

Jika Anda dapat memperluas sel tetapi tidak dapat mengubahnya ke nasib yang tepat pada waktu yang tepat, proses Anda akan terhenti pada diferensiasi. Itulah inti dari poin ini: sirkuit gen sintetis memberi Anda kontrol di dalam sel atas komitmen, waktu, memori, dan campuran garis keturunan, di mana perubahan media saja sering meninggalkan populasi heterogen, yang sebagian berkomitmen.
Jika saya membangun alur kerja diferensiasi daging yang dibudidayakan, saya akan mengambil empat poin dari artikel ini segera:
- Mulailah dengan jaringan asli, bukan konstruksi. Gunakan analisis lintasan snRNA-seq, , inferensi GRN, dan profil miRNA untuk menemukan di mana sel terhenti, melayang, atau bercabang ke nasib yang salah.
-
Cocokkan jenis sirkuit dengan masalah proses.
Sakelar toggle switch cocok untuk penguncian, desain feedforward atau band-pass cocok untuk kontrol waktu, gerbang logika logic gate cocok untuk pengaturan multi-sinyal, dan miSFITs cocok untuk keluaran bertingkat. - Desain untuk kebocoran rendah, kebisingan rendah, dan keamanan sejak hari pertama. Bagian ortogonal, autoregulasi negatif, iFFLs, transgen cm, dan modul pembunuhan atau penahanan pertumbuhan yang dapat diinduksi adalah bagian dari pembangunan, bukan pemikiran setelahnya.
- Validasi di bawah kondisi yang relevan dengan skala lebih awal. Sirkuit yang bekerja dalam 2D dapat berubah dalam 3D, microcarrier atau suspensi yang diaduk karena gradien inducer, batas oksigen, dan gesekan.
Artikel ini juga membuat poin praktis yang penting bagi tim proses: kontrol garis keturunan tunggal dan kontrol rasio adalah pekerjaan yang berbeda. Sebuah kaset Tet-On MyoD mungkin mendorong masuknya miogenik, tetapi produk potong utuh memerlukan kontrol proporsi otot, lemak, dan ECM, yang biasanya berarti umpan balik, sinyal parakrin, dan penyaringan klon yang lebih ketat.
Beberapa angka yang mendukung pesan tersebut:
- Diferensiasi myogenik standar dapat terhenti dengan indeks fusi sekitar 50–60%
- GRN yang direkayasa dalam iPSC meningkatkan diferensiasi garis keturunan target dari 52% menjadi 81%
- Sirkuit sintetis dalam MSC yang dimodifikasi mendorong diferensiasi jantung hingga 76%
