

細胞を拡張できても、適切なタイミングで適切な運命に切り替えられない場合、プロセスは分化で停滞します。 ここが核心です: 合成遺伝子回路は、 細胞内のコミットメント、タイミング、メモリ、系統のミックスを制御しますが、メディアの変更だけではしばしば 不均一な, 部分的にコミットされた集団が残ります。
培養肉の分化ワークフローを構築するなら、この記事からすぐに4つのポイントを取り入れます:
- 構築物ではなく、ネイティブネットワークから始める。 snRNA-seq, の軌跡分析、GRN推論、miRNAプロファイリングを使用して、細胞がどこで停滞、漂流、または誤った運命に分岐するかを見つける。
-
回路タイプをプロセスの問題に合わせる。
トグルスイッチはロックインに適しており、 フィードフォワードまたはバンドパス設計はタイミング制御に適しており、 論理ゲートはマルチシグナルゲーティングに適しており、 miSFITsは段階的出力に適しています。 - 初日から低漏れ、低ノイズ、安全性を考慮した設計を行います。 直交部品、負の自己調節、iFFLs、cmトランスジーン、誘導可能なキルまたは成長停止モジュールは、ビルドの一部であり、後から考えるものではありません。
- 早期にスケールに関連する条件下で検証します。 2Dで動作する回路は、 3D、マイクロキャリア、または撹拌懸濁液では、誘導体の勾配、酸素制限、せん断のために変化する可能性があります。
この記事は、プロセスチームにとって重要な実用的なポイントも示しています:単一系統制御と比率制御は異なる作業です. A Tet-On MyoDカセットは筋原性のエントリーを促進するかもしれませんが、ホールカット製品には 筋肉、脂肪、ECMの割合, の制御が必要であり、通常はフィードバック、パラクリンシグナル伝達、より厳しいクローンスクリーニングを意味します。
そのメッセージを支えるいくつかの数字:
- 標準的な筋原性分化は、融合指数が約50–60%で停滞する可能性があります
- iPS細胞におけるエンジニアードGRNは、ターゲット系統の分化を52%から81%に増加させました
- 修飾されたMSCにおける合成回路は、心臓分化を76%に促進しました
- 一部のブタのTet-On-PAX7系統は、40継代を超えて高い筋原性ポテンシャルを維持しました
- ヒト多能性幹細胞の約20%が癌関連の変異を持っている可能性があり、そのため誘導可能な安全モジュールが重要です
培養肉分化のための合成遺伝子回路ワークフロー
科学者の物語: マイケル・エロウィッツ、細胞内の遺伝子回路
クイック比較
| トピック | 記事が平易な言葉で言っていること |
|---|---|
| 主なボトルネック | 拡張ではなく差別化 |
| 回路が役立つ理由 | 細胞内にしきい値制御と細胞状態メモリを追加する |
| 最初の最善のステップ | ネイティブな運命制御をsnRNA-seqと関連ツールでマッピングする |
| 主な回路オプション | トグル、フィードフォワード、バンドパス、論理ゲート、miSFITs、CRISPRa/i |
| 筋形成の例 | 成長と終末分化を分けるためのTet-On-MyoD |
| 脂肪生成/ECM制御 | miSFITsと段階的なPPARG/BMP4タイプの出力のためのボウタイデザイン |
| ホールカットの課題 | 筋肉、脂肪、結合組織間の比率制御 |
| スケールアップのリスク | 2Dの挙動は3Dやバイオリアクターでは保持されない可能性がある |
| 統合の選択肢 | レンチウイルス、トランスポゾン、CRISPR ノックイン、エピソーマルベクター |
| 規制上のポイント | エンジニアリングされたラインにはより広範な安全パッケージが必要です。バニリック酸のような食品安全な誘導剤が可能な限りDOXよりも好まれます |
つまり、簡単に言えば:これは単なる回路設計の記事ではありません. 回路アーキテクチャ、系統生物学、クローン選択、バイオリアクターの性能、UK/EUの安全文書を一つの差別化戦略に結びつけるためのガイドとして読んでください。
ネイティブネットワークマッピングから構築選択、スケールアップチェック、規制適合までの完全な道筋を知りたい場合は、読み進めてください。
2. 分化制御回路の設計原則
2.1 回路を設計する前にネイティブな細胞運命ネットワークをマッピングする
回路を設計する前に、細胞が既に何をしているのかを明確に把握する必要があります。
シングルヌクレウスRNAシーケンシング(snRNA-seq)は始めるのに良い場所です。これは、NOTCH2と HEYL, でマークされた予備細胞を含む静止サブポピュレーションを示し、分化を改善する可能性のある経路ターゲットを指摘することができます [3].
