

リボソーム工学は、細胞レベルでのタンパク質合成を改善することにより、培養肉の生産を再構築しています。リボソームは細胞のタンパク質工場であり、肉の食感や栄養価を決定するアクチン、ミオシン、その他のタンパク質を生成するために重要です。しかし、標準的な細胞株は、大規模な肉の栽培に必要な高い生産性に最適化されていません。
主な進歩には以下が含まれます:
- 最適化されたリボソームRNAバリアント: 1.7 × 10⁷のバリアントを持つスクリーニングライブラリーは、翻訳活性の向上の可能性を示しています。
- 直交リボソーム: これらの工学的に設計されたリボソームは、通常の細胞機能を妨げることなく、ミオシンなどの特定のタンパク質を生成することに特化しています。
- コドン最適化: リボソームの好みに合わせてmRNA配列を調整することで、最大72倍のタンパク質発現が得られました。
- マイオカインシグナル伝達: IL-15やマイオネクチンのようなタンパク質は、筋肉分化中のリボソーム生合成とタンパク質合成を促進します。
エネルギー需要のバランスを取ること、細胞の安定性を維持すること、そして生産を産業レベルに拡大することには課題が残っています。例えば、リボソームの過活動は誤った折りたたみのタンパク質や代謝の負担を引き起こす可能性があり、バイオリアクター内の栄養拡散の制限は200μmを超える組織成長を制限します。これらの問題に対処するには、リボソーム工学を高度なバイオプロセッシング戦略と統合する必要があります。
この記事では、これらの方法が培養肉の未来をどのように形作っているか、そして商業的な実現可能性を達成するために克服しなければならない障害について探ります。
リボソームとタンパク質生合成: 入門
哺乳類細胞におけるリボソームの構造と機能
リボソームはタンパク質合成の中心にあり、mRNA配列を機能的なタンパク質に翻訳します。哺乳類細胞において、リボソームは80S粒子として分類され、mRNAをデコードする40S小サブユニットと、ペプチド結合形成を触媒する60S大サブユニットの2つのサブユニットで構成されています。翻訳プロセスは3つの主要なステップを含みます:開始, 開始コドンが認識される段階;伸長, アミノ酸が順次成長中のポリペプチド鎖に追加される段階;そして終結, ストップコドンに達したときに起こる段階。
大サブユニットの2つの特定の領域は、工学的応用において特に重要です:ペプチジルトランスフェラーゼセンター(PTC), ペプチド結合形成を促進する領域と、新たに合成されたポリペプチドが退出する出口トンネル, です。[3].
これらの核心的なメカニズムを理解することは、リボソームの性能を最適化し、培養肉の生産を改善する方法を探るために不可欠です。
培養肉におけるタンパク質生合成の重要性
タンパク質合成の効率は、特にin vitro筋形成中の培養肉の開発において重要な要素です。このプロセスは、筋衛星細胞(MSCs)をアクチンやミオシンのような収縮性タンパク質に富む多核筋線維に変換します。リボソームはこの変換において中心的な役割を果たします[4].
「5,000 Lの容量を持つ従来のバイオリアクターから1 kgのタンパク質を生産するには、約8兆個の筋細胞が必要です」[5]