- Beberapa garis Tet-On-PAX7 babi mempertahankan potensi myogenik tinggi lebih dari 40 kali pasase
- Sekitar 20% sel induk pluripoten manusia mungkin membawa mutasi yang terkait dengan kanker, itulah sebabnya modul keamanan yang dapat diinduksi penting
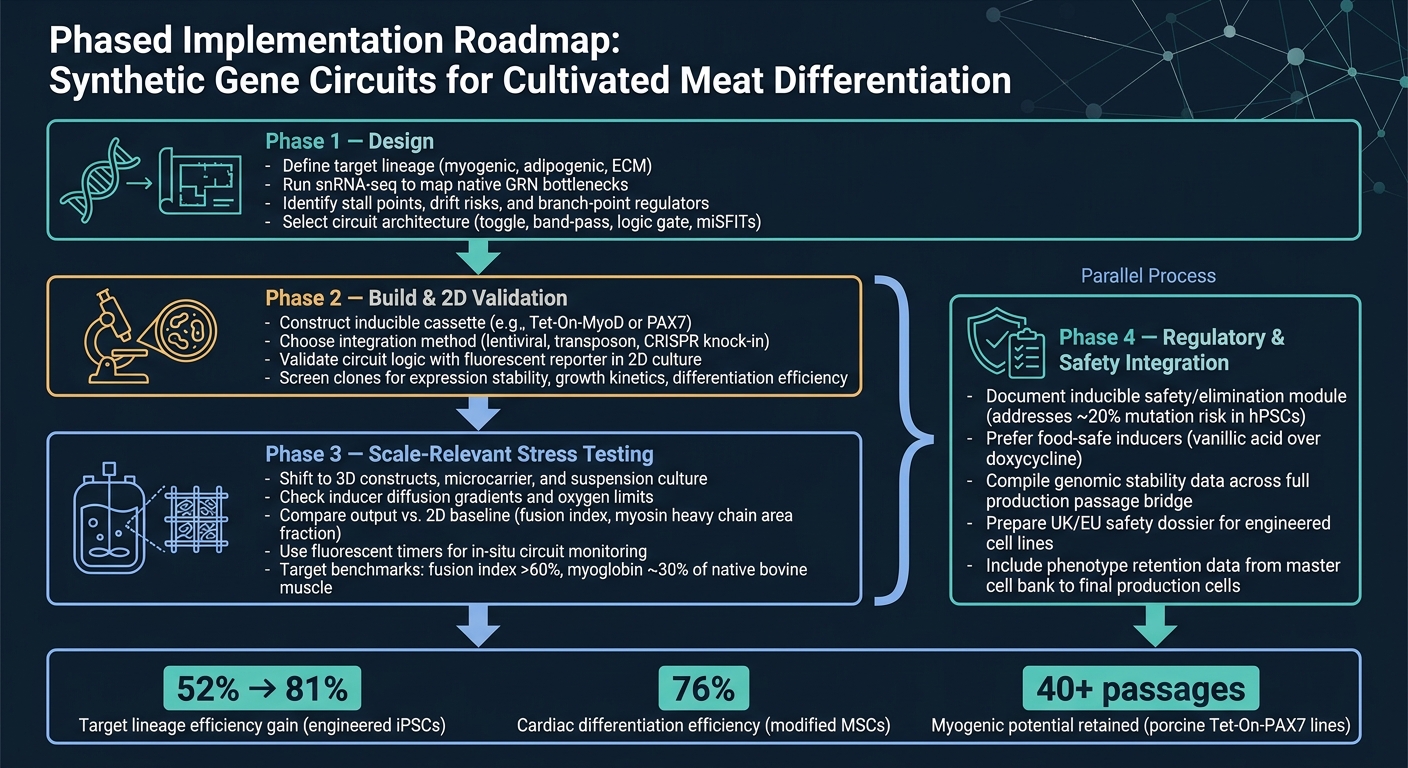
Alur Kerja Sirkuit Gen Sintetis untuk Diferensiasi Daging Budidaya
Kisah Ilmuwan: Michael Elowitz, Sirkuit Genetik dalam Sel Hidup
Perbandingan cepat
| Topik | Apa yang dikatakan artikel dalam istilah sederhana |
|---|---|
| Hambatan utama | Diferensiasi, bukan ekspansi |
| Mengapa sirkuit membantu | Mereka menambahkan kontrol ambang dan memori keadaan sel di dalam sel |
| Langkah pertama terbaik | Peta kontrol nasib asli dengan snRNA-seq dan alat terkait |
| Opsi sirkuit utama | Toggle, feedforward, band-pass, gerbang logika, miSFITs, CRISPRa/i |
| Contoh miogenesis | Tet-On-MyoD untuk memisahkan pertumbuhan dari diferensiasi terminal |
| Adipogenesis / kontrol ECM | miSFITs dan desain bow-tie untuk keluaran tipe PPARG/BMP4 bertingkat |
| Tantangan whole-cut | Kontrol rasio di antara otot, lemak, dan jaringan ikat |
| Risiko peningkatan skala | Perilaku 2D mungkin tidak berlaku dalam 3D atau bioreaktor |
| Pilihan integrasi | Lentivirus, transposon, CRISPR knock-in, vektor episomal |
| Titik regulasi | Garis yang direkayasa memerlukan paket keamanan yang lebih luas; inducer yang aman untuk makanan seperti asam vanilat lebih disukai daripada DOX jika memungkinkan |
Jadi, dalam istilah sederhana: ini bukan hanya artikel desain sirkuit. Saya akan membacanya sebagai panduan untuk menghubungkan arsitektur sirkuit, biologi garis keturunan, pemilihan klon, kinerja bioreaktor, dan dokumentasi keselamatan UK/EU ke dalam satu strategi diferensiasi.
Baca terus jika Anda ingin jalur lengkap dari pemetaan jaringan asli hingga pilihan konstruksi, pemeriksaan peningkatan skala, dan kesesuaian regulasi.
2. Prinsip desain untuk sirkuit kontrol diferensiasi
2.1 Peta jaringan nasib sel asli sebelum merancang sirkuit
Sebelum Anda merancang sirkuit, Anda memerlukan gambaran yang jelas tentang apa yang sudah dilakukan sel.
