

エピジェネティックサイレンシングは、培養肉の生産アプローチを変革しています。R&Dの専門家にとって、DNAを永久に変化させることなく遺伝子発現を制御する方法を提供し、細胞増殖、分化、品質管理などの重要な課題に対処します。. 以下が知っておくべきことです:
- それが何であるか: DNAメチル化、ヒストン修飾、またはRNA干渉を介した遺伝子活動の抑制 - 遺伝子配列をそのままにしておく可逆的で精密な方法です。
- なぜ重要なのか: 細胞の寿命を延ばし、筋細胞の分化を促進し、スケーラビリティを向上させる一方で、永久的な遺伝子編集による発癌のリスクを回避します。
- 主要なツール: CRISPR-dCas9システム(KRABやDNMT3Aのような)やTALEベースのエディターは、高いサイレンシング効率を達成し、一部の効果は300日以上持続します。 html
- 課題: これらのツールを大規模に提供すること、特にバイオリアクターでの提供や、種特異的な経路に合わせたアプローチの調整が課題として残っています。
バイオプロセスエンジニアや細胞培養科学者にとって、細胞の挙動を正確に制御して生産性と製品品質を向上させることが焦点です。エピジェネティックサイレンシングは、培養肉生産のボトルネック. を克服する鍵となる可能性があります。
家畜細胞におけるエピジェネティックサイレンシングの核心メカニズム
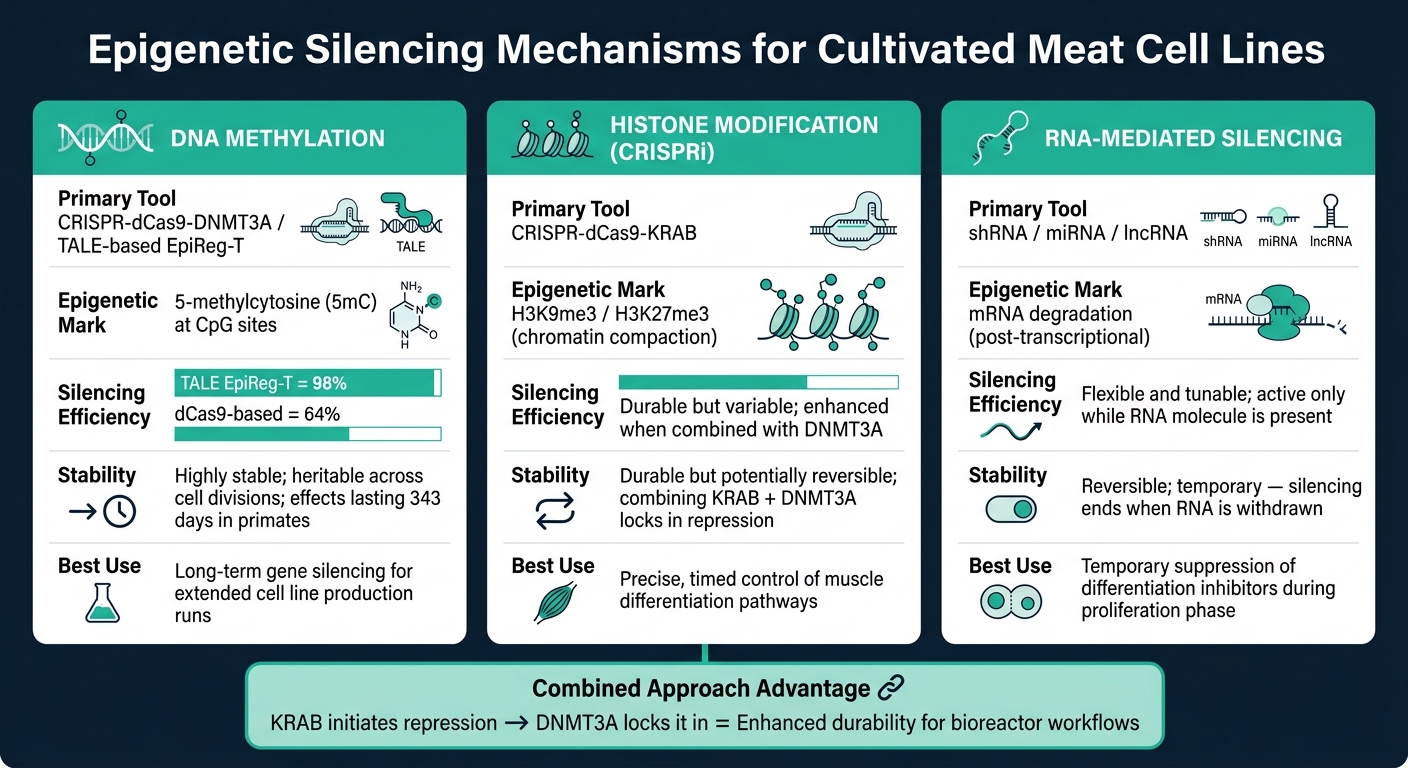
培養肉のためのエピジェネティックサイレンシングツール: メカニズム、効率性&安定性
培養肉細胞株の性能向上は、エピジェネティックメカニズムの正確な制御に大きく依存しています。以下は、家畜細胞で使用される主要な方法の概要です。
DNAメチル化に基づくサイレンシング
DNAメチル化は、DNAメチルトランスフェラーゼ(DNMTs)によって駆動されるCpGサイトへのメチル基の付加を伴います。これが遺伝子プロモーター領域で発生すると、転写機構が遺伝子にアクセスするのを防ぎ、実質的に遺伝子をオフにします[6]. このサイレンシングは遺伝的であり、DNMT1が細胞分裂を通じてメチル化パターンを維持します[7].