この驚異的な要求は、リボソーム効率のわずかな改善でも生産量を大幅に向上させ、培養肉の商業的実現可能性に直接影響を与えることを強調しています。
細胞が成熟するにつれて、リボソームの活動は変化を遂げます。増殖期において、MSCは急速な分裂を優先します。しかし、分化が始まってから三日から五日後には、収縮タンパク質の成人型アイソフォームの合成と、細胞が筋管に融合することに焦点が移ります[4]. この移行は、特定のシグナル伝達分子、すなわちマイオカインによって調節されます。
例えば、インターロイキン-15 (IL-15)は、筋発達中の主要な同化因子として、ミオシン重鎖 (MyHC) タンパク質の蓄積を促進し、タンパク質の分解を抑制します [4]. 同様に、マイオネクチンは、PI3K/Akt/mTORシグナル伝達経路を通じてタンパク質合成を強化することで筋肉の成長をサポートします [4]. これらのシグナル伝達経路がリボソーム活性にどのように影響するかを理解することは、生産需要を満たすスケーラブルな細胞株を設計するために重要です。これらの洞察は、後続のセクションで議論されるエンジニアリング戦略の基礎を築きます。
リボソーム工学に関する現在の研究
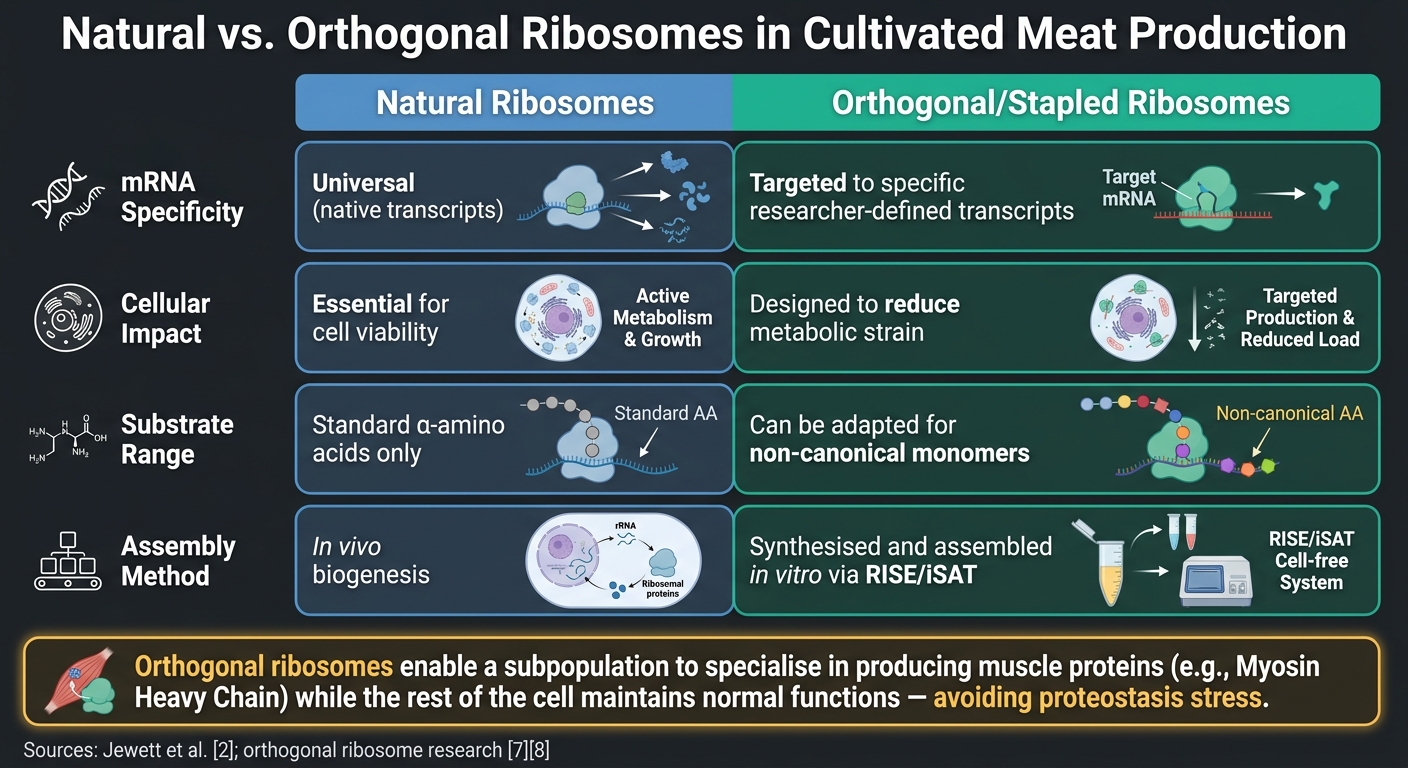
培養肉生産における自然リボソームとオルソゴナルリボソーム
リボソーム生合成と翻訳制御
リボソーム生合成は、細胞が新しいリボソームを構築するプロセスであり、高度に調節されエネルギー集約的な活動です。哺乳類の細胞では、細胞の代謝出力の大部分を占めます。翻訳だけで、細胞の総エネルギー予算の75%を消費することがあり、 [8], 最も資源を要求する細胞プロセスの一つとなっています。
リボソームの割り当てが非効率的な場合 - 例えば、リボソームが初期のコーディング領域で停止する場合 - ボトルネックが生じ、自由なリボソームの利用可能性が減少し、最終的にタンパク質生産が制限されます。計算モデルは、100の遺伝子を工学的に改変することで、リボソームの配分を酵母(Saccharomyces cerevisiae)で35%、大腸菌( Escherichia coli)で57% 向上させることができることを示しています[8]. 。これらの発見は、特にエネルギー効率とタンパク質出力が重要な培養肉産業において、哺乳類細胞のリボソーム動態を最適化するための直接的な影響を持っています。
培養肉の文脈におけるリボソーム工学
リボソーム工学の進歩は、リボソーム生合成の基礎知識に基づき、培養肉の生産に応用されています。筋細胞で直接行われていない研究でさえ、培養肉の細胞株に関連する洞察をもたらしています。
2020年12月、テルアビブ大学のハダス・ズールとタミール・タラーは、成長率とタンパク質出力を向上させるための リボソーム交通工学 (RTE)の可能性を示しました。 CRISPR-Cas9, を使用して、RPO21と CYS4のランプ領域(コドン11–50)に同義変異を導入しました。 S. cerevisiae. その結果、二重変異体は対数成長期の成長と細胞密度が改善されました。しかし、研究者たちは、翻訳最適化と成長率の関係は、二相性シフトおよび静止期においては、翻訳以外の要因が速度制限要因となるため、低下することを警告しました[8]. この洞察は、培養肉生産における分化プロトコルの設計に特に関連しています。
