

培養肉生産の研究者にとって、アポトーシスを最小限に抑えることは、バイオリアクター内での細胞の生存率と生産性を向上させるために不可欠です。栄養素の枯渇、浸透圧の不均衡、廃棄物の蓄積などのストレス要因は、しばしば細胞死を引き起こし、収量を減少させます。抗アポトーシス遺伝子は、培養中の細胞の寿命を延ばすことで、これらの課題を軽減できます。以下は、主要な遺伝子とその役割の概要です:
- BCL-2: ミトコンドリアの孔形成を防ぎ、アポトーシスの開始を阻止します。未分化細胞に効果的ですが、プロアポトーシスタンパク質との慎重なバランスが必要です。
- BCL-xL: 分化中の細胞を保護し、エネルギー代謝をサポートします。バイオリアクターの高ストレス段階に理想的です。
- MCL-1: 栄養変化に迅速に対応し、分化中も安定しています。他の遺伝子と組み合わせて効果的に機能します。
- BIRC5 (Survivin) : カスパーゼを阻害してアポトーシスを下流でブロックします。急速に分裂する細胞の増殖をサポートします。
- XIAP: 高密度培養のような極端なストレス条件下で効果的な強力なカスパーゼ阻害剤です。これらの条件を監視するには、培養肉バイオリアクター用のセンサーを選択して、栄養レベルと廃棄物の蓄積をリアルタイムで追跡する必要があります。
クイック比較
| 遺伝子 | 主要な役割 | 分化中の安定性 | 最適な使用ケース |
|---|---|---|---|
| BCL-2 | 初期アポトーシス(BAX/BAK)をブロック | 安定 | 未分化細胞の保存 |
| BCL-xL | カスパーゼ活性化を防ぎ、代謝をサポート | ステージ特異的 | ストレス下での分化細胞 |
| MCL-1 | 栄養変化への迅速な対応 | 安定 | 多段階の生存 |
| BIRC5 | 下流のカスパーゼを阻害 | 分化とともに減少 | 急速に分裂する細胞 |
| XIAP | 広範なカスパーゼ阻害 | 安定 | 高ストレスバイオリアクター条件 |
1.BCL-2
BCL-2は、内因性(ミトコンドリア)アポトーシス経路において重要な役割を果たす、よく研究された抗アポトーシス遺伝子です。この経路は細胞死の主要なメカニズムであり、培養肉細胞においては、栄養不足や低酸素レベルなどのバイオリアクターのストレスによってしばしば引き起こされます。
BCL-2は、BAXやBAKのようなプロアポトーシスタンパク質に結合して中和することで機能します。この作用により、ミトコンドリアの孔の形成が防がれ、シトクロムcの放出が止まり、アポトーシスの下流カスケードが停止します。このメカニズムは、培養肉生産における細胞の生存可能な寿命を延ばすために重要です。Rønning SBらは次のように説明しています:
"Bcl-2とBaxの比率が、細胞がアポトーシスを起こす感受性を決定する。"[5]
ミトコンドリアでの役割を超えて、BCL-2は小胞体(ER)にも存在します。ここでは、カルシウムレベルを低下させ、IP3受容体を介したカルシウム放出を抑制し、高密度バイオリアクター培養で頻繁に発生するカルシウム誘発性アポトーシスを軽減します。[4]. これらのスケーリングの課題を管理することは、業界の主要な焦点です。この二重局在化により、BCL-2は複数のアポトーシス誘発因子から細胞を保護することができます。
BCL-2の分子構造は、8つのアルファヘリックスバンドルと4つの明確に定義されたBHドメインで構成されており、遺伝子改変のための優れた候補となります。CRISPR/Cas9を介した過剰発現や安定したベクター統合などの技術は、培養肉細胞株におけるBCL-2の保護能力を活用することができます。[4]. さらに、BCL-2はウシやブタのような哺乳類種間で高度に保存されているため、ある細胞株からの発見は、培養肉生産で使用される他の細胞株にも適用されることが多い[3] .