高度なツールの一つ、CRISPR-dCas9-DNMT3A, は、触媒活性を持たないdCas9タンパク質とDNMT3A酵素を組み合わせて、特定のゲノム位置にメチル化を誘導します。この方法は、DNAを切断することなく高いサイレンシング効率を達成します。より洗練されたアプローチであるTALEベースのエピジェネティックレギュレーター(EpiReg-T), は、マウスで98%のサイレンシング効率を示し、以前のdCas9ベースのシステムの64%と比較されます [5]. 非ヒト霊長類を対象とした研究では、このシステムの単回投与で遺伝子サイレンシングが最大343日間維持されました [5].
DNAメチル化の確立に続いて、ヒストン修飾が遺伝子調節の二次的で動的な層を提供します。
ヒストン修飾とCRISPRi
ヒストン修飾はヒストンタンパク質を標的にすることでクロマチン構造を変化させ、遺伝子をよりアクセスしやすくまたはしにくくします。H3K9me3や H3K27me3のようなマークはクロマチンをコンパクトにし、転写因子がDNAにアクセスするのを防ぎます [6].
CRISPR干渉(CRISPRi)は、dCas9をKRABリプレッサードメインと融合させて利用します。この複合体は特定の遺伝子プロモーターに導かれ、抑制的なヒストンマークを付加する抑制タンパク質をリクルートします。 羊の研究では、 H3K27me3 が筋肉発達中の重要な抑制シグナルとして強調されており、アクティブなエンハンサーは優れた成長性能を促進する遺伝子に関連付けられています [8]. 家畜の筋肉分化を調節するヒストン状態を理解することで、科学者は細胞の挙動を精密に調整することができます。
"エピジェネティック編集は、遺伝子発現を修正するための有望な戦略であり、ゲノム編集技術の永久的な変化や潜在的な遺伝毒性を避けることができます。" - Nature Biotechnology [5]
ヒストン修飾はDNAメチル化よりも動的であることが多く、その効果を維持するためには持続的またはタイミングを合わせた介入が必要です。KRABとDNMT3Aを単一の構造に組み合わせることで耐久性を高めることができます:ヒストンマークが抑制を開始し、メチル化がそれを固定します[5].