そこから、軌道解析と遺伝子調節ネットワーク(GRN)の推論が、調節因子の活性化順序をマッピングし、細胞が停滞しやすい箇所を強調します。筋形成において、主なカスケードはMYOD1とMYOGを通ります。脂肪形成において、主なノードは PPARGとCEBPAであり、線維脂肪前駆細胞(FAP)の運命が主な分岐点リスクとして作用します。以下の表は、主な調節因子とボトルネックをまとめたものです。
| 系譜 | 主要なマスターレギュレーター | 重要なシグナル伝達経路 | 特定されたボトルネック |
|---|---|---|---|
| 筋原性 | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | 予備細胞形成(静止) |
| 脂肪生成 | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | 線維脂肪前駆細胞(FAP)の運命 |
| 多能性 | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | 自発的分化 / 異質性 |
もう一つの有用な層はmiRNA発現プロファイリング. 内因性miRNA、例えばmiR-302a, 多能性に関連し、miR-375, 分化に関連するものは、センス・アンド・レスポンス設計における内部分類子として機能することができます。 それにより、回路は外部の誘導体に依存するだけでなく、セルの実際の状態を読み取ることができます。[5].
これらのボトルネックは回路選択を促進するべきです。主な問題がドリフトである場合、ロックインが必要かもしれません。タイミングが問題である場合、パルスがより適しているかもしれません。運命の制御が複数のキューに依存する場合、マルチ入力ロジックが通常より理にかなっています。
2.2 適切な回路アーキテクチャを選択する
ここでトレードオフが明確に現れます。適切なアーキテクチャは、コミットメントの永続性、タイミングの厳密な制御の必要性、セルが問題なく運べる遺伝的ペイロードの量という3つの実用的なポイントに依存します。
双安定トグルスイッチ は、系統のコミットメントをロックしておく必要があるケースに適しています。主な問題は、転写ノイズによって引き起こされる自発的な状態の反転です。
バンドパスフィルターは、転写因子が特定の発達期間内でのみ必要とされる場合に適しています。問題は、誘導体のレベルを厳密に制御する必要があることで、そうでないとタイミングがずれてしまいます。
論理ゲートは、同時に複数の入力を必要とすることで特異性を追加します。例えば、外因性の誘導体が存在し 、細胞が適切な内因性miRNAプロファイルを示す場合にのみ分化が許可されます。これにより、ターゲット外のコミットメントのリスクを減らすことができます。
以下の表は、主なアーキテクチャとそれに伴うトレードオフを概説しています。
| アーキテクチャ | 可逆性 | 時間的精度 | 統合の複雑さ | 主な使用例 | 主要なリスク |
|---|---|---|---|---|---|
| 双安定スイッチ | 低い(ロック後) | 中程度 | 中程度 | 永久的な系譜の確定 | ノイズによる自発的な反転 |
| バンドパスフィルタ | 高い(濃度依存) | 高い | 高い | 一時的な発達段階 | 厳密な誘導体制御が必要 |
| 論理ゲート(AND/OR/NOT) | 可変 | 中程度 | 中程度〜高い | 細胞型特異的な活性化 | OFF状態での漏れ |
| 蝶ネクタイ / マルチ入力 | 高い | 中程度 | 中程度 | マルチシグナル統合 | 内因性miRNAの安定性に依存 |
| miSFITs | 高い | 中程度 | 低〜中程度 | 段階的な出力調整 | 調整が不十分な場合、狭い動的範囲 |
「機能を維持しながら計算層の数を最小限に抑えることで、この戦略は遺伝子回路工学におけるスケーラビリティの障壁に対処します。" - Nature Communications [9]
追加される各規制層はペイロードを増加させ、細胞資源を消費します。実際には、2つの設計が同じ仕事をする場合、規模が重要なときには通常、より単純な方が良い選択です。
アーキテクチャが設定されたら、次の仕事は低漏れ性、ノイズ抑制、フェイルセーフ制御の下でそれを維持することです。
2.3 信頼性、低漏れ性、安全性を考慮した構築
回路は長期間の培養にわたって安定していなければなりません。生産用途には、短期間の良好な性能だけでは不十分です。
直交部品は最初の防御です。ネイティブの機械と交差反応しないプロモーター、転写因子、規制要素は、オフターゲット効果を制限し、内因性シグナルが回路をオンにしたりシャットダウンしたりする可能性を減少させます。修正された高密度プロモーター、例えばPCREmは、誘導性哺乳類システムにおける基礎的な漏れを減少させるために使用されています[6] .