しかし、重要な注意点があります。BCL-2とBAXのようなプロアポトーシスタンパク質とのバランスは慎重に管理されなければなりません。プロアポトーシスシグナルが強すぎる場合、BCL-2の発現レベルが高くてもアポトーシスを防ぐことができないことがあります[2]. このバランスを監視することは、最適な細胞生存率を達成するために不可欠です。
sbb-itb-ffee270
2. BCL-xL
BCL-xL, はBCL2L1遺伝子によってコードされ、外部ミトコンドリア膜に局在し、アポトーシスを防ぐことでBCL-2ファミリーにおいて中心的な役割を果たします。これは、BAXやBAKのようなプロアポトーシスタンパク質に対抗することで達成されます。さらに、これは細胞死を阻止するために不可欠な切断型カスパーゼ-3(CASP3)を抑制します。このメカニズムは、高密度バイオリアクター培養, において、代謝ストレスが細胞の生存率を脅かす可能性がある場合に特に価値があります。
興味深いことに、BCL-xLの活動は特定の分化段階と一致しています。ある段階ではその発現が増加し、他の抗アポトーシスタンパク質、例えばBCL-2やMCL-1は変化しません。これは、分化中の細胞生存を維持する上でのその重要性を強調しています。Cell Death & Disease: で指摘されているように
「BCL-xL/BCL2L1は、分化中の細胞の生存を促進する重要な抗アポトーシスタンパク質です。」[2]
アポトーシスにおける役割を超えて、BCL-xLは細胞のエネルギー代謝をサポートします。これは、解糖系と酸化的リン酸化の両方を強化し、高い代謝活動を確保します。BCL-xLの阻害は、代謝遺伝子の発現を減少させ、基礎および最大のミトコンドリア呼吸を低下させることが示されています。この機能は、持続的な代謝出力に依存する培養肉細胞にとって特に重要です。
BCL-xLは、培養肉研究で一般的に使用される遺伝子編集戦略と非常に互換性があります。レンチウイルス形質導入のような技術は、BCL2L1遺伝子の安定した統合を可能にし、ドキシサイクリン誘導性CRISPR/Cas9システムは、その発現に対する正確な時間的制御を提供します[2] [6]. このレベルの精度は、高度なバイオプロセス制御ソフトウェア. を通じて管理されることが多いです。これらの属性により、BCL-xLは培養肉生産における細胞株の生存率を向上させるための強力な候補となります。
高い代謝要求を持つ分化段階では、BCL-xLはBCL-2よりも効果的である可能性があります。研究者は、永久的な遺伝子改変を進める前に、細胞株のBCL-xLへの依存性をテストするために阻害剤WEHI-539を使用できます[2]. さらに、BCL-xLとMCL-1を共発現させることで、これらのタンパク質が一部の耐性細胞タイプで相乗的に作用することが観察されているため、細胞の生存率をさらに向上させることができます[6].
3. MCL-1
MCL-1(骨髄性白血病-1)は、内因性アポトーシス経路の調節において重要な役割を果たします。外ミトコンドリア膜に存在し、アポトーシスを防ぐために、プロアポトーシスタンパク質BAXおよびBAKと結合してそれらを隔離し、それらのオリゴマー化およびその後の膜透過を阻止します。この作用により、シトクロムcの放出が阻止され、アポトーシスカスケードが実行段階に達する前に停止します[8] . さらに、MCL-1はBim、PUMA、NOXAなどのBH3のみのタンパク質と高い親和性で結合します[8]. BCL-2やBCL-xLと同様に、MCL-1は特にバイオリアクターストレス時にアポトーシスシグナルに対抗するために重要です。
MCL-1のユニークな特性の一つは、その短い半減期であり、栄養の利用可能性や代謝シグナル、特にAMPK/mTOR経路を通じてその発現が非常に敏感に反応することです。研究によれば、カロリー摂取量を25%減少させると、MCL-1の翻訳が約39% ± 10%減少することが示されています[7]. この感受性は、培養肉の生産において特に関連性があり、大規模な懸濁培養中の成長媒体の組成の変動や栄養枯渇(慎重な生産規模の計画が必要)によってMCL-1レベルが大幅に低下する可能性があります。このような減少は細胞の生存率を損ない、抗アポトーシス戦略によって達成されたIVCC(積分生存細胞濃度)の改善を損ないます。これを緩和するためには、強力なmTORC1活性をサポートする無血清培地の配合が不可欠です[7] .