これらのDNAベースの方法に加えて、RNA媒介サイレンシングは柔軟で一時的な代替手段を提供します。
RNA媒介サイレンシング
RNA媒介サイレンシングは、mRNAレベルを直接減少させることに焦点を当てています。マイクロRNA(miRNA)およびショートヘアピンRNA(shRNA) は、相補的なmRNA配列に結合し、翻訳前にそれらを分解します [6]. 一方、長鎖ノンコーディングRNA(lncRNA)は、特定のゲノム領域にクロマチン修飾複合体をリクルートすることによって、より早い段階で作用します [6].
培養肉の応用において、RNA媒介サイレンシングは大きな利点を提供します:可逆性と柔軟性. サイレンシングはRNA分子が存在する間のみ活性を保つため、一時的な介入に理想的です。例えば、増殖期に分化阻害因子を抑制し、その後正常な筋肉発達を可能にするために除去することができます。しかし、RNA分子の継続的な供給を維持することは、バイオリアクター培養のために細胞株をスケールアップする際に. 複雑さを増す可能性があります。
以下の表は、これらのメカニズムの主な特徴をまとめたものです:
| メカニズム | 主要ツール | エピジェネティックマーク | 安定性 |
|---|---|---|---|
| DNAメチル化 | CRISPR-dCas9-DNMT3A | 5-メチルシトシン (5mC) | 非常に安定しており、分裂を超えて遺伝可能[5][7] |
| ヒストン抑制 (CRISPRi) | CRISPR-dCas9-KRAB | H3K9me3 / H3K27me3 | 耐久性があるが、潜在的に可逆的[5][8] |
| RNA干渉 | shRNA / miRNA | mRNA分解 | 可逆的で調整可能[6] |
sbb-itb-ffee270
細胞株の性能向上のためのターゲット遺伝子と経路
エピジェネティックなメカニズムに関する以前の議論を踏まえ、適切な遺伝子ターゲットを選択することは、細胞株の性能向上にとって重要です。これらの介入の成功は、ターゲットを特定するだけでなく、適切なサイレンシング方法を選択することにもかかっています。研究により、抑制されると培養肉細胞株の増殖、分化、代謝安定性などの重要な側面を強化するコアセットの遺伝子ターゲットが特定されています。
増殖と不死化
増殖能力の向上は、しばしばCDKN2AやTP53のような遺伝子をターゲットにすることを伴います。. CDKN2Aは、細胞周期を制限し老化を促進するタンパク質であるp16^INK4Aとp14^ARFをコードしています。CDKN2AをサイレンシングすることでG1/S停止を防ぎ、強力な細胞拡張を可能にします。例えば、 ブタ細胞は CDKN2Aをサイレンシングすることで、18〜30回の継代にわたって筋原性の特性を維持しましたが、野生型細胞は10回の継代でこれらの特性を失いました。さらに、 CDKN2Aの枯渇は、パッセージ20で PAX7の発現を約194倍に増加させ、筋肉幹細胞のアイデンティティにとって重要な要素です [9] .
「CDKN2A遺伝子座を標的にすることは、老化を防ぐか細胞の不死化を誘導するために不可欠です。」 - 食品材料研究[9]
TP53はもう一つの重要なターゲットです。ウシ間葉系幹細胞における600遺伝子のCRISPRスクリーニングで、TP53が増殖を促進するための最も効果的なターゲットとして特定されました。 TP53のノックアウトは、30日間で細胞の豊富さを1,000倍に増加させ、長期的な拡張においても一貫したパフォーマンスを示しました [1] . さらに、PTEN, PI3K/AKT/mTOR経路の負の調節因子をサイレンシングすることで、細胞の倍加速度とmTOR活性が向上します。しかし、このアプローチは、分化効率を低下させる可能性があるため、注意深い監視が必要です[1].