Single-nucleus RNA sequencing (snRNA-seq) adalah tempat yang baik untuk memulai. Ini dapat menunjukkan subpopulasi quiescent, termasuk sel cadangan yang ditandai oleh NOTCH2 dan HEYL , dan menunjukkan target jalur yang dapat meningkatkan diferensiasi [3].
Dari sana, analisis trajektori dan inferensi jaringan regulasi gen (GRN) membantu memetakan urutan aktivasi regulator dan menyoroti di mana sel kemungkinan besar akan terhenti. Dalam miogenesis, kaskade utama berjalan melalui MYOD1 dan MYOG . Dalam adipogenesis, node utama adalah PPARG dan CEBPA , dengan nasib progenitor fibro-adipogenik (FAP) bertindak sebagai titik cabang utama risiko. Tabel di bawah ini merangkum regulator utama dan hambatan.
| Garis Keturunan | Regulator utama kunci | Jalur sinyal kritis | Hambatan yang teridentifikasi |
|---|---|---|---|
| Miogenik | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Pembentukan sel cadangan (kekosongan) |
| Adipogenik | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Nasib progenitor fibro-adipogenik (FAP) |
| Pluripoten | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Diferensiasi spontan / heterogenitas |
Lapisan berguna lainnya adalah profil ekspresi miRNA. miRNA endogen seperti miR-302a, terkait dengan pluripotensi, dan miR-375, terkait dengan diferensiasi, dapat bertindak sebagai pengklasifikasi internal dalam desain sense-and-respond.Yang memungkinkan sirkuit membaca keadaan sebenarnya dari sel alih-alih hanya bergantung pada induktor eksternal [5].
Bottleneck tersebut harus mendorong pilihan sirkuit. Jika masalah utamanya adalah drift, Anda mungkin memerlukan lock-in. Jika masalahnya adalah waktu, pulsa mungkin lebih cocok. Jika kontrol nasib bergantung pada lebih dari satu isyarat, logika multi-input biasanya lebih masuk akal.
2.2 Pilih arsitektur sirkuit yang tepat
Di sinilah trade-off muncul dengan jelas. Arsitektur yang tepat bergantung pada tiga poin praktis: seberapa permanen komitmen harus, seberapa ketat waktu perlu dikendalikan, dan seberapa banyak muatan genetik yang dapat dibawa sel tanpa masalah.
Saklar toggle bistabil cocok untuk kasus di mana komitmen garis keturunan perlu tetap terkunci. Masalah utamanya adalah flipping keadaan spontan yang disebabkan oleh kebisingan transkripsi.
Filter band-pass cocok untuk kasus di mana faktor transkripsi hanya dibutuhkan dalam jendela perkembangan tertentu. Tantangannya adalah bahwa tingkat inducer perlu dikendalikan dengan ketat, atau waktu bisa meleset.
Gerbang logika menambah spesifisitas dengan memerlukan lebih dari satu input pada saat yang sama. Misalnya, diferensiasi hanya dapat diizinkan ketika ada inducer eksogen dan sel menunjukkan profil miRNA endogen yang tepat. Itu membantu mengurangi risiko komitmen yang salah sasaran.
Tabel di bawah ini menguraikan arsitektur utama dan kompromi yang menyertainya.
| Arsitektur | Reversibilitas | Presisi temporal | Kompleksitas integrasi | Kasus penggunaan utama | Risiko utama |
|---|---|---|---|---|---|
| Saklar bistabil | Rendah (setelah terkunci) | Sedang | Sedang | Komitmen garis keturunan permanen | Pembalikan spontan karena kebisingan |
| Filter pita lebar | Tinggi (tergantung konsentrasi) | Tinggi | Tinggi | Tahap perkembangan sementara | Membutuhkan kontrol inducer yang ketat |
| Gerbang logika (AND/OR/NOT) | Variabel | Sedang | Sedang–tinggi | Aktivasi spesifik tipe sel | Kebocoran dalam keadaan OFF |
| Dasi kupu-kupu / multi-input | Tinggi | Sedang | Sedang | Integrasi multi-sinyal | Bergantung pada stabilitas miRNA endogen |
| miSFITs | Tinggi | Sedang | Rendah–sedang | Penyetelan keluaran bertingkat | Rentang dinamis sempit jika penyetelan buruk |
"Dengan meminimalkan jumlah lapisan komputasi sambil mempertahankan fungsionalitas, strategi ini mengatasi hambatan skalabilitas dalam rekayasa sirkuit gen." - Nature Communications [9]
Setiap lapisan regulasi tambahan meningkatkan beban dan menggunakan sumber daya seluler. Dalam praktiknya, jika dua desain melakukan pekerjaan yang sama, yang lebih sederhana biasanya menjadi pilihan yang lebih baik ketika skala menjadi penting.
