול ההתמיינות [11].
"השתקת mOSKM וסמני פלוריפוטנטיות... היא קריטית למעבר מפלוריפוטנטיות לקו המיוגני." - Frontiers in Nutrition [11]
מעבר לפיתוח סיבי שריר, שליטה אפיגנטית מדויקת על מסלולי התמיינות תאי שומן משחקת תפקיד קריטי בהשגת שיש. שיש הוא גורם מפתח שמשפיע על הטעם והמרקם, ושיפורים אלו יכולים להיות מושגים ללא ביצוע שינויים קבועים בגנום.
שיקולים רגולטוריים וצרכניים
ההתקדמות בהתרבות תאים וביצירת רקמות גם מביאה את הפרספקטיבות הרגולטוריות והצרכניות למוקד.גופים רגולטוריים בדרך כלל תומכים בהשתקה אפיגנטית בשל השפעתה הלא-קבועה על הגנום. כלים כמו dCas9-KRAB ו-EpiReg-T מבוסס TALE נמנעים מהסיכונים הקשורים לשבירות DNA דו-גדיליות, מה שהופך אותם למתאימים לקווי תאים בדרגת מזון שחייבים להראות יציבות גנטית לאורך כל הייצור [5].
עם זאת, שמירה על מצב ללא גנים זרים נותרת אתגר. מחקר שפורסם במאי 2025 על ידי חוקרים מ-אוניברסיטת סאו פאולו ו-אוניברסיטת קופנהגן, כולל קיאנה רקיה וקריסטין פרוידה, חקר את הנושא הזה. הם תכנתו מחדש פיברובלסטים עובריים של בקר באמצעות וקטורים אפיזומליים לא-משלבים, ומצאו שבעוד שהמושבות נותרו יציבות ליותר מ-33 מעברים, פלסמידים אפיזומליים עדיין היו ניתנים לזיהוי במעברים 12 ו-17 [11].
מצד הצרכן, שקיפות לגבי השיטות שבהן נעשה שימוש היא קריטית. תקשורת ברורה שהשתקת אפיגנטית אינה משנה את ה-DNA לצמיתות תהיה מפתח לבניית אמון הציבור ככל שמוצרים של בשר מתורבת מתקרבים למסחור.
כיוונים עתידיים ופערי מחקר
אתגרים ספציפיים למינים
אחד המכשולים הגדולים בתחום הוא חוסר הבנה מפורטת של מסלולים מיוגניים במיני בעלי חיים. בעוד שמסלולים כמו IGF-1, MAPK/Erk, ו-Wnt/β-catenin מתועדים היטב בבני אדם ועכברים, תפקידיהם בבקר וחזירים מובנים רק באופן חלקי [11]. ללא מפה מלאה, זיהוי מטרות גנים ספציפיות להשתקה אפיגנטית הופך לאתגר משמעותי.
הרכב סיבי השריר מוסיף שכבת מורכבות נוספת. לדוגמה, שריר Longissimus של חזיר מכיל כ-55% סיבי Type IIb מהירים, אך סיבים אלו נעדרים במינים כמו כבשים וסוסים.כאשר משלבים זאת עם ביטוי גנים HOX ספציפי לאזור, מתברר כי יש להתאים אסטרטגיות השתקה לכל מין [13]. תאי לוויין, השומרים על ביטוי גנים HOX לפי מיקום (e.g. , HOXA11 ו-HOXA13 בשרירי הגפיים האחוריות), מסבכים את העניינים עוד יותר. דפוסים אלה יכולים להשפיע על נטיית התאים לפרוליפרציה מהירה או לדיפרנציאציה חזקה [14].
"מכיוון שתאי SC יכולים לשמור על חתימות מיקום אלו, יכולות הפרוליפרציה והדיפרנציאציה שלהם עשויות להשתנות לפי מקור השריר." - npj Science of Food [14]
במונחים מעשיים, משמעות הדבר היא שחוקרים צריכים לבדוק קווי תאים לביטוי גנים HOX לפני יישום השתקה אפיגנטית.חתימות גנים אלו יכולות לפעול כברקודים ביולוגיים, ולעזור לאמת את הזהות האזורית של תאים וליישר אותם עם המאפיינים הרצויים של המוצר הסופי.
אתגרים ספציפיים למינים אלו מדגישים את החשיבות של שקילת מקורות תאים חלופיים, כגון iPSCs, בפיתוח אסטרטגיות לבנקאות תאים.
קישורים לפיתוח iPSC ובנקאות תאים
תאי גזע פלוריפוטנטיים מושרים (iPSCs) מציעים חלופה מבטיחה לתאי לוויין, הנוטים להזדקנות ודורשים ביופסיות חוזרות. במאי 2025, חוקרים מאוניברסיטת סאו פאולו ואוניברסיטת קופנהגן - כולל קיאנה רקיה וקריסטין פרוידה - פיתחו בהצלחה קווי iPSC של בקר באמצעות וקטורים אפיזומליים לא משולבים. תאים אלו שמרו על יציבות במשך יותר מ-33 מעברים והתמיינו למיוטיובים רב-גרעיניים עד יום 30 [11]. עם זאת, אישור הסטטוס שלהם ללא טרנסגן באמצעות PCR גנומי קפדני נותר שלב קריטי.