増殖におけるこれらの進歩は、次の重要なステップである分化の最適化の舞台を整えます。
分化の制御
細胞の拡大と組織形成のバランスを取ることは、培養肉生産における複雑な課題です。よく研究されているターゲットの一つはミオスタチン (MSTN), で、筋形成の負の調節因子です。MSTNをサイレンシングすることで、特定の牛の品種に見られる「ダブルマッスリング」特性に似た筋繊維形成が促進されます[4] . MYOD1の活性化と、デジタルライトプロセッシング(DLP)3Dバイオプリンティングのような高度な技術を溝パターンのヒドロゲル上で組み合わせることで、筋細胞の配列と分化が表面機能化によって大幅に改善されます [4] .
もう一つの重要な側面は、SOX2やOCT4のような多能性調節因子の管理です. CRISPR/dCas9-KRABプラットフォームを使用した SOX2の可逆的サイレンシングにより、72時間以内に最大85%の抑制が達成され、編集構造が撤回された後、基礎発現が約90%まで回復します[3] . この可逆性により、細胞増殖中の制御された抑制と、適切な組織発達をサポートするためのタイムリーな解放が可能になります。
ストレスと代謝経路
長期間の生産サイクルにおいて細胞の品質を維持するには、ストレスと代謝の課題に対処する必要があります。TP53は腫瘍抑制因子およびストレスセンサーとしての二重の役割を果たします。培養条件下では、顕著なゲノム損傷がなくても早期に老化を引き起こすことがあります[1] . TP53をサイレンシングすることにより、細胞は初期継代細胞の遺伝子発現プロファイルを保持し、タンパク質合成やDNA複製などの重要な機能を維持します[1].
以下の表は、主要な遺伝子ターゲットとその機能的役割をまとめたものです:
| ターゲット遺伝子 | 経路 | サイレンシングの効果 | 種のコンテキスト |
|---|---|---|---|
| CDKN2A | 細胞周期の抑制 | 老化を防ぐ; ~194× PAX7 のアップレギュレーションが20回の継代で発生 [9] | ブタ |
| TP53 | ストレス応答 / 腫瘍抑制因子 | 30日間で細胞量が1,000倍増加; 一貫した長期拡張 [1] | ウシ |
| PTEN | PI3K/AKT/mTOR | 倍加速度を増加させる; mTOR活性を強化 [1] | ウシ |
| MSTN | 筋形成調節 | 筋繊維の形成と分化効率を向上させる [4] | ウシ |
| SOX2 | 多能性維持 | 幹細胞性から分化への移行を管理; 72時間で85%抑制 [3] | 複数 |
注目を集めている有望なアプローチは、複数の遺伝子を同時にサイレンシングするマルチプレックスターゲティングです。 例えば、CDKN2Aの抑制とGATA4の活性化を組み合わせることで、個別の介入を上回る相乗効果が示されています[9] [10] . このシステムレベルの戦略は、培養肉における最先端の研究開発をサポートする
エピジェネティックツールとデリバリー方法
特定の遺伝子ターゲットを利用するために、研究者は専門のエピジェネティックツールと効率的なデリバリーシステムに依存しています。
合成エピジェネティックプラットフォーム
適切な遺伝子ターゲットを特定することは方程式の一部に過ぎません - これらの遺伝子をサイレンスするために使用されるツールも同様に重要です。培養肉研究に関連する2つのプログラム可能なシステムが際立っています:CRISPRoffとTALEベースのエピジェネティックレギュレーター(EpiReg-T).
CRISPRoffは、dCas9足場をKRABおよびDNMT3A/3Lドメインと組み合わせて使用し、DNAメチル化やH3K9me3などの遺伝可能な抑制マークを導入することで、DNAの切断を伴わずに持続的な遺伝子サイレンシングを実現します。このアプローチは、細胞株を長期間維持するのに特に有用であり、培養肉生産におけるスケーラビリティと一貫性の課題に対処するための重要な要素です。対照的に、TALEベースのEpiReg-Tは、同様のdCas9ベースのシステムで見られる64%に比べて98%の優れたサイレンシング効率を示しました[5] .
