

Se você pode expandir células, mas não consegue direcioná-las para o destino certo no momento certo, seu processo irá estagnar na diferenciação. Esse é o ponto principal aqui: circuitos genéticos sintéticos oferecem controle dentro da célula sobre compromisso, tempo, memória e mistura de linhagem, onde apenas mudanças de meio frequentemente deixam populações heterogêneas, parcialmente comprometidas.
Se eu estivesse construindo um fluxo de trabalho de diferenciação de carne cultivada, eu tiraria quatro pontos deste artigo imediatamente:
- Comece com a rede nativa, não com o construto. Use análise de trajetória snRNA-seq, , inferência de GRN e perfil de miRNA para encontrar onde as células estagnam, desviam ou ramificam para o destino errado.
-
Combine o tipo de circuito com o problema do processo.
Um interruptor de alternância é adequado para bloqueio, um design de feedforward ou passa-faixa é adequado para controle de tempo, uma porta lógica é adequada para controle de múltiplos sinais, e miSFITs são adequados para saída graduada. - Projete para baixo vazamento, baixo ruído e segurança desde o primeiro dia. Peças ortogonais, autorregulação negativa, iFFLs, transgenes cm e um módulo de morte ou parada de crescimento induzível fazem parte da construção, não são uma reflexão tardia.
- Valide sob condições relevantes para a escala desde cedo. Um circuito que funciona em 2D pode mudar em 3D, microcarrier ou suspensão agitada devido a gradientes de indutor, limites de oxigênio e cisalhamento.
O artigo também faz um ponto prático que importa para as equipes de processo: controle de linhagem única e controle de proporção são trabalhos diferentes. Uma cassete Tet-On MyoD pode impulsionar a entrada miogênica, mas produtos de corte inteiro precisam de controle das proporções de músculo, gordura e ECM, o que geralmente significa feedback, sinalização parácrina e triagem mais rigorosa de clones.
Algumas figuras ancoram essa mensagem:
- A diferenciação miogênica padrão pode estagnar com índices de fusão de aproximadamente 50–60%
- GRNs projetados em iPSCs aumentaram a diferenciação da linhagem alvo de 52% para 81%
- Circuitos sintéticos em MSCs modificados impulsionaram a diferenciação cardíaca para 76%
- Algumas linhas porcina Tet-On-PAX7 mantiveram alto potencial miogênico além de 40 passagens
- Cerca de 20% das células-tronco pluripotentes humanas podem carregar mutações ligadas ao câncer, por isso os módulos de segurança induzíveis são importantes
Fluxo de Trabalho de Circuito Genético Sintético para Diferenciação de Carne Cultivada
Histórias de Cientistas: Michael Elowitz, Circuitos Genéticos em Células Vivas
Comparação rápida
| Tópico | O que o artigo diz em termos simples |
|---|---|
| Principal gargalo | Diferenciação, não expansão |
| Por que os circuitos ajudam | Eles adicionam controle de limiar e memória de estado celular dentro da célula |
| Melhor primeiro passo | Mapear controle de destino nativo com snRNA-seq e ferramentas relacionadas |
| Principais opções de circuito | Toggle, feedforward, band-pass, portas lógicas, miSFITs, CRISPRa/i |
| Exemplo de miogênese | Tet-On-MyoD para separar crescimento de diferenciação terminal |
| Adipogênese / controle de ECM | miSFITs e designs bow-tie para saída graduada do tipo PPARG/BMP4 |
| Desafio de corte total | Controle de proporção entre músculo, gordura e tecido conjuntivo |
| Risco de ampliação | O comportamento 2D pode não se manter em 3D ou biorreatores |
| Escolhas de integração | Lentivírus, transposons, CRISPR knock-in, vetores episomais |
| Ponto regulatório | Linhas engenheiradas precisam de um pacote de segurança mais amplo; indutores seguros para alimentos, como ácido vanílico, são preferidos em relação ao DOX sempre que possível |
Então, em termos simples: isto não é apenas um artigo de design de circuitos. Eu leria isso como um guia para vincular arquitetura de circuitos, biologia de linhagem, seleção de clones, desempenho de biorreatores e documentação de segurança do Reino Unido/UE em uma estratégia de diferenciação.
Continue lendo se você deseja o caminho completo desde o mapeamento da rede nativa até a escolha da construção, verificações de escala e adequação regulatória.
2. Princípios de design para circuitos de controle de diferenciação
2.1 Mapeie a rede de destino celular nativa antes de projetar o circuito
Antes de projetar um circuito, você precisa de uma imagem clara do que a célula já está fazendo.
Sequenciamento de RNA de núcleo único (snRNA-seq) é um bom ponto de partida. Ele pode mostrar subpopulações quiescentes, incluindo células de reserva marcadas por NOTCH2 e HEYL , e apontar para alvos de vias que podem melhorar a diferenciação [3].