MCL-1のもう一つの注目すべき特徴は、分化中の安定性です。膵臓前駆細胞モデルでは、MCL-1の発現は17日間の分化プロトコルを通じて安定しており、BCL-xLとは異なり、段階に依存した変動を示しました[2]. この安定性は、細胞が正確なタイミングの介入を必要とせずに複数の成熟段階を生き延びる必要がある培養肉の応用において、MCL-1を特に有利にします。&
遺伝子編集ツールを使用して、他の抗アポトーシス遺伝子と同様にMCL-1を修正することができ、細胞株エンジニアリングのための多用途なターゲットとなります。
他の抗アポトーシス遺伝子と組み合わせて使用することで、MCL-1は追加の利点を提供します。例えば、MCL-1とBCL-xLを組み合わせると相乗効果が示されており、両方のタンパク質を同時に阻害することで、生存薬のEC50が約10 μMから20 nM未満に低下しました[6]. このアプローチは、培養肉生産の高ストレス段階での細胞生存率を大幅に向上させることができます。
4. BIRC5 (Survivin)
BIRC5 , は、しばしばSurvivinと呼ばれ、アポトーシス阻害(IAP)タンパク質ファミリーの一員です[2]. BCL-2ファミリーのタンパク質とは異なり、ミトコンドリア膜でアポトーシスの開始を防ぐのではなく、BIRC5はさらに下流で機能します。アポトーシスを実行するカスパーゼをブロックし、プログラムされた細胞死に対する最終防衛線として効果的に機能します[10].
懸濁培養では、栄養素の枯渇、代謝廃棄物の蓄積、機械的せん断応力などのストレッサーがアポトーシスを引き起こす可能性があります。この後期段階でカスパーゼ活性を阻害することにより、BIRC5の過剰発現は細胞の生存率と生産性を延ばすのに役立ちます。これにより、生存細胞濃度の時間積分が改善されます - 細胞培養のパフォーマンスを最適化するための重要な指標です[9]. KAISTの研究者である Eric Baek, は説明します:
"アポトーシス、つまり細胞死を克服することによって生存細胞濃度の時間積分を改善することは、治療用タンパク質[および細胞]の効率的な生産のために最も広く使用されている戦略の一つです。" [9]
この下流の介入は、豚の衛星細胞や牛の筋芽細胞. を含む培養肉の細胞株におけるバイオリアクターの収量を向上させることが示されています。
最も効果的な戦略は、組み合わせ工学, でBIRC5をBCL-2やBCL-xLのようなミトコンドリア保護剤と組み合わせることです。ジョンズ・ホプキンス大学のマイケル・ベテンボー教授はこのアプローチを強調しています:
"カスケードに沿った複数のポイントで細胞死をブロックする戦略は、これらのアポトーシス信号の増幅を制限する可能性があります。" [10]
BIRC5のカスパーゼ阻害を上流のミトコンドリア保護と組み合わせることで、研究者はアポトーシスに対する多層防御を確立できます。
BIRC5はまた、遺伝子編集のワークフローにシームレスに統合されます。CRISPR/Cas9は、過剰発現を伴う安定した細胞株を作成するための主要な方法です[9], が、ジンクフィンガーヌクレアーゼは正確な代替手段を提供します。siRNAは、ゲノム統合に取り組む前に経路の検証に使用できます[9].