נושא קשור הוא זיכרון אפיגנטי . תאי iPSCs לעיתים קרובות שומרים על עקבות של רקמת הסומטית המקורית שלהם, מה שיכול להטות את ההתמיינות מהשושלת המיועדת [12]. לצורך בנקאות תאים, חשוב לבחור רקמות תורם עם פרופילים אפיגנטיים שכבר מכוונים ליצירת שריר או שומן. בנוסף, הבטחת השתקה אפקטיבית של סמני פלוריפוטנטיות שאריים היא חיונית ליצירת בנקי תאים אמינים וארוכי טווח.
פיתוח פרוטוקולים חזקים ל-iPSC גם מדגיש את הצורך בבדיקות סטנדרטיות ופרקטיקות עקביות לשיתוף נתונים בין מאמצי מחקר.
סטנדרטיזציה ונתונים חסרים
כדי לנצל במלואו את הפוטנציאל של התערבויות אפיגנטיות בבשר מתורבת, יש לטפל בבעיות הסטנדרטיזציה. נכון לעכשיו, אין מסגרת אוניברסלית לניטור יציבות אפיגנטית במהלך הכפלת תאים נרחבת הנדרשת לייצור בקנה מידה תעשייתי [12]. ללא שיטות סטנדרטיות, קשה להשוות תוצאות בין מעבדות, והחלטות על הגדלת הייצור לעיתים קרובות מסתמכות על נתונים לא שלמים.
צעדים מעשיים יכולים לעזור להתמודד עם הפער הזה. לדוגמה, אימוץ פרוטוקולי טיהור FACS עקביים - המכוונים לסמנים כמו CD31⁻/CD45⁻/CD29⁺/CD56⁺ - יהפוך את אוכלוסיות תאי הלוויין ליותר ברות השוואה בין מינים ומקורות אנטומיים [14]. מעבר ממדיה מבוססת סרום למדיה מוגדרת כימית יכול גם להפחית את השונות בין אצוות, וליצור סביבות אפיגנטיות עקביות יותר [12].
מבט לעתיד, שילוב מודלים in silico מונעי AI יכול לחולל מהפכה באופטימיזציה של פרוטוקולים אפיגנטיים.עם זאת, כדי שמודלים אלה יהיו יעילים, חיוני לתאם נתונים בקהילת המחקר של בשר מתורבת. פרקטיקות שיתוף נתונים סטנדרטיות יאפשרו לחוקרים לחזות את תוצאות המניפולציות האפיגנטיות בצורה מדויקת יותר, ולהאיץ את ההתקדמות בתחום.
שאלות נפוצות
כיצד השתקה אפיגנטית שונה מעריכת גנים קבועה בתאי בשר מתורבת?
השתקה אפיגנטית מווסתת את פעילות הגנים מבלי לבצע שינויים קבועים ברצף ה-DNA, בניגוד לעריכת גנים, הכוללת שינוי פיזי של הגנום. מכיוון שגישות אפיגנטיות אינן כוללות שבירה או שינוי של ה-DNA, הן נחשבות לעיתים קרובות כאופציות בטוחות יותר לשימוש בייצור בשר מתורבת. טכניקות כמו כלים מבוססי CRISPR מציעות את היתרון של ויסות גנים גמיש ובמקרים מסוימים הפיך.עבור חוקרים העובדים עם שיטות אלו,
אילו גנים יש להשתיק תחילה כדי להגביר את הפרוליפרציה מבלי לפגוע בהתמיינות?
כדי לעודד פרוליפרציה של תאים תוך שמירה על יכולתם להתמיין, חשוב להשתיק גנים שחוסמים את מחזור התא או מובילים לגורלות תאים לא רצויים. לדוגמה, דיכוי CDKN2A הוכח כמגביר באופן משמעותי את הפרוליפרציה בתאי לוויין של חזירים מבלי לפגוע בפוטנציאל ההתמיינות שלהם. באופן דומה, מיקוד בגנים מדכאי גידולים כמו TP53 ו-PTEN יכול לשפר את הצמיחה, אם כי התערבויות אלו דורשות פיקוח קפדני.
כיצד ניתן לספק עורכי אפיגנטיקה באופן אמין בקנה מידה של ביוריאקטור?
אספקת עורכי אפיגנטיקה בקנה מידה גדול לייצור בשר מתורבת מציבה אתגר משמעותי. זאת בעיקר בשל הגודל המשמעותי של כלי CRISPR והמגבלות של שיטות אספקה קונבנציונליות כמו אלקטרופורציה או וקטורים ויראליים. עם זאת, כמה אסטרטגיות מבטיחות מתפתחות. לדוגמה, מערכות אספקה זמניות המשתמשות בננו-חלקיקים שומניים או חלקיקים דמויי וירוס מהונדסים מראות פוטנציאל. שיטות אלו יכולות לעטוף מטעני CRISPR גדולים, ולאפשר כניסה יעילה לתאים מבלי לגרום לאינטגרציה גנומית. כדי לתמוך ביוזמות מתקדמות כאלה,