Setelah arsitektur ditetapkan, pekerjaan berikutnya adalah membuatnya bertahan di bawah kebocoran rendah, penekanan kebisingan, dan kontrol aman.
2.3 Bangun untuk keandalan, kebocoran rendah, dan keamanan
Sebuah sirkuit harus tetap stabil selama kultur yang diperpanjang. Ledakan kinerja yang baik dalam waktu singkat tidak cukup untuk penggunaan produksi.
Bagian ortogonal adalah pertahanan pertama. Promotor, faktor transkripsi, dan elemen regulasi yang tidak bereaksi silang dengan mesin asli membantu membatasi efek di luar target dan mengurangi kemungkinan sinyal endogen akan mengaktifkan atau mematikan sirkuit."Promotor dengan keketatan tinggi yang dimodifikasi seperti PCREm telah digunakan untuk mengurangi kebocoran basal dalam sistem mamalia yang dapat diinduksi [6] .
Autoregulasi negatif juga layak ditambahkan jika memungkinkan. Ini adalah salah satu motif yang lebih dikenal untuk mengurangi kebisingan transkripsi dan menghasilkan respons yang lebih linear terhadap konsentrasi inducer [6] . Loop umpan maju tidak koheren (iFFLs) dapat menambahkan lapisan kontrol lain dengan menyaring fluktuasi stokastik, sehingga sel merespons sinyal yang berkelanjutan daripada lonjakan kebisingan singkat.
Versi faktor transkripsi sintetis yang dimodifikasi kodon ( cm) juga memudahkan karakterisasi. Mereka memungkinkan Anda memisahkan ekspresi yang digerakkan oleh sirkuit dari ekspresi genom endogen (g) selama validasi [1]. Itu mungkin terdengar seperti detail kecil, tetapi itu menghemat waktu ketika Anda mencoba menentukan apakah pembacaan berasal dari sirkuit atau genom inang.
Modul keamanan diperlukan. Sekitar 20% dari sel induk pluripoten manusia membawa mutasi yang terkait dengan kanker [7] . Jadi, jika sebuah sirkuit akan dimasukkan ke dalam garis yang berasal dari sel induk, itu harus mencakup modul penghentian pertumbuhan atau eliminasi yang dapat diinduksi. Asam vanilat adalah inducer yang berguna untuk diprioritaskan di sini karena ini adalah aditif makanan berlisensi, yang membantu kasus penggunaannya sebagai pemicu sirkuit dalam garis sel daging yang dibudidayakan [1] .
"Biologi sintetis menyediakan insinyur dengan sarana untuk menerapkan sirkuit untuk dengan mudah dan tepat menyetel ekspresi beberapa gen untuk... menghilangkan potensi efek samping off-target yang merugikan." - npj Systems Biology and Applications [6]
Pilihan-pilihan ini menetapkan sirkuit spesifik garis keturunan di Bagian 3.
sbb-itb-ffee270
3. Strategi sirkuit untuk diferensiasi miogenik, adipogenik, dan terkontrol rasio
3.1 Sirkuit miogenik yang memisahkan ekspansi dari diferensiasi terminal
Setelah arsitektur sirkuit ditetapkan, tugas berikutnya adalah penerapan spesifik garis keturunan. Untuk miogenesis, masalah utamanya mudah untuk dinyatakan tetapi sulit untuk dilaksanakan: sel perlu berkembang biak terlebih dahulu, kemudian beralih ke diferensiasi ketika diminta, tanpa menyimpang terlalu awal ke arah mana pun.
Sebuah Tet-On-MyoD cassette adalah salah satu cara paling langsung untuk melakukan ini. Dalam pengaturan ini, sel berkembang biak di bawah kondisi standar ketika doksisiklin (DOX) tidak ada. Tambahkan DOX, dan sirkuit mendorong komitmen miogenik.Para peneliti di beberapa institusi Tiongkok menggunakan pendekatan ini pada fibroblas embrio ayam dan melaporkan pembentukan miotube yang efisien setelah induksi [4] .
Sebuah bistable toggle switch menawarkan kontrol keadaan yang lebih ketat. Sistem yang dibangun dari penekan yang saling menghambat seperti E-KRAB dan Pip-KRAB dapat mengubah pulsa DOX singkat menjadi program miogenik yang stabil [6] . Dalam praktiknya, ini berarti keadaan terdiferensiasi tetap terkunci setelah induksi daripada memudar setelah sinyal dihapus. Menambahkan autoregulasi negatif juga membantu mengurangi kebisingan stokastik dan membatasi diferensiasi bocor selama fase ekspansi [6].