2025年10月にNature Biotechnologyに発表された重要な研究は、TALEベースのエディターの可能性を強調しました。研究者たち、Epigenic Therapeuticsや中国科学院, を含む、は、EpiReg-Tの単回投与が脂質ナノ粒子(LNPs)を介してマカクのPCSK9 遺伝子を90%以上の効率で343日間 . サイレンシングしたことを示しました。これは、マルチオミクス解析を通じて確認されたように、最小限のオフターゲット効果で達成されました[5]. このような結果は、耐久性と効力が重要な場合にTALEベースのシステムを際立たせています。
配送の課題
これらのツールを家畜細胞に効果的に届けること - 特に大規模に - は依然として主要な技術的課題です。エピジェネティックエディターは二本鎖DNA切断のリスクを回避しますが、それでも信頼性のある配送メカニズムが必要です。脂質ナノ粒子(LNPs)は、主要な非ウイルスオプションとして浮上しています。彼らはエピジェネティックエディターをコードするmRNAを一時的に送達し、DNA統合なしで持続的な遺伝子サイレンシングを確立する「ヒット・アンド・ラン」アプローチを可能にします[5]. この一時的な性質は、遺伝子改変に関する規制上の懸念が重要な問題である培養肉にとって特に重要です。
しかし、LNPの効率は細胞の種類によって大きく異なることがあります。 初代ウシまたはブタの筋衛星細胞のための製剤の最適化、特にバイオリアクタースケールの設定においては、依然として活発な研究分野です。小規模な実験でうまく機能する送達方法は、撹拌槽型バイオリアクター. では一貫して性能を発揮できないことがよくあります。これらの送達の課題を解決することは、研究を進め、生産を拡大するために不可欠であり、ますます専門化されたプラットフォームによってサポートされています。
どのように Cellbase はエピジェネティック研究をサポートするか&D
エピジェネティックに修飾された細胞株は、正確に検証された試薬を必要とします。研究者は、エピジェネティック修飾に対応した十分に特徴付けられた細胞株、エピジェネティック安定性を維持する定義された培地処方、クロマチンレベルでの遺伝子サイレンシングを確認できる分析ツールへのアクセスが必要です。一般的な実験室のサプライヤーは、培養肉のアプリケーションとの互換性を確保する専門知識を欠いていることが多いです。
培養肉のバイオプロセシングにおけるエピジェネティックサイレンシングの意味
測定可能な細胞株の改善
エピジェネティックサイレンシングは、特に細胞株の生産寿命を延ばす点で、ますます明らかになっている実用的な利点を提供します。一時的な「ヒット・アンド・ラン」戦略を採用することで、ゲノムを永久に変更することなく老化に関与する遺伝子を一時的に抑制することができます[2]. このアプローチは、ウシおよびブタの筋衛星細胞, で成功を収めており、細胞の倍加回数を大幅に増やし、一般的なバイオプロセシングのボトルネックに対処しています。 重要なことに、この方法は可逆的です - 構造体が撤回されると、遺伝子発現はほぼ基準値に戻ります[3]. この可逆的な制御はバイオリアクターのワークフローに理想的であり、細胞が拡張段階で増殖を続け、適切なタイミングで分化を誘発できるようにします。細胞の拡張が強化されることで、組織の分化がより効率的になり、製品の品質が向上します。
組織形成と製品の品質
細胞増殖の向上は、より良い組織形成の基盤を作ります。制御された分化は、エピジェネティックなサイレンシングが最終製品の品質に直接影響を与えるところです。例えば、ウシ細胞の再プログラミングでは、OCT4, SOX2, およびNANOGのような多能性マーカーをサイレンシングすることで、筋原性系統への移行を促進します。このプロセスは、分化プロトコルの30日目までに細長い多核筋管の形成をもたらします。[11].