5. XIAP
XIAP(アポトーシス阻害因子)は、IAP(アポトーシス阻害タンパク質)ファミリー内で最も強力なカスパーゼ阻害因子として認識されています。BCL-2やMCL-1のような遺伝子と共に、XIAPはアポトーシスの実行段階を標的とする上で重要な役割を果たします。Genes & Development:
で強調されているように"XIAPはin vitroで最も強力なカスパーゼ阻害因子と考えられています。" [12]
XIAPはアポトーシスを阻害するために2つの異なるメカニズムを採用しています。まず、そのBIR2ドメインとリンカー領域がエフェクターカスパーゼ-3および-7をブロックします。第二に、そのBIR3ドメインはカスパーゼ-9を阻害し、内因性ミトコンドリアアポトーシス経路を効果的に停止させます。さらに、そのC末端のRINGドメインは、標的カスパーゼのユビキチン化とその後のプロテアソーム分解を促進します。[11]. 内因性および外因性アポトーシス経路の両方に介入することにより、XIAPは栄養不足、代謝副産物、機械的ストレスなどのアポトーシスの引き金に対処するのに非常に効果的です。これらは培養肉生産システムで一般的に遭遇する要因です。. その機能は、種を超えた強い保存性によってさらに強化されています。
例えば、ヒトのXIAPはBos taurus(ウシ)と87.7%のタンパク質同一性を共有し、Mus musculus (マウス)と89.5%を共有しています。[11]. この高い類似性により、哺乳類モデルシステムからの研究を培養肉生産に使用される細胞株に信頼性を持って適用することができます。
XIAPは、shRNA、アンチセンスオリゴヌクレオチド、またはCRISPR/Cas9などのツールを使用して調節することができます[11]. 極度のストレス下では、そのRINGドメインが自己ユビキチン化を誘導する可能性があります[12], 一方、SMAC/DIABLOやHTRA2のような内因性阻害剤は、カスパーゼからXIAPを置換することができます[11][13]. これらの発見は、培養肉開発のための細胞株を最適化することを目的とした遺伝子編集アプローチにおいて、XIAPを魅力的なターゲットにしています。
抗アポトーシス遺伝子の比較
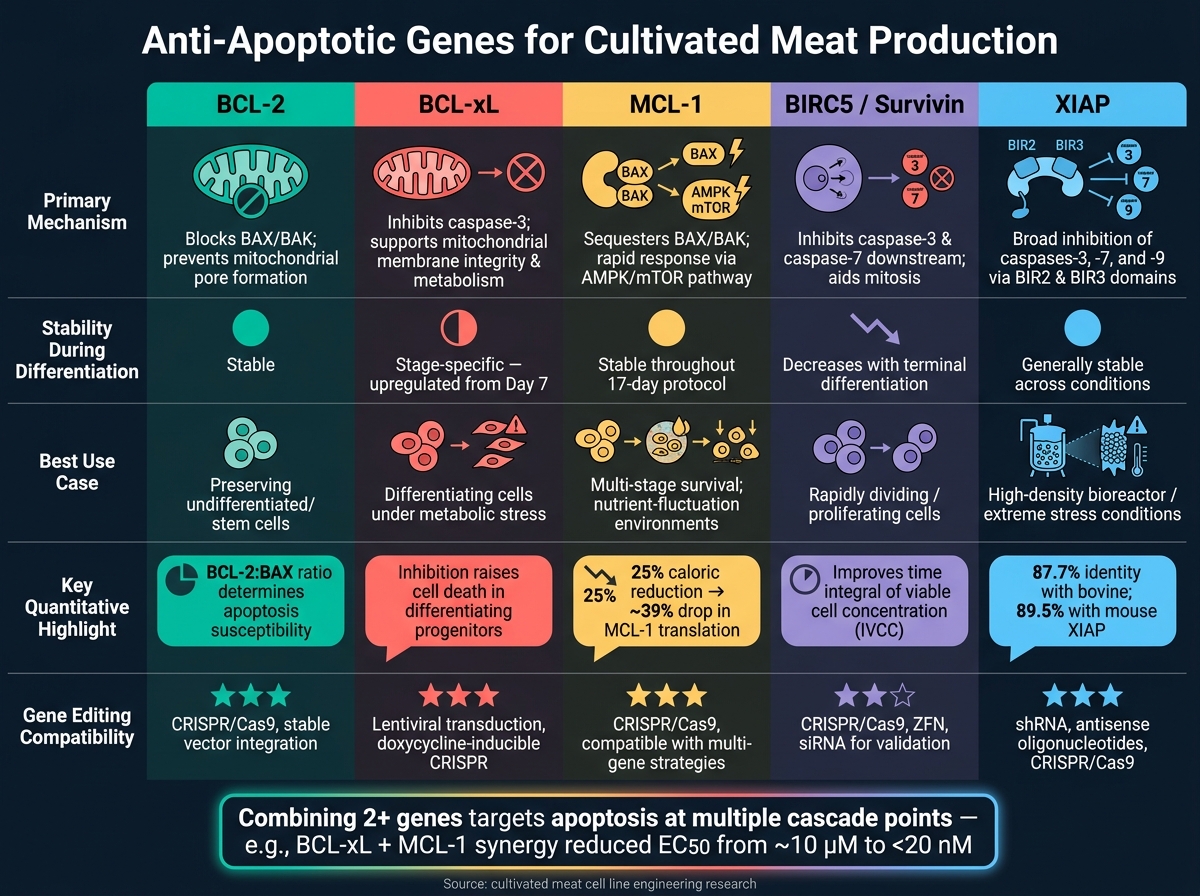
培養肉のための抗アポトーシス遺伝子:並列比較
培養肉の生産に取り組む際、異なる抗アポトーシス遺伝子がどのように機能するかを理解することは、細胞株のエンジニアリングを微調整するのに役立ちます。各遺伝子には独自のメカニズム、分化中の挙動、および潜在的な応用があります。以下の表はこれらの違いを要約しており、どの遺伝子または遺伝子の組み合わせがあなたのニーズに最適かを決定するのを容易にします。
| 遺伝子 | 主要メカニズム | 発現の安定性 | 報告された生存率への影響 | 編集の互換性 |