可能であれば負の自己調節を追加する価値があります。これは転写ノイズを減少させ、誘導物質濃度に対するより線形な応答を生み出すためのよく知られたモチーフの一つです[6]. 非整合フィードフォワードループ(iFFLs) は、確率的な変動をフィルタリングすることにより、短いノイズスパイクではなく持続的な信号に細胞が応答するように、もう一つの制御層を追加できます。
コドン修飾された(cm)合成転写因子のバージョンは、特性評価を容易にします。また、検証中に回路駆動の発現を内因性ゲノム( g)発現から分離することができます[1]. それは小さな詳細のように聞こえるかもしれませんが、回路からの読み出しかホストゲノムからの読み出しかを判断しようとしているときに時間を節約します。
安全モジュールが必要です。 ヒト多能性幹細胞の約20%が癌関連の変異を持っています [7]. したがって、回路が幹細胞由来のラインに入る場合、誘導可能な成長停止または除去モジュールを含めるべきです。バニリン酸 はここで優先すべき有用な誘導剤です。なぜなら、それは認可された食品添加物であり、培養肉細胞ラインでの回路トリガーとして使用するケースを助けるからです [1].
「合成生物学は、エンジニアに複数の遺伝子の発現を簡単かつ正確に調整するための回路を展開する手段を提供し、潜在的な悪影響のあるオフターゲット副作用を排除します。" - npj Systems Biology and Applications [6]
これらの選択は、第3節で系統特異的回路を設定します。
sbb-itb-ffee270
3. 筋原性、脂肪原性、および比率制御された分化のための回路戦略
3.1 拡張と終末分化を分離する筋原性回路
回路アーキテクチャが設定されると、次の仕事は系統特異的な展開です。筋形成において、主な問題は簡単に述べることができますが、実行するのは難しいです:細胞は最初に増殖し、要求されたときに分化に切り替える必要がありますが、どちらの方向にも早すぎて漂流しないようにします。
A Tet-On-MyoDカセット はこれを行う最も直接的な方法の1つです。この設定では、ドキシサイクリン(DOX)が存在しない標準条件下で細胞が増殖します。DOXを追加すると、回路が筋原性コミットメントを促進します。複数の中国の研究機関の研究者たちは、このアプローチを鶏胚線維芽細胞で使用し、誘導後に効率的な筋管形成を報告しました[4].
双安定トグルスイッチは、より厳密な状態制御を提供します。E-KRABやPip-KRABのような相互抑制リプレッサーから構築されたシステムは、短いDOXパルスを安定した筋原性プログラムに変えることができます[6]. 実際には、これは誘導後に分化状態がロックされ、信号が除去された後も消えないことを意味します。負の自己調節を追加することも、拡張フェーズ中の確率的ノイズを削減し、漏れ分化を制限するのに役立ちます[6] .
これは重要です。なぜなら、標準的なin vitro筋原性分化は途中で停滞することが多いからです。融合指数は通常50〜60%程度であり、大きな非融合予備集団が残ります[3]. 回路駆動型MyoD活性化はコミットメントを改善できますが、コミットメントだけでは均一な融合を保証しません。MyoD回路がMEK、NOTCH、RXRの調節と組み合わさると、2D培養でほぼ完全な融合が可能になります[3]. 回路は系統のエントリーのタイミングを制御し、小分子はよりクリーンで均一な細胞周期の終了を促進します。
3.2 組成と構造のための脂肪生成および線維生成の制御
筋肉だけでは不十分です。筋原性制御が確立されたら、次の課題は組成です:どれだけの脂肪が形成されるか、どれだけのECMが沈着するか、そしてそれらのプログラムがいつオンになるかです。ここでは、バイナリのON/OFF制御はしばしば粗すぎます。チームが通常必要とするのは、特に脂肪生成-線維生成の分岐点周辺での段階的な出力です。
miSFITsは段階的に発現を調整する実用的な方法を提供します。変異したmiRNAターゲットサイト、例えばmiR-17のサイトをPPARGやBMP4のような出力遺伝子の3′UTRに配置することで、研究者はバリアントライブラリから発現レベルを選択できます[5]. それにより、脂肪細胞の誘導はライトスイッチよりも調光スイッチに近くなります。細胞を全か無かの反応に押し込む代わりに、チームは脂肪生成をより慎重に調整できます[5].