"mOSKMと多能性マーカーのサイレンシングは、多能性から筋原性系統への移行にとって重要です。" - Frontiers in Nutrition [11]
筋繊維の発達を超えて、脂肪細胞分化経路に対する正確なエピジェネティック制御は、霜降りを達成する上で重要な役割を果たします。霜降りは風味と口当たりの両方に影響を与える重要な要素であり、これらの改善はゲノムに恒久的な変化を加えることなく達成できます。
規制および消費者の考慮事項
細胞増殖と組織形成の進歩は、規制および消費者の視点にも焦点を当てています。 規制当局は、ゲノムに対する非永続的な影響のため、エピジェネティックサイレンシングを一般的に支持しています。dCas9-KRABやTALEベースのEpiReg-Tのようなツールは、二本鎖DNA切断に関連するリスクを回避し、生産中に遺伝的安定性を示さなければならない食品グレードの細胞株に適しています[5].
しかし、トランスジーンフリーの状態を維持することは依然として課題です。2025年5月に発表された、サンパウロ大学とコペンハーゲン大学, の研究者であるカイアナ・レッキアとクリスティーネ・フロイデを含む研究では、この問題が探求されました。彼らは非統合型エピソーマルベクターを使用してウシ胎児線維芽細胞を再プログラムし、コロニーが33回以上の継代で安定している一方で、エピソーマルプラスミドは12回目と17回目の継代で依然として検出可能であることを発見しました[11].
消費者側では、使用された方法についての透明性が重要です。エピジェネティックサイレンシングがDNAを永久に変化させないことを明確に伝えることは、培養肉製品が商業化に近づくにつれて、公共の信頼を築くための鍵となります。
将来の方向性と研究のギャップ
種特異的な課題
この分野での最大の障害の一つは、家畜種における筋原性経路の詳細な理解の欠如です。IGF-1、MAPK/Erk、Wnt/β-カテニンのような経路は人間やマウスでよく文書化されていますが、牛や豚におけるそれらの役割は部分的にしか理解されていません[11]. 完全な地図がないと、エピジェネティックサイレンシングのための特定の遺伝子ターゲットを特定することが大きな課題となります。
筋繊維の構成は、さらに複雑さを増します。例えば、豚のLongissimus筋は約55%のタイプIIb速筋繊維を含んでいますが、これらの繊維は羊や馬のような種には存在しません。これを地域特異的なHOX遺伝子発現と組み合わせると、サイレンシング戦略は各種に合わせて調整する必要があることが明らかになります[13]. 位置的なHOX 遺伝子発現を保持する衛星細胞(後肢筋肉におけるe.g. , HOXA11およびHOXA13)は、さらに事態を複雑にします。これらのパターンは、細胞が急速な増殖または強固な分化に向かう傾向に影響を与える可能性があります [14].
"SCはこれらの位置的シグネチャを保持できるため、その増殖および分化能力は起源の筋肉によって異なる可能性があります。" - npj Science of Food [14]
実際的には、研究者はエピジェネティックサイレンシングを適用する前にHOX遺伝子発現について細胞株をスクリーニングする必要があることを意味します。これらの遺伝子シグネチャは、生物学的バーコードとして機能し、細胞の地域的なアイデンティティを確認し、最終製品の望ましい特性と一致させるのに役立ちます。
このような種特異的な課題は、細胞バンキング戦略の開発において、iPSCなどの代替細胞源を考慮することの重要性を浮き彫りにしています。
iPSCの開発と細胞バンキングへのリンク
誘導多能性幹細胞(iPSC)は、老化しやすく、繰り返しの生検を必要とする衛星細胞に代わる有望な選択肢を提供します。2025年5月、サンパウロ大学とコペンハーゲン大学の研究者たち - カイアナ・レッキアとクリスティーネ・フロイデを含む - は、非統合型エピソーマルベクターを使用してウシiPSC系統を成功裏に開発しました。これらの細胞は33回以上の継代にわたって安定性を維持し、30日目には多核筋管に分化しました [11]. しかし、厳密なゲノムPCRを通じてトランスジェンフリーの状態を確認することは重要なステップです。