2020年2月、ノースウェスタン大学のマイケル・ジュエットのチームは、 RISE(リボソームのインビトロ合成と進化)メソッドを検証しました。この技術は、約 1.7 × 10⁷のリボソームRNA変異体のライブラリをスクリーニングすることを含みます。 [2]. RISEは生細胞の外で完全に動作するため、致死的なリボソーム変異によって課される制約を回避し、これらはin vivoでは研究できません。
「インビトロアプローチは細胞の生存性の制約を克服し、致死的なリボソーム変異の探求を可能にします。」 - マイケル・ジュエット他 [2]
培養肉のもう一つの有望な革新は、オルソゴナルリボソームの使用です。. これらのエンジニアリングされたリボソーム–mRNAペアは、細胞のネイティブな翻訳機構とは独立して機能します。これにより、研究者は必須の細胞プロセスに干渉することなく、筋肉の質感に重要なミオシン重鎖(MyHC)アイソフォームなどの特定のターゲットにリボソーム活性を集中させることができます[6]. 比較研究は、自然のリボソームに対する直交リボソームの利点を強調しています:
| 特徴 | 自然リボソーム | 直交/ステープルリボソーム |
|---|---|---|
| mRNA特異性 | 普遍的(ネイティブ転写物) | 特定の研究者が定義した転写物にターゲット[6] |
| 細胞への影響 | 生存に必須 | 代謝負担を軽減するように設計[7] |
| 基質範囲 | 標準α-アミノ酸 | 非標準モノマーに適応可能[7] |
| 組み立て | 生体内生合成 | 合成および組み立ては、RISE/iSAT [2] |
ここでの重要なポイントは、直交リボソームがリボソームのサブポピュレーションに筋肉タンパク質(MyHCなど)の生産を専門化させる一方で、細胞の残りの部分は通常の機能を維持することを可能にすることです。 これは、特定のタンパク質を過剰生産するように翻訳システム全体が押し付けられたときに発生する可能性のあるプロテオスタシスストレスのリスクを回避します。
リボソーム性能を向上させるための戦略
リボソーム生合成の増加
リボソームの数を増やすことは、タンパク質生産を強化する直接的な方法であり、2つの主要な方法が注目されています。最初の方法は、リボソームRNA(rRNA)遺伝子のエピジェネティックな状態を変更して、その翻訳能力を高めることです。
"リボソームRNA遺伝子のエピジェネティック工学はタンパク質生産を強化します。" - Santoro R., Lienemann P., Fussenegger M. [1]
2番目のアプローチは、PI3K/Akt/mTORシグナル伝達経路を活用します。IL-15、マイオネクチン、イリシンのようなマイオカインは、この経路を活性化し、以前に議論されたように筋管成熟中にリボソーム生合成を促進します。
しかし、リボソームの生産の増加は、リボソーム合成が生細胞において最もエネルギーを消費するプロセスの一つであるため、細胞の代謝能力と慎重にバランスを取る必要があります[1].