|---|---|---|---|---|
| BCL-2 | プロアポトーシスBAX/BAKをブロックし、未分化細胞の生存を確保する[2] | 分化中に比較的安定している[2] | 初期の幹細胞プールを維持するために不可欠[2] | 編集ツールとの高い互換性 |
| BCL-xL | 切断されたカスパーゼ-3を阻害し、ミトコンドリア膜の完全性と代謝を維持する[2] | 分化の7日目からアップレギュレーションされる[2] | 分化前駆細胞のサポートに重要であり、その阻害は細胞死を増加させます[2] | 編集ツールとの高い互換性 |
| MCL-1 | BCL-2ファミリーの一部として、プロアポトーシスシグナルを調節します[2] | 系統特異化の間、発現は安定しています[2] | 広範な生存利益を提供しますが、BCL-xLのような段階特異的な効果はありません[2] | 編集ツールとの高い互換性 |
| BIRC5 (Survivin) | カスパーゼ-3とカスパーゼ-7をブロックし、有糸分裂中の染色体分離を支援します | 増殖中の細胞で高く、終末分化とともに減少します | 急速に分裂する細胞の生存と増殖をサポートします | shRNAノックダウンとCRISPR編集の両方に対応 |
| XIAP | 複数のカスパーゼを阻害し、広範なアポトーシス保護を提供 | さまざまな条件で一般的に安定 | 高密度バイオリアクター条件などのストレス下で特に効果的 | 編集ツールとの高い互換性 |
BCL-xL は、プロアポトーシスタンパク質のような重要な分化段階で、細胞の生存を促進し、代謝活動を支援する二重の役割で際立っています。BCL-2, 一方、未分化細胞の保存に理想的であり、XIAPは特に高密度培養のようなストレスの多い環境で広範な保護を提供します。
すべてのシナリオで最適に機能する単一の遺伝子はありません。例えば、BIRC5は急速な細胞分裂が必要な状況で特に有用です。実際には、2つ以上の遺伝子を組み合わせることで、さまざまなアポトーシスの引き金に同時に対処し、最も効果的な保護を提供することが多いです。
これらの発見は、培養肉生産のための細胞株エンジニアリング戦略にこれらの遺伝子を組み込むための基盤を提供します。これには、スケーラビリティを確保するための適切な培養肉の入力の選択が含まれます。
培養肉細胞株エンジニアリングにおけるこれらの遺伝子の使用
培養肉生産における細胞の生存率を向上させるためには、重要な遺伝子を戦略的に統合することが重要です。抗アポトーシス遺伝子を特定するだけでは不十分であり、それらを細胞株に効果的に組み込むことが重要です。一般的に採用される2つの主要な戦略は、抗アポトーシス遺伝子を過剰発現させる ことです。例えば、BCL-2, BCL-xL, およびMCL-1 を用いて細胞の生存を強化するか、プロアポトーシス遺伝子をノックアウトすることです。例えば、BAX, BAK , およびBOK を用いて細胞死の要因を排除します。これらのアプローチを組み合わせることで、大規模生産に適した細胞株が得られることが多いです。[1].