線維芽細胞もここでは単なる傍観者ではありません。彼らはテクスチャを形作るECMタンパク質を供給します[10]. それにより、線維形成制御は単なる副次的な問題ではなく、製品設計の一部となります。回路は線維形成状態と脂肪形成状態の間のシフトを管理するのに役立ち、培養された家禽では、線維芽細胞におけるPPARGの直接活性化が意味のある脂肪沈着を生成するために必要かもしれません[10].
ボウタイアーキテクチャは、この問題に適しており、センシングと出力を分離します。センシング層は細胞の現在の状態を読み取り、出力層はPPARG、CEBPA、または他の系統調節因子を調整します。この分離により、細胞が適切な発達段階に達する前に脂肪生成や結合組織プログラムがオンになるのを防ぎます。
3.3 マルチ系統比率制御とフィードバックセンシング
比率制御回路は異なる問題に対処します。それは分化が起こるかどうかではなく、最終的な集団の混合が適切な場所に留まるかどうかです。ホールカット製品では、筋肉、脂肪、ECMを適切な割合にすることが、どの系統を分化させるかと同じくらい重要です。
これらのシステムは、フィードバック制御を細胞自体に組み込みます。状態特異的プロモーターは、既に特定の系統にコミットした細胞にシグナル伝達タンパク質の発現を制限します。合成パラクリンモジュールは、コミットされた筋原性細胞が抑制信号を放出し、近隣の細胞における脂肪生成のコミットメントを抑制します。このロジックは、デルタ-ノッチ発生システムにおける側方抑制に似ています[1][6]. 分岐がより複雑になる場合、マルチインプットゲートは細胞外の手がかりと内部状態信号を組み合わせることができます [9].
miSFITsもこの段階で機能します。BMP4や他のモルフォゲンの出力強度を調整することで、チームは上流の意思決定ロジックを書き換えることなく系統バランスをシフトできます。エンジニアリングされた遺伝子調節ネットワークでは、このタイプの制御により、ターゲット系統の分化効率がコントロールの52%からエンジニアリングされたiPSCsで81%に増加しました[2]. 修正された間葉系幹細胞では、合成回路が心臓分化効率を76%に向上させました[2].
以下の表は、生産において最も重要なポイントを使用して、単一系統アプローチと比率制御アプローチを比較しています。
| 特徴 | 単一系統回路 | 比率制御回路 |
|---|---|---|
| 複雑さ | 低; 通常は単一の誘導性プロモーターとレギュレーター[4] | 高; 論理ゲートとパラクリンシグナルが必要[6][9] |
| 監視負担 | 低; 通常は単一のレポーターに従う[4] | 高; 複数の系統マーカーの追跡が必要[5] |
| 堅牢性 | 中程度; 異質性と予備細胞形成に弱い[3] | 高; フィードバックと側抑制を使用して集団をバランスさせる [1] |
| 生産価値 | バイオマスには高い; 複雑な組織構造には限られている [10] | 筋肉、脂肪、ECMを必要とするホールカット製品に不可欠 [4] |
比率制御回路はより重い検証負荷を追加します。 しかし、彼らの組み込みフィードバックは、プロセス制御だけでは一致させるのが難しく、クローン選択とプロセステストにより多くのプレッシャーをかけます。
4. 構築からプロセスへ: 検証、スケールアップ、規制適合
4.1 安定したパフォーマンスのための統合戦略とクローン選択
回路設計の後、難しい部分が始まります: その回路を生産規模の継代を通じて安定した状態で細胞に導入することです。
レンチウイルスデリバリーはしばしば効率的で、安定したインテグラントを迅速に提供します。しかし、統合はランダムです。それは、より多くの規制上の注意が必要であり、サイレンシングによって発現が時間とともに低下するリスクがあることを意味します。トランスポゾンシステム、例えばPiggyBacやSleeping Beautyは中間に位置します。これらは多くの継代を通じてパフォーマンスを維持できますが、コピー数と挿入部位をスクリーニングする必要があります。眠れる森の美女 , 例えば、TERTとCDK4を過剰発現させることにより、ウシ衛星細胞を安定的に不死化するために使用されており、40回以上の継代後も筋原性の可能性を保持しています [10]. CRISPRノックインは、構造が着地する場所を最も厳密に制御し、最も正確なゲノム設定を提供しますが、クローン選択は遅く、スループットは低くなります。