関連する問題はエピジェネティックメモリー . iPSCsはしばしば元の体細胞組織の痕跡を保持し、意図した系統からの分化を歪める可能性があります[12]. 細胞バンキングのためには、筋肉や脂肪形成に向けたエピジェネティックプロファイルを既に持つドナー組織を選択することが重要です。さらに、残存する多能性マーカーの効果的なサイレンシングを確保することは、信頼性の高い長期的な細胞バンクを作成するために不可欠です。
堅牢なiPSCプロトコルの開発は、研究努力全体で標準化されたアッセイと一貫したデータ共有の実践の必要性をも強調しています。
標準化と欠損データ
培養肉におけるエピジェネティック介入の可能性を完全に活用するためには、標準化の問題に対処する必要があります。現在、工業規模の生産に必要な広範な細胞倍加中のエピジェネティック安定性を監視するための普遍的なフレームワークは存在しません[12]. 標準化された方法がないため、異なる研究所間で結果を比較することが難しく、生産の拡大に関する決定はしばしば不完全なデータに依存しています。
このギャップに対処するための実用的なステップが役立つ可能性があります。例えば、CD31⁻/CD45⁻/CD29⁺/CD56⁺のようなマーカーをターゲットにした一貫したFACS精製プロトコルを採用することで、種や解剖学的なソースを超えて衛星細胞集団をより比較可能にすることができます[14]. 血清ベースから化学的に定義された培地への切り替えも、バッチ間の変動を減らし、より一貫したエピジェネティック環境を作り出すことができます[12].
将来を見据えると、AI駆動のインシリコモデリングを統合することで、エピジェネティックプロトコルの最適化が革命的に進化する可能性があります。しかし、これらのモデルを効果的にするためには、培養肉研究コミュニティ全体でデータを調和させることが不可欠です。標準化されたデータ共有の実践により、研究者はエピジェネティックな操作の結果をより正確に予測できるようになり、この分野の進展を加速させることができます。
よくある質問
培養肉細胞におけるエピジェネティックサイレンシングは、永久的な遺伝子編集とどのように異なりますか?
エピジェネティックサイレンシングは、遺伝子編集とは異なり、DNA配列に永久的な変更を加えることなく遺伝子の活動を調節します。エピジェネティックなアプローチはDNAを破壊したり修正したりしないため、培養肉の生産においてより安全な選択肢と見なされることが多いです。CRISPRベースのツールなどの技術は、柔軟で場合によっては可逆的な遺伝子調節の利点を提供します。 これらの方法を使用する研究者向けに、
増殖を促進しつつ分化を損なわないためには、どの遺伝子を最初にサイレンシングすべきでしょうか?
細胞の増殖を促進しつつ、分化能力を維持するためには、細胞周期を阻害するか、望ましくない細胞運命をもたらす遺伝子をサイレンシングすることが重要です。例えば、CDKN2Aを抑制することで、ブタの衛星細胞の増殖が著しく向上し、分化の可能性を損なうことはありません。同様に、 TP53やPTENのような腫瘍抑制遺伝子を標的にすることで成長を促進できますが、これらの介入には慎重な監視が必要です。
エピジェネティックエディターをバイオリアクタースケールで信頼性高く届けるにはどうすればよいですか?
培養肉生産のためにエピジェネティックエディターを大規模に届けることは、大きな課題です。これは主に、CRISPRツールの大きさと、エレクトロポレーションやウイルスベクターのような従来のデリバリー方法の制約によるものです。しかし、有望な戦略がいくつか出現しています。例えば、リポソームナノ粒子やエンジニアリングされたウイルス様粒子を使用した一時的なデリバリーシステムは可能性を示しています。これらの方法は、大きなCRISPR貨物をカプセル化し、ゲノム統合を引き起こすことなく細胞内に効率的に侵入させることができます。このような先進的な取り組みを支援するために、