Ini penting karena diferensiasi miogenik in vitro standar sering terhenti di tengah jalan. Indeks fusi biasanya hanya sekitar 50–60%, yang meninggalkan populasi cadangan yang tidak menyatu dalam jumlah besar [3]. Aktivasi MyoD yang digerakkan oleh sirkuit dapat meningkatkan komitmen, tetapi komitmen saja tidak menjamin fusi yang seragam. Ketika sirkuit MyoD digabungkan dengan modulasi MEK, NOTCH, dan RXR, fusi yang hampir lengkap dalam kultur 2D menjadi mungkin [3]. Sirkuit mengontrol waktu masuk garis keturunan; molekul kecil mendorong keluar siklus sel yang lebih bersih dan lebih seragam.
3.2 Kontrol adipogenik dan fibrogenik untuk komposisi dan struktur
Otot saja tidak cukup. Setelah kontrol miogenik diterapkan, masalah berikutnya adalah komposisi: berapa banyak lemak yang terbentuk, berapa banyak ECM yang disimpan, dan kapan program-program tersebut diaktifkan. Di sini, kontrol ON/OFF biner seringkali terlalu kasar. Yang biasanya dibutuhkan tim adalah keluaran yang bertingkat, terutama di sekitar titik cabang adipogenik-fibrogenik.
miSFITs memberikan cara praktis untuk menyesuaikan ekspresi secara bertahap.Dengan menempatkan situs target miRNA yang bermutasi - misalnya, situs untuk miR-17 - di 3′UTR dari gen output seperti PPARG atau BMP4, peneliti dapat memilih tingkat ekspresi dari perpustakaan varian [5]. Hal ini membuat induksi adiposit lebih mirip dengan saklar dimmer daripada saklar lampu. Alih-alih mendorong sel ke dalam respons semua-atau-tidak sama sekali, tim dapat menitrasi adipogenesis dengan lebih hati-hati [5].
Fibroblas juga bukan hanya pengamat di sini. Mereka menyediakan protein ECM yang membentuk tekstur [10]. Hal ini membuat kontrol fibrogenik menjadi bagian dari desain produk, bukan hanya masalah sampingan. Sirkuit dapat membantu mengelola pergeseran antara keadaan fibrogenik dan adipogenik, dan dalam unggas yang dibudidayakan, aktivasi PPARG langsung pada fibroblas mungkin diperlukan untuk menghasilkan deposisi lemak yang berarti [10].
Arsitektur bow-tie cocok untuk masalah ini karena menjaga pemisahan antara sensing dan output. Lapisan sensing membaca keadaan sel saat ini, sementara lapisan output menyesuaikan PPARG, CEBPA, atau regulator garis keturunan lainnya. Pemisahan tersebut membantu mencegah program adipogenik atau jaringan ikat menyala sebelum sel mencapai tahap perkembangan yang tepat.
3.3 Kontrol rasio multi-garis keturunan dan sensing umpan balik
Sirkuit kontrol rasio menangani masalah yang berbeda: bukan apakah diferensiasi terjadi, tetapi apakah campuran populasi akhir tetap di tempat yang seharusnya. Untuk produk whole-cut, mendapatkan otot, lemak, dan ECM dalam proporsi yang tepat sama pentingnya dengan mendapatkan satu garis keturunan untuk berdiferensiasi.
Sistem ini membangun kontrol umpan balik ke dalam sel itu sendiri. Promotor spesifik keadaan membatasi ekspresi protein sinyal ke sel yang telah berkomitmen pada garis keturunan tertentu.Modul parakrin sintetis kemudian memungkinkan sel miogenik yang berkomitmen melepaskan sinyal penghambat yang menekan komitmen adipogenik pada sel di sekitarnya. Logikanya mirip dengan inhibisi lateral dalam sistem perkembangan Delta-Notch [1] [6]. Di mana percabangan menjadi lebih rumit, gerbang multi-input dapat menggabungkan isyarat ekstraseluler dengan sinyal keadaan internal [9].
miSFITs juga bekerja pada tahap ini. Dengan menyetel kekuatan keluaran BMP4 atau morfogen lainnya, tim dapat menggeser keseimbangan garis keturunan tanpa menulis ulang logika keputusan hulu. Dalam jaringan pengaturan gen yang direkayasa, jenis kontrol ini meningkatkan efisiensi diferensiasi garis keturunan target dari 52% dalam kontrol menjadi 81% dalam iPSC yang direkayasa [2]. Dalam sel punca mesenkimal yang dimodifikasi, sirkuit sintetis mendorong efisiensi diferensiasi jantung hingga 76% [2].