| 統合方法 | 挿入制御 | 安定性 | スケーラビリティ | 規制上の考慮事項 |
|---|---|---|---|---|
| レンチウイルスデリバリー | 低い(ランダム統合) | 高いがサイレンシングの可能性あり | 高い | ランダム挿入とウイルス残留物による規制の厳格化 |
| トランスポゾン(PiggyBac/SB) | 中程度 | 多くのパッセージで高い | 高い | コピー数と挿入部位のスクリーニングが必要 |
| CRISPRノックイン | 高い(部位特異的) | 非常に高い | 中程度 | 好ましい;内因性遺伝子の破壊リスクを低減 |
| エピソーマルベクター | なし(染色体外) | 低い;分裂中に失われる可能性あり | 低い | 統合のハードルは低いが、長期的な拡張には不向き |
クローンのスクリーニングは、構築物が存在することを確認する以上のことを行う必要があります。それは、発現ドリフト、挿入プロファイル、成長動態、分化効率、および高継代数での表現型保持を追跡する必要があります。snRNA-seqは、Pax7⁺/Ki-67⁻予備細胞に富むクローンをスケールアップ前に除外できるため、ここで有用です - 分化せずに細胞周期を離れる細胞です - [3]. Tet-On-PAX7回路を持つブタEPSCsは、3Dマイクロキャリアおよび懸濁培養で40継代以上にわたり高い筋分化を保持しました[8].
4.2 3D、マイクロキャリアおよびバイオリアクター培養における回路挙動の変化
クローンを得たら、次のテストはそれが2Dの外で同じように振る舞うかどうかです。多くの場合、そうではありません。2Dでのパフォーマンスは、懸濁、マイクロキャリア、またはスキャフォールド培養にきれいに引き継がれることはめったにありません。なぜなら、拡散勾配、酸素制限、およびせん断がすべて回路出力を変化させるからです。
最初のチェックの一つはインデューサー拡散です。撹拌槽型バイオリアクターでは、小分子インデューサーが均等に細胞に到達する必要があります。実際には、特に高密度のマイクロキャリア培養や凝集体や足場のコア内部で勾配が形成されることがあります。懸濁培養は通常、大規模な培養肉に適しており、より高い細胞密度をサポートし、プロセス制御を厳密に行うことができます。
システムが拡大するにつれて、細胞状態のモニタリングも難しくなります。2Dで顕微鏡で簡単に読み取れる蛍光レポーター信号は、不透明な3D構造では解決が難しくなることがあります。蛍光タイマー - タンパク質が成熟するにつれて発光が青から赤に変わるプローブ - は、in situでリアルタイムの回路活性化データを提供できます。[1]. 賢明なルートは段階的な検証です:最初に2Dで、次に3D構造で、そして最終的なバイオリアクター条件下で行います [3][8].
4.3 特性評価、食品安全文書および英国/EUの考慮事項
プロセステストの後、特性評価は回路機能、表現型、安全性が依然として維持されていることを示さなければなりません。コアデータスタックには、フローサイトメトリー、cm配列を用いたqPCR、RNA-seqタイムコース、およびミオシン重鎖面積分率やミオグロビン発現などの機能的読み出しが含まれるべきです [1]. 最適化された無血清分化培地は、ミオグロビン発現をネイティブの牛筋肉で見られるレベルの約30%まで引き上げることが示されています [3]. それにより、チームは漠然とした目標ではなく、明確なベンチマークを得ることができます。
タンパク質、アミノ酸、脂肪のプロファイルと感覚特性を文書化する必要もあります[10][3].