リボソームの数が増加したら、それらが翻訳に完全に従事していることを確認することに焦点が移ります。
翻訳開始と伸長の改善
すべてのリボソームの活動を最大化することは不可欠です。成長が最適化された細胞でも、15〜20%のリボソームが非活性のままです[9]. これは、培養肉細胞株における未開発の能力の重要な予備を表しています。
翻訳伸長の速度は、リボソームの固有の速度と翻訳に積極的に従事しているリボソームの割合の2つの要因に依存します[9]. これらを最適化するためには、培地中のアミノ酸レベルを高く維持することが重要です。さらに、リボソームタンパク質を安定化するために細胞株を工学的に設計することは、rRNAの誤折り畳みや分解から保護し、成長のピーク時におけるrRNAの典型的な10%の損失を減少させます[9].
リボソームの活動が最大化されたら、タンパク質合成をさらに加速するためにmRNA配列を精緻化することが次のステップとなります。
mRNA最適化とコドン使用
リボソームの性能は、処理するmRNAの質に大きく依存しています。コドン最適化は、標的タンパク質のコーディング配列を宿主種特有のtRNAプールに合わせて調整します - 例えば、ウシ、ブタ、または魚。この調整により、伸長中のリボソームの停止を防ぎ、MyoDやMyf5のような重要な筋原性タンパク質のスループットを増加させます。
コドン最適化に加えて、転写調整は細胞内のrRNAとmRNAレベルの適切なバランスを確保します。これらのコンポーネント間の不一致はボトルネックを生み出し、全体的な効率を低下させる可能性があります[1].
実用的な応用として、統合合成、組立、および翻訳(iSAT)システムは貴重なツールを提供します。これらのシステムは、細胞フリー抽出物と蛍光ベースのアッセイを使用して、最適化されたmRNAをin vitroでプロトタイプ化し、それを安定した細胞株に統合する前に使用します。この反復的なアプローチにより、研究者はコドン最適化されたバリアントを迅速に比較し、重要な筋原性タンパク質の収量を向上させ、培養肉生産のスケーラビリティを強化することができます[1].
トレードオフ:成長、分化、および製品品質
リボソームの性能を最適化することは、タンパク質合成を促進することと、細胞の成長と分化への影響を管理することの間の微妙なバランスを伴います。
代謝負担とプロテオスタシスストレス
タンパク質生産を強化するためにリボソームを工学的に改良することは、ATPやアミノ酸を他の重要な細胞機能から逸らすため、エネルギー需要の増加を伴います。リボソーム合成はすでに細胞内で最もエネルギー集約的なプロセスの一つであり、さらなる増幅はこれらのエネルギー課題を悪化させる可能性があります。
この強化された活動は、タンパク質の品質にも影響を与える可能性があります。過剰に活性化されたリボソームは細胞シャペロンを圧倒し、誤った折り畳みのタンパク質を生じさせ、未折り畳みタンパク質応答(UPR)を活性化させる可能性があります。このようなストレスは成長を抑制したり、細胞死を引き起こすことさえあります。牛や羊のような家畜種の成体幹細胞は、自然に増殖能力が限られているため、これらの追加のストレスは、老化が始まる前に有効な細胞分裂の回数を大幅に減少させる可能性があります。[5].
培養肉の生産において、組織の厚さは栄養拡散の制約により200μmを超えることは稀であり、これが大きな組織凝集体の中心部での細胞死を引き起こす可能性があります[5]. エネルギー消費を増加させる戦略は、これらの重要な領域での栄養枯渇を加速させるリスクがあり、安定したタンパク質合成が不可欠です。さらに、代謝負荷の増加は、筋肉分化に必要な微調整されたシグナル伝達経路を妨げる可能性があります。
筋肉分化とタンパク質組成への影響
リボソーム工学によって引き起こされるストレスは、代謝を超えて筋肉の発達を妨げる可能性があります。筋形成のプロセスである筋形成は、転写因子の厳密に調整されたシーケンスに依存しています:Pax7は幹細胞が静止状態を維持することを保証し、Myf5は筋芽細胞の増殖を促進し、 MyoDは分化を引き起こします [5] . タンパク質合成の変化はこのシーケンスを乱し、分化を停滞させたり、非典型的な筋繊維組成を生じさせる可能性があります。これにより、培養肉の望ましい食感と風味を達成するために重要な筋内脂肪沈着が減少する可能性があります[5].