A partir daí, a análise de trajetória e a inferência da rede regulatória gênica (GRN) ajudam a mapear a ordem de ativação dos reguladores e destacam onde as células têm maior probabilidade de estagnar. Na miogênese, a cascata principal passa por MYOD1 e MYOG. Na adipogênese, os principais nós são PPARG e CEBPA, com o destino do progenitor fibro-adipogênico (FAP) atuando como o principal ponto de risco de ramificação. A tabela abaixo resume os principais reguladores e gargalos.
| Linhagem | Principais reguladores mestres | Caminhos de sinalização críticos | Gargalos identificados |
|---|---|---|---|
| Miogênico | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Formação de células de reserva (quiescência) |
| Adipogênico | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Destino de progenitores fibro-adipogênicos (FAP) |
| Pluripotente | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Diferenciação espontânea / heterogeneidade |
Outra camada útil é perfil de expressão de miRNA. miRNAs endógenos como miR-302a, ligados à pluripotência, e miR-375, ligados à diferenciação, podem atuar como classificadores internos em designs de sentido e resposta. Isso permite que o circuito leia o estado real da célula em vez de depender apenas de indutores externos [5].
Esses gargalos devem orientar a escolha do circuito. Se o principal problema for desvio, pode ser necessário um lock-in. Se o problema for o tempo, um pulso pode se ajustar melhor. Se o controle do destino depender de mais de uma pista, a lógica de múltiplas entradas geralmente faz mais sentido.
2.2 Escolha a arquitetura de circuito correta
É aqui que os compromissos aparecem claramente. A arquitetura correta depende de três pontos práticos: quão permanente o compromisso deve ser, quão rigorosamente o tempo precisa ser controlado e quanto de carga genética a célula pode carregar sem problemas.
Chaves de alternância biestáveis se encaixam em casos onde o compromisso de linhagem precisa permanecer bloqueado. O principal problema é a inversão espontânea de estado causada por ruído transcricional.
Filtros passa-faixa se encaixam em casos onde um fator de transcrição é necessário apenas dentro de uma janela de desenvolvimento definida. O problema é que os níveis de indutor precisam de controle rigoroso, ou o tempo escapa.
Portas lógicas adicionam especificidade ao exigir mais de uma entrada ao mesmo tempo. Por exemplo, a diferenciação pode ser permitida apenas quando um indutor exógeno está presente e a célula mostra o perfil correto de miRNA endógeno. Isso ajuda a reduzir o risco de comprometimento fora do alvo.
A tabela abaixo descreve as principais arquiteturas e os compromissos que vêm com elas.
| Arquitetura | Reversibilidade | Precisão temporal | Complexidade de integração | Uso principal | Risco chave |
|---|---|---|---|---|---|
| Interruptor biestável | Baixo (uma vez travado) | Moderado | Moderado | Comprometimento de linhagem permanente | Inversão espontânea devido ao ruído |
| Filtro passa-faixa | Alto (dependente de concentração) | Alto | Alto | Estágios de desenvolvimento transitórios | Requer controle rigoroso do indutor |
| Porta lógica (AND/OR/NOT) | Variável | Moderado | Moderado–alto | Ativação específica de tipo celular | Vazamento em estados OFF |
| Gravata borboleta / entrada múltipla | Alto | Moderado | Moderado | Integração de múltiplos sinais | Depende da estabilidade endógena de miRNA |
| miSFITs | Alto | Moderado | Baixo–moderado | Ajuste de saída graduado | Faixa dinâmica estreita se mal ajustada |
"Ao minimizar o número de camadas computacionais enquanto mantém a funcionalidade, essa estratégia aborda barreiras de escalabilidade na engenharia de circuitos genéticos." - Nature Communications [9]
Cada camada regulatória adicionada aumenta a carga útil e consome recursos celulares. Na prática, se dois designs fazem o mesmo trabalho, o mais simples geralmente é a melhor aposta quando a escala importa.
Uma vez que a arquitetura está definida, o próximo trabalho é fazê-la resistir a baixa vazão, supressão de ruído e controle à prova de falhas.
2.3 Construa para confiabilidade, baixa vazão e segurança
Um circuito precisa permanecer estável durante o cultivo prolongado. Um curto período de bom desempenho não é suficiente para uso em produção.
Partes ortogonais são a primeira defesa. Promotores, fatores de transcrição e elementos regulatórios que não reagem cruzadamente com a maquinaria nativa ajudam a limitar efeitos fora do alvo e reduzem a chance de que sinais endógenos ativem ou desativem o circuito.Promotores modificados de alta rigidez, como PCREm, têm sido usados para reduzir a vazão basal em sistemas mamíferos induzíveis [6] .
A autorregulação negativa também vale a pena adicionar onde possível. É um dos motivos mais conhecidos para reduzir o ruído transcricional e produzir uma resposta mais linear à concentração do indutor [6]. Os loops de alimentação direta incoerente (iFFLs) podem adicionar outra camada de controle, filtrando flutuações estocásticas, para que as células respondam a sinais sustentados em vez de picos de ruído curtos.