規制の観点から、英国とEUは自発的に不死化された(非GMO)細胞株と遺伝子工学的に改変された細胞株を明確に区別しています。後者はより広範な安全性の書類が必要です[10][3]. 安定性パッケージは、マスターセルバンクから最終生産細胞までの完全な生産ブリッジにわたって表現型の保持とゲノムの安定性を示す必要があり、トレーサビリティ記録はその間のすべてのパッセージを考慮する必要があります[10]. 回路が化学誘導体に依存する場合、バニル酸のような食品安全または認可された添加物がドキシサイクリンよりも望ましいです[1].
ルーチンのゲノム監視は必須であり、誘導可能な自殺または除去スイッチは、コアのリスク管理措置として文書化されるべきです[7]. その機能は安全性に関する書類にも含まれるべきであり、特に英国とEUの培養肉に関する規則が形成され続ける中で重要です。
5. 実用的なロードマップと結論
5.1 培養肉チームのための段階的な実施ロードマップ
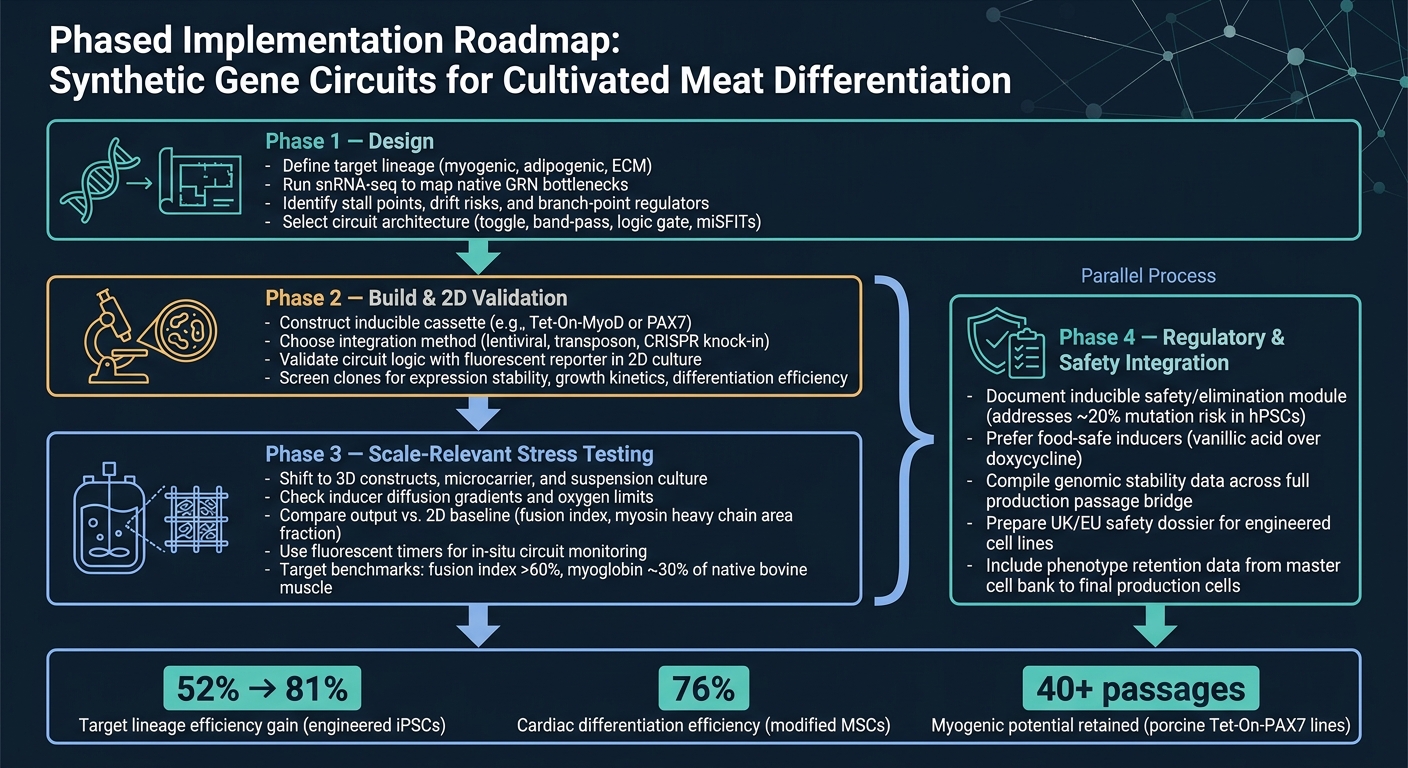
コンセプトから生産までの最もクリーンなルートは段階的なワークフローです。
フェーズ1は設計です。ターゲット系統を定義することから始め、snRNA-seqを使用して主なボトルネックを確認し、回路アーキテクチャを選択します。このステップは重要であり、回路は実際に特定した制約しか解決できないからです。
フェーズ2は構築と2D検証です。構築物を作成し、シンプルなレポーターリードアウトを使用して回路が意図した通りに動作するかを2Dで確認します。この段階では、目標は単純です:より困難で高価なモデルに移行する前に、ロジックが機能することを確認します。
フェーズ3は、スケールに関連したストレステストです。3Dシステムとバイオリアクターに関連する条件に移行し、2Dベースラインと出力を比較します。ここで、多くのデザインが弱点を示し始めます。特に、物質移動、せん断、マトリックス効果が関与する場合です。
フェーズ4は、規制および安全性の統合であり、フェーズ3と並行して実行する必要があります。安全性と規制の作業は、最後まで待つべきではありません。スケールアップと並行して実行し、誘導可能な安全モジュールのドキュメントを含めます。
5.2 Cellbase を通じたツールと材料の調達
ワークフローが設定されると、調達がしばしば速度制限ステップになります。
- 細胞株
- 無血清および化学的に定義された培地
- 足場
- バイオリアクターコンポーネント
- センサー
- 分析機器
各段階での互換性のある材料への信頼できるアクセスは、スケールに関連する条件下で回路の挙動をどれだけ速く特性化できるかに直接影響します。