その結果、エンジニアリングプロセス全体で筋形成マーカーの発現を監視することによって厳格な品質管理を維持することが、適切な筋肉の発達と製品の品質を確保するために不可欠です。
sbb-itb-ffee270
研究のギャップと将来の方向性
リボソーム工学の進歩は有望ですが、商業的な培養肉生産への応用には依然として大きな課題があります。これらのギャップを埋めるためには、研究者は高度な分子プロファイリング技術と、長期生産の要求に耐えうるスケーラブルなバイオプロセス戦略に焦点を当てる必要があります。
マルチオミクスと長期安定性研究
大きな課題は、エンジニアリングされた細胞株の長期安定性データの欠如にあります。時間が経つにつれて、これらの細胞は自発的な突然変異を蓄積し、潜在的にその表現型を変える可能性があります。ノヴィサド大学のイヴァナ・パイチンはこの懸念を強調しています:不死化細胞は「長期培養中の潜在的な自発的突然変異のため、常に一次培養を代表するものではありません」[13]. リボソーム工学系統においては、リボソーム成分の変異が翻訳効率を損なう可能性があり、即座に検出されない場合もあるため、リスクはさらに高まります。
マルチオミクスアプローチは、これらの問題に対処する方法を提供します。トランスクリプトミクス、プロテオミクス、メタボロミクスを統合することで、研究者はPax7, MyoD, およびMyogenin, のような重要な筋原性マーカーやMyHCアイソフォームの変化を監視できます。ゲノムスケールの代謝モデルは、これらの洞察をメディア組成の実行可能な変更に変換し、工学的リボソームの独自の要求を満たすことができます[5][11]. 培養肉においては、長期間にわたる一貫したタンパク質生産を確保することが不可欠です。このような長期的な監視がなければ、持続可能な改善と短命な効果を区別することは困難です。
遺伝的および代謝的安定性に加えて、これらの革新を産業レベルに拡大することは独自の課題を提示します。
バイオプロセスの統合とスケールアップ
リボソーム改変細胞を小さなフラスコから産業用バイオリアクターにスケールアップすることは容易ではありません。5,000 Lの撹拌タンクバイオリアクターで1 kgのタンパク質を生産するには、約8兆個の筋細胞が必要です[5]. このような密度では、栄養素の勾配が重要な問題となります。酸素や他の栄養素の200 μmの拡散限界は、3D組織構造の中心部にある細胞が飢餓に直面する可能性があることを意味します。特に高いタンパク質合成のために資源の需要がピークに達しているときに。
バイオリアクターの攪拌によるせん断応力は、複雑さをさらに増します。未改変の細胞はこの乱流に耐えられるかもしれませんが、翻訳機構が改変された細胞はより脆弱である可能性があります。ストレスは細胞経路を妨げるだけでなく、代謝負荷を受けている細胞に物理的な損傷を与える可能性もあります[13]. これらの問題に対処するには、リアルタイムデータをデジタルバイオマニュファクチャリングモデルと統合し、計算流体力学シミュレーションを含めて、大規模容器内の多様な微小環境をよりよく理解し予測する必要があります[10]. 収穫のような下流プロセスも注意が必要です - トリプシンを用いた酵素法は、エンジニアリングされた細胞の表面プロテオームを変化させる可能性があります[14], リボソーム工学の利点を無効にする可能性があります。
| スケールアップファクター | 主要なボトルネック | リボソーム工学への関連性 |
|---|---|---|
| 栄養素の拡散 | 200 μmの浸透限界[5] | 3D組織で高タンパク質合成要求のある細胞を飢餓状態にする可能性 |
| 遺伝的安定性 | 自然発生的な突然変異[13] | 時間とともに工学的翻訳効率を損なう可能性 |
| せん断応力 | 撹拌槽の乱流[13] | 工学的細胞経路を破壊するリスク |
| 収穫方法 | トリプシンによるプロテアーゼ損傷[14] | プロテオームを変化させ、タンパク質品質の改善を隠す可能性 |
これらのスケールアップの課題を解決することは、リボソーム工学を研究室から商業生産に移行するために不可欠です。 各戦略は、工業条件下で信頼できるタンパク質の収量、安定性、安全性を確保するために厳密にテストされなければなりません。
結論: 培養肉におけるリボソーム工学の必要性
5,000 Lのバイオリアクターで1 kgのタンパク質を生産するには、驚くべきことに8兆個の筋細胞が必要です[5]. これは、培養肉生産のスケーリングにおける巨大な課題を浮き彫りにしています。リボソーム工学は、単に細胞数を増やすのではなく、個々の細胞のタンパク質出力を改善することで解決策を提供します。