Tabel di bawah ini membandingkan pendekatan single-lineage dan ratio-control menggunakan poin-poin yang paling penting dalam produksi.
| Fitur | Sirkuit garis keturunan tunggal | Sirkuit kontrol rasio |
|---|---|---|
| Kompleksitas | Rendah; biasanya satu promotor dan regulator yang dapat diinduksi [4] | Tinggi; memerlukan gerbang logika dan sinyal parakrin [6][9] |
| Beban pemantauan | Rendah; biasanya mengikuti satu pelapor [4] | Tinggi; memerlukan pelacakan beberapa penanda garis keturunan [5] |
| Ketahanan | Sedang; rentan terhadap heterogenitas dan pembentukan sel cadangan [3] | Tinggi; menggunakan umpan balik dan inhibisi lateral untuk menyeimbangkan populasi [1] |
| Nilai produksi | Tinggi untuk biomassa; terbatas untuk struktur jaringan kompleks [10] | Esensial untuk produk potongan utuh yang membutuhkan otot, lemak, dan ECM [4] |
Sirkuit kontrol rasio menambah beban validasi yang lebih berat.Tetapi umpan balik bawaan mereka sulit untuk dicocokkan hanya dengan kontrol proses, yang menambah tekanan pada pemilihan klon dan pengujian proses.
4. Dari konstruksi ke proses: validasi, peningkatan skala, dan kesesuaian regulasi
4.1 Strategi integrasi dan pemilihan klon untuk kinerja yang stabil
Setelah desain sirkuit, bagian yang sulit dimulai: memasukkan sirkuit tersebut ke dalam sel dengan cara yang tetap stabil melalui pasase skala produksi.
Pengiriman lentiviral sering kali efisien dan memberikan integran stabil dengan cepat. Namun, integrasi bersifat acak. Itu berarti lebih banyak perhatian regulasi, ditambah risiko bahwa ekspresi menurun seiring waktu karena pembungkaman. Sistem transposon seperti PiggyBac dan Sleeping Beauty berada di tengah. Mereka dapat mempertahankan kinerja di banyak pasase, tetapi Anda masih perlu menyaring jumlah salinan dan situs penyisipan. Sleeping Beauty, misalnya, telah digunakan untuk mengabadikan sel satelit sapi secara stabil dengan mengekspresikan TERT dan CDK4 secara berlebihan, dengan garis yang mempertahankan potensi myogenik lebih dari 40 kali pasase [10] . CRISPR knock-in memberikan kontrol paling ketat atas lokasi pendaratan konstruksi dan pengaturan genom yang paling tepat, meskipun pemilihan klon lebih lambat dan throughput lebih rendah.
| Metode Integrasi | Kontrol Penyisipan | Stabilitas | Skalabilitas | Pertimbangan Regulasi |
|---|---|---|---|---|
| Pengiriman Lentiviral | Rendah (integrasi acak) | Tinggi, tetapi rentan terhadap pembungkaman | Tinggi | Pemeriksaan regulasi lebih ketat karena penyisipan acak dan sisa virus |
| Transposon (PiggyBac/SB) | Sedang | Tinggi selama banyak lintasan | Tinggi | Memerlukan penyaringan untuk jumlah salinan dan situs penyisipan |
| CRISPR Knock-in | Tinggi (spesifik situs) | Sangat tinggi | Sedang | Menguntungkan; mengurangi risiko mengganggu gen endogen |
| Vektor Episomal | Tidak ada (ekstrakromosomal) | Rendah; dapat hilang selama pembelahan | Rendah | Hambatan integrasi lebih rendah, tetapi tidak cocok untuk ekspansi jangka panjang |
Penyaringan klon perlu melakukan lebih dari sekadar mengonfirmasi bahwa konstruksi ada.Itu harus melacak pergeseran ekspresi, profil penyisipan, kinetika pertumbuhan, efisiensi diferensiasi, dan retensi fenotip pada jumlah pasase tinggi. snRNA-seq berguna di sini karena dapat mengesampingkan klon yang diperkaya untuk sel cadangan Pax7⁺/Ki-67⁻ - sel yang meninggalkan siklus sel tanpa berdiferensiasi - sebelum skala-up [3]. EPSCs babi dengan sirkuit Tet-On-PAX7 mempertahankan diferensiasi otot tinggi dalam kultur mikrokorier 3D dan suspensi selama lebih dari 40 pasase [8].