5.3 重要なポイント
合成遺伝子回路は、培養肉チームにメディアのみのプロトコルでは実現できないタイミング、閾値、系統バランスのプログラム可能な制御を提供します。アーキテクチャの選択は、可逆性、漏れやすさ、安全性を形作ります。誘導システムは、条件付き制御と低い代謝負荷を提供するため、通常は好まれます[6].
「合成生物学ツールキットは、調整可能な遺伝子発現を持つ細胞株を確立するために使用でき、PATおよび計算モデリングと組み合わせることで、最適な製品収量と品質を提供する閉ループ制御システムを可能にします。」 - npj Systems Biology and Applications [6]
成功した展開は、生物学の問題だけではありません。それは、回路工学、バイオプロセス設計、規制文書、および調達の間の緊密な連携に依存しています。
よくある質問
合成遺伝子回路はどのように分化の一貫性を向上させますか?
合成遺伝子回路は、細胞の挙動と系統のコミットメントに対するプログラムされた制御を提供するため、分化をより一貫性のあるものにすることができます。実際には、モジュール式の論理操作を使用して、遺伝子と転写因子の発現を厳密なタイミングで調整することを意味します。
そのタイミングが重要です。これにより、細胞が混合または望ましくない状態に漂うのではなく、正しい順序で定義された状態変化を通過するのを助けます。また、ターゲット外の分化を減少させ、培養全体のノイズを減らします。
その結果は明白です:培養肉生産のためのより均一で安定した成熟した細胞集団。
筋原性または脂肪原性制御に適した回路設計はどれですか?
培養肉研究では、同じ鶏の線維芽細胞をどちらの系統にも押し進めることができます。筋形成は一連の誘導プロトコルに従い、脂肪形成は鶏血清や脂肪酸などの入力に細胞をさらすことでオンにすることができます。
そこから、これらの細胞運命は、3Dハイドロゲル足場内で段階的に制御され、定義された脂肪とコラーゲンの比率を持つ肉構造を構築することができます。
なぜ遺伝子回路は3D培養でしばしば異なる動作をするのですか?
3D培養 では、遺伝子回路はしばしば異なる動作をします。なぜなら、細胞は2D単層には存在しない物理的および構造的入力に対処しているからです。これらの入力には、機械的張力、せん断応力、マトリックスの硬さ、局所的な細胞密度が含まれます。
これらの手がかりは、Notchのようなシグナル伝達経路を変えることができます。また、合成回路が力を検出し、細胞間接着や組織形態形成を含む下流の応答を調整する方法を変えることもできます。