リボソーム工学を適用する際のタイミングは非常に重要です。翻訳を誤った段階で強化すると、筋形成を妨げ、MyHCのような重要な収縮タンパク質の生産に影響を与える可能性があります[5]. 翻訳と筋形成のバランスを適切に取ることは、工学そのものと同じくらい重要です。
"高品質なCBMとその高収率での生産を達成するためには、商業生産のための良好な実験室実践を達成するために分子の側面を徹底的に検査する必要があります。" - Asim Azhar et al., Frontiers in Food Science and Technology [5]
翻訳開始因子(eIF3iおよびeIF3c)の過剰発現、コドン最適化、mRNA修飾のターゲティングなど、組換えタンパク質の出力を増加させるいくつかの技術がすでに有望であることが示されています[15]. しかし、これらの方法は、代謝負荷、プロテオスタシスストレス、長期的な遺伝的不安定性などの問題を避けるために注意して適用する必要があります。分子最適化は重要ですが、栄養拡散の限界、剪断応力感受性、収穫中のプロテオームの破壊などの課題を完全に解決することはできません。これらの障害は、バイオプロセス設計における同時進行の進歩を必要とします。
培養肉の潜在的な環境利益は非常に大きいです。従来の家畜農業と比較して、温室効果ガスの排出を78%〜96%削減し、土地利用を99%削減し、水使用を82%〜96%削減する可能性があります[12]. これらの利益を大規模に達成するには、現在の細胞培養の生産性と経済的実現可能性のギャップを埋めることが必要です。リボソーム工学はこのギャップを埋めるための強力なツールですが、分子生物学、バイオプロセスの革新、包括的なマルチオミクスモニタリングを含む、より広範で統合されたアプローチの一部でなければなりません。これらの努力を組み合わせることによってのみ、培養肉の完全な可能性が実現されます。
どのようにCellbase が培養肉研究をサポートするか
分子最適化から培養肉の大規模生産に進むには、各段階で正確なツールと材料が必要です。
細胞系の最適化に取り組むチームにとって、
生産のスケールアップに関しては、
よくある質問
培養肉の細胞株に最も有望なリボソーム工学アプローチはどれですか?
培養肉のためのリボソーム工学の研究は、タンパク質の生合成を強化し、細胞の運命決定に影響を与えることを目的としています。有望なアプローチの一つは、翻訳効率を改善するためにリボソームRNAオペロンを修正するリボソームプール工学, です。iSATやRISEのようなツールは、in vitroでのリボソーム進化のためのプラットフォームを提供し、機能性が向上したリボソームの開発を可能にします。さらに、
翻訳率を高めることなく、誤ったタンパク質の折りたたみや細胞ストレスを引き起こさずに、どのようにして翻訳率を向上させることができますか?
翻訳率を向上させる際にタンパク質の誤った折りたたみや細胞ストレスを引き起こさないようにするために、研究者は翻訳プロセス全体を加速するのではなく、微調整に焦点を当てています。主なアプローチには以下が含まれます:
- 遅い翻訳コドンの使用: これにより、翻訳のペースをタンパク質の自然な折りたたみプロセスと一致させ、適切な構造形成を確保します。
- 5' コーディング領域の自由折りたたみエネルギーの削減: この調整により、細胞の健康を維持しながらタンパク質生産効率を向上させることができます。
その他の技術には、低誘導レジーム, 温度ダウンシフト, および SINEUP RNA. のような高度な合成ツールが含まれます。これらの戦略により、細胞に過負荷をかけることなく、より高いタンパク質収率を実現できます。
専門的な材料を扱う方々にとって、
200 µmを超えるリボソーム工学筋組織をサポートするために、バイオリアクターにはどのような変更が必要ですか?
200 µmを超える筋組織を成長させるには、バイオリアクターは栄養素、酸素、pHの拡散に関連する課題を克服する必要があります。これらは三次元構造における細胞の生存に不可欠な要素です。撹拌槽型バイオリアクターは、均一な条件を維持しつつ、細胞を損傷する可能性のある剪断応力を低減するために正確な調整が求められます。多くの場合、灌流ベースのシステムが安定した環境を作り出す上で重要な役割を果たし、特に密集した組織において重要です。専門的なバイオリアクターや材料を扱う方々にとって、