現代の遺伝子編集ツールである CRISPR/Cas9は、Bak1, Bax, およびBok を一度にノックアウトするなど、同時編集を可能にします。ZFNsやRNA干渉のような代替手段は、カスパーゼの活性を一時的に低下させるために使用できます。e.g. カスパーゼ-3、-7、-8、および-9)。過剰発現戦略では、合成プロモーターが遺伝子の一貫した高発現レベルを保証し、 BCL-2のような遺伝子がスケールアップ中に重要であり、 フィードバッチまたは連続培養システム. での細胞性能を維持するために重要です。これらの組み合わせた方法は、培養肉の応用のための細胞株開発を強化します。
このような遺伝子改変は、培養肉生産における重要な指標である総有効細胞濃度(IVCC) , の改善に直接影響を与えます。細胞死は分化の最初の5日間で最も顕著であり、BCL-2やBCL-xLのような遺伝子による早期介入が不可欠です。Cell Death & Diseaseに発表された研究は、 BCL-xLの発現が細胞の分化に伴って増加することを示しており、より成熟した前駆細胞がその保護的役割に大きく依存していることを示しています [2] . 成長段階を通じて BCL-2ファミリー遺伝子の発現レベルを監視することにより、介入を最大の効果を得るために正確にタイミングを合わせることができます。
「抗アポトーシス遺伝子を過剰発現させるか、プロアポトーシス遺伝子をダウンレギュレートする安定した細胞株を確立することにより、細胞が環境ストレスに対してより耐性を持つようになり、最終製品の収量を向上させることができます。」 - Gyun Min Lee et al. [1]
バイオリアクターを用いた生産では、細胞は高浸透圧ストレスや栄養欠乏に耐えるように設計されなければなりません。. スケールアップする前に、Western blotやFACSのようなツールを使用して遺伝子編集を検証することが不可欠です。高密度バイオリアクター環境に特化した細胞株や遺伝子材料を求める研究者にとって、
結論
培養肉の細胞株に対する抗アポトーシス遺伝子の選択には、個別のアプローチが必要です。BCL-2, BCL-xL, およびMCL-1のような遺伝子は、それぞれ細胞を保護する上で独自の役割を果たしますが、その成功は細胞の種類、発達段階、生産中に遭遇する特定のストレスなどの要因に依存します。研究で強調されているように:
"抗アポトーシスとプロアポトーシスのメンバー間のバランスが、最終的に細胞が生きるか死ぬかを決定する"[2]
生存を超えて、抗アポトーシス工学は代謝機能も維持します。例えば、BCL-xLのようなタンパク質は、解糖系と酸化的リン酸化の維持に密接に関連しています。しかし、実行が不十分な介入はこれらの重要なプロセスを混乱させる可能性があります[2]. 設計された細胞株が生産中にその意図されたアイデンティティと代謝活動を維持することは、重要でありながら時に見過ごされがちなステップです。これらの洞察は、細胞株エンジニアリングの未来を形作っています。
新しいマルチジーンアプローチが登場しており、保護遺伝子の過剰発現と、BAX, BAK1, およびBOKのようなアポトーシス促進遺伝子のCRISPRノックアウトを組み合わせて、産業用途により強固な細胞株を作成しています[1]. バイオエネルギーアッセイなどの代謝プロファイリングツールは、これらの遺伝子改変が細胞の全体的な性能を向上させることを確認するために不可欠になっています。豚細胞株, 遺伝子材料、またはバイオリアクター機器を調達する研究者にとって、
よくある質問
私の細胞株にどの抗アポトーシス遺伝子を使用すべきですか?
BCL-2は、細胞株を扱う際の出発点としてよく提案されます。このよく研究された抗アポトーシス遺伝子は、細胞の生存率を向上させる能力で知られており、培養肉研究において人気のある選択肢です。細胞の生存性をサポートするその機能は、初期段階の実験において実用的な選択肢となります。
抗アポトーシス遺伝子を過剰発現させるのと、プロアポトーシス遺伝子をノックアウトするのとではどちらが良いですか?
培養肉の生産においては、BCL-xL, のようなBCL-2ファミリーのメンバーなど、抗アポトーシス遺伝子の発現を増加させる方が、プロアポトーシス遺伝子を無効化するよりも良い結果をもたらす傾向があります。この戦略は、細胞の自然な調節システムを維持しながら、細胞の生存と増殖の両方をサポートし、生産のスケールアップにおいて重要な要素となります。
抗アポトーシス遺伝子の活動を強化することで、細胞はアポトーシスに対する抵抗力を高め、特にストレスの多い条件下での耐性が向上します。これにより、培養プロセス中の細胞の生存率を維持するためのより信頼性が高く安全なアプローチとなります。
抗アポトーシス編集がバイオリアクター内のIVCCを改善することをどのように確認できますか?
抗アポトーシス遺伝子編集がin vitro細胞の生存率と増殖(IVCC)を向上させるかどうかを判断するには、体系的なアプローチが必要です:
- 生存率と増殖率を評価する: 遺伝子編集の前後でこれらの率を測定するために、細胞計数やフローサイトメトリーなどの方法を使用します。
- 遺伝子発現を確認する : qPCRやウエスタンブロッティングなどの技術を使用して、ターゲット遺伝子の発現が成功したことを確認できます。
- アポトーシスマーカーのモニタリング: 編集がアポトーシスを効果的に減少させることを確認するために、カスパーゼ活性のようなマーカーをチェックします。
完全な評価を行うためには、バイオリアクター内で編集された細胞の長期安定性と増殖をテストすることが重要です。これにより、改善が複数の培養サイクルにわたって持続することが保証されます。