4.2 Bagaimana perilaku sirkuit berubah dalam kultur 3D, mikrokorier, dan bioreaktor
Setelah Anda memiliki klon, tes berikutnya adalah apakah ia berperilaku sama di luar 2D. Dalam banyak kasus, tidak. Kinerja dalam 2D jarang sekali dapat diterapkan dengan bersih ke kultur suspensi, mikrokorier, atau scaffold karena gradien difusi, batas oksigen, dan gesekan semuanya menggeser keluaran sirkuit.
Salah satu pemeriksaan pertama adalah difusi inducer. Dalam bioreaktor tangki berpengaduk, inducer molekul kecil perlu mencapai sel secara merata. Dalam praktiknya, gradien dapat terbentuk, terutama dalam kultur mikrokari yang padat dan di dalam agregat atau inti scaffold. Kultur suspensi biasanya lebih cocok untuk daging budidaya skala besar karena mendukung kepadatan sel yang lebih tinggi dan memberikan kontrol proses yang lebih ketat.
Memantau keadaan sel juga menjadi lebih sulit seiring dengan skala sistem. Sinyal reporter fluoresen yang mudah dibaca dengan mikroskopi dalam 2D dapat menjadi sulit untuk dipecahkan dalam konstruksi 3D yang buram. Timer fluoresen - probe yang menggeser emisi dari biru ke merah saat protein matang - dapat memberikan data aktivasi sirkuit waktu nyata in situ [1]. Rute yang masuk akal adalah validasi bertahap: pertama dalam 2D, kemudian dalam konstruksi 3D, lalu di bawah kondisi bioreaktor akhir [3] [8].
4.3 Karakterisasi, dokumentasi keamanan pangan, dan pertimbangan UK/EU
Setelah pengujian proses, karakterisasi harus menunjukkan bahwa fungsi sirkuit, fenotipe, dan keamanan masih terjaga. Tumpukan data inti harus mencakup flow cytometry, qPCR dengan urutan cm, RNA-seq time courses, dan pembacaan fungsional seperti fraksi area rantai berat myosin dan ekspresi myoglobin [1]. Media diferensiasi bebas serum yang dioptimalkan telah terbukti membawa ekspresi myoglobin hingga sekitar 30% dari tingkat yang ditemukan dalam otot sapi asli [3]. Itu memberikan tim tolok ukur yang jelas daripada target yang samar.
Anda juga perlu mendokumentasikan profil protein, asam amino, dan lemak, bersama dengan sifat sensorik [10][3].
Dari sudut pandang regulasi, Inggris dan Uni Eropa menarik garis yang jelas antara garis sel yang diabadikan secara spontan (non-GMO) dan garis yang direkayasa secara genetik. Yang terakhir memerlukan dokumen keamanan yang lebih luas [10][3]. Paket stabilitas harus menunjukkan retensi fenotip dan stabilitas genomik di seluruh jembatan produksi penuh - dari bank sel master hingga sel produksi akhir - dan catatan keterlacakan perlu mencakup setiap lintasan di antaranya [10]. Jika sirkuit bergantung pada inducer kimia, aditif yang aman untuk makanan atau berlisensi seperti asam vanilat lebih disukai daripada doksisiklin [1].
Pengawasan genomik rutin adalah suatu keharusan, dan saklar bunuh diri atau eliminasi yang dapat diinduksi harus didokumentasikan sebagai langkah pengendalian risiko inti [7]. Fungsinya juga harus dimasukkan dalam dokumen keselamatan, terutama karena aturan UK dan EU untuk daging budidaya terus berkembang.
5. Peta jalan praktis dan kesimpulan
5.1 Peta jalan implementasi bertahap untuk tim daging budidaya
Rute terbersih dari konsep ke produksi adalah alur kerja bertahap.
Fase 1 adalah desain. Mulailah dengan mendefinisikan garis keturunan target, kemudian gunakan snRNA-seq untuk mengkonfirmasi hambatan utama sebelum Anda memilih arsitektur sirkuit. Langkah itu penting karena sirkuit hanya dapat menyelesaikan kendala yang benar-benar Anda identifikasi.
Fase 2 adalah membangun dan validasi 2D. Bangun konstruksi dan periksa bahwa sirkuit berperilaku sesuai dengan yang diinginkan dalam 2D, menggunakan pembacaan pelapor sederhana.Pada tahap ini, tujuannya sederhana: memastikan logika berfungsi sebelum beralih ke model yang lebih sulit dan mahal.
Fase 3 adalah pengujian tekanan yang relevan dengan skala. Beralih ke sistem 3D dan kondisi yang relevan dengan bioreaktor, kemudian bandingkan output dengan baseline 2D. Di sinilah banyak desain mulai menunjukkan titik lemahnya, terutama ketika transfer massa, geseran, dan efek matriks mulai berperan.
Fase 4 adalah integrasi regulasi dan keselamatan, dan ini harus berjalan paralel dengan Fase 3. Pekerjaan keselamatan dan regulasi tidak boleh menunggu hingga akhir. Jalankan bersamaan dengan peningkatan skala, termasuk dokumentasi untuk modul keselamatan yang dapat diinduksi.
5.2 Sumber alat dan bahan pendukung melalui Cellbase
Setelah alur kerja ditetapkan, pengadaan sering kali menjadi langkah pembatas laju.
- garis sel
- media bebas serum dan didefinisikan secara kimiawi
- rangka
- komponen bioreaktor
- sensor
- peralatan analitik
Akses yang andal ke bahan yang kompatibel pada setiap tahap memiliki dampak langsung pada seberapa cepat perilaku sirkuit dapat dikarakterisasi dalam kondisi yang relevan dengan skala.
5.3 Poin penting
Sirkuit gen sintetis memberikan tim daging budidaya kontrol terprogram atas waktu, ambang batas, dan keseimbangan garis keturunan yang tidak dapat dicapai oleh protokol hanya media. Pilihan arsitektur membentuk reversibilitas, kebocoran, dan keamanan.Sistem yang dapat diinduksi biasanya lebih disukai karena mereka memberikan kontrol kondisional dan beban metabolik yang lebih rendah [6].
"Perangkat alat biologi sintetis dapat digunakan untuk membangun garis sel dengan ekspresi gen yang dapat disesuaikan, yang, ketika digabungkan dengan PAT dan pemodelan komputasi, dapat memungkinkan sistem kontrol loop tertutup untuk memberikan hasil dan kualitas produk yang optimal." - npj Systems Biology and Applications [6]
Keberhasilan penerapan bukan hanya masalah biologi. Ini tergantung pada keterkaitan erat antara rekayasa sirkuit, desain bioproses, dokumentasi regulasi, dan pengadaan.
FAQs
Bagaimana sirkuit gen sintetis meningkatkan konsistensi diferensiasi?
Sirkuit gen sintetis dapat membuat diferensiasi lebih konsisten karena mereka memberikan kontrol terprogram atas perilaku sel dan komitmen garis keturunan.Dalam praktiknya, itu berarti menggunakan operasi logika modular untuk menyetel ekspresi gen dan faktor transkripsi dengan waktu yang ketat.
Waktu itu penting. Ini membantu sel bergerak melalui perubahan keadaan yang ditentukan dalam urutan yang benar, alih-alih melayang ke nasib campuran atau yang tidak diinginkan. Ini juga mengurangi diferensiasi di luar target dan mengurangi kebisingan di seluruh kultur.
Hasilnya jelas: populasi sel yang lebih seragam, stabil, dan matang untuk produksi daging budidaya.
Desain sirkuit mana yang cocok untuk kontrol miogenik atau adipogenik?
Dalam penelitian daging budidaya, fibroblas ayam yang sama dapat didorong ke salah satu garis keturunan. Miogenesis mengikuti satu set protokol induksi, sementara adipogenesis dapat diaktifkan dengan mengekspos sel ke input seperti serum ayam atau asam lemak.
Dari sana, nasib sel ini dapat dikendalikan langkah demi langkah di dalam kerangka hidrogel 3D untuk membangun struktur daging dengan rasio lemak dan kolagen yang ditentukan.
Mengapa sirkuit gen sering berperilaku berbeda dalam kultur 3D?
Dalam kultur 3D, sirkuit gen sering berperilaku berbeda karena sel-sel menghadapi input fisik dan struktural yang tidak ada dalam lapisan monolayer 2D. Input tersebut termasuk ketegangan mekanis, tekanan geser, kekakuan matriks, dan kepadatan sel lokal.
Isyarat-isyarat ini dapat menggeser jalur sinyal seperti Notch. Mereka juga dapat mengubah cara sirkuit sintetis mendeteksi gaya dan mengoordinasikan respons hilir, termasuk adhesi sel-sel dan morfogenesis jaringan.