Versões modificadas por códons (cm ) de fatores de transcrição sintéticos também facilitam a caracterização. Elas permitem separar a expressão dirigida por circuitos da expressão genômica endógena (g) durante a validação [1]. Isso pode parecer um detalhe pequeno, mas economiza tempo quando você está tentando descobrir se uma leitura vem do circuito ou do genoma hospedeiro.
Módulos de segurança são necessários. Cerca de 20% das células-tronco pluripotentes humanas carregam mutações associadas ao câncer [7]. Portanto, se um circuito está sendo inserido em uma linha derivada de células-tronco, ele deve incluir um módulo de parada de crescimento ou eliminação induzível. Ácido vanílico é um indutor útil a ser priorizado aqui porque é um aditivo alimentar licenciado, o que ajuda no caso de usá-lo como um gatilho de circuito em linhas celulares de carne cultivada [1].
"A biologia sintética fornece aos engenheiros os meios para implantar circuitos para ajustar fácil e precisamente a expressão de múltiplos genes para... eliminar potenciais efeitos colaterais adversos fora do alvo." - npj Systems Biology and Applications [6]
Essas escolhas configuram os circuitos específicos de linhagem na Seção 3.
sbb-itb-ffee270
3. Estratégias de circuitos para diferenciação miogênica, adipogênica e controlada por proporção
3.1 Circuitos miogênicos que separam a expansão da diferenciação terminal
Uma vez que a arquitetura do circuito está definida, o próximo trabalho é o desenvolvimento específico de linhagem. Para a miogênese, o principal problema é simples de enunciar, mas difícil de executar: as células precisam proliferar primeiro, depois mudar para a diferenciação quando solicitado, sem desviar muito cedo em qualquer direção.
Um cassete Tet-On-MyoD é uma das maneiras mais diretas de fazer isso. Nesta configuração, as células proliferam sob condições padrão quando a doxiciclina (DOX) está ausente. Adicione DOX, e o circuito impulsiona o compromisso miogênico.Pesquisadores de várias instituições chinesas usaram essa abordagem em fibroblastos embrionários de frango e relataram formação eficiente de miotubos após indução [4].
Um interruptor de alternância bistável oferece um controle de estado mais rigoroso. Sistemas construídos a partir de repressores mutuamente inibitórios, como E-KRAB e Pip-KRAB, podem transformar um pulso curto de DOX em um programa miogênico estável [6]. Na prática, isso significa que o estado diferenciado permanece bloqueado após a indução, em vez de desaparecer uma vez que o sinal é removido. A adição de autorregulação negativa também ajuda a reduzir o ruído estocástico e limitar a diferenciação vazada durante a fase de expansão [6].
Isso é importante porque a diferenciação miogênica padrão in vitro muitas vezes para no meio do caminho. Os índices de fusão geralmente são apenas em torno de 50–60%, o que deixa uma grande população de reserva não fundida [3]. A ativação do MyoD dirigida por circuitos pode melhorar o comprometimento, mas o comprometimento por si só não garante uma fusão uniforme. Quando os circuitos MyoD são combinados com a modulação de MEK, NOTCH e RXR, a fusão quase completa em cultura 2D se torna possível [3]. O circuito controla o tempo de entrada na linhagem; as pequenas moléculas promovem uma saída do ciclo celular mais limpa e uniforme.
3.2 Controle adipogênico e fibrogênico para composição e estrutura
Somente o músculo não é suficiente. Uma vez que o controle miogênico está em vigor, a próxima questão é a composição: quanto de gordura se forma, quanto de ECM é depositado e quando esses programas são ativados. Aqui, o controle binário LIGADO/DESLIGADO é muitas vezes muito brusco. O que as equipes geralmente precisam é de uma saída graduada, especialmente em torno do ponto de ramificação adipogênico-fibrogênico.
miSFITs oferecem uma maneira prática de ajustar a expressão em etapas.Ao colocar sítios-alvo de miRNA mutados - por exemplo, sítios para miR-17 - no 3′UTR de genes de saída como PPARG ou BMP4, os pesquisadores podem selecionar níveis de expressão de uma biblioteca de variantes [5]. Isso torna a indução de adipócitos mais semelhante a um dimmer do que a um interruptor de luz. Em vez de empurrar as células para uma resposta de tudo ou nada, as equipes podem titular a adipogênese com mais cuidado [5].
Os fibroblastos também não são apenas espectadores aqui. Eles fornecem proteínas da ECM que moldam a textura [10]. Isso torna o controle fibrogênico parte do design do produto, não apenas uma questão secundária. Os circuitos podem ajudar a gerenciar a mudança entre estados fibrogênicos e adipogênicos, e em aves cultivadas, a ativação direta de PPARG em fibroblastos pode ser necessária para gerar uma deposição de gordura significativa [10].
Uma arquitetura de gravata borboleta se adapta bem a este problema porque mantém a detecção e a saída separadas. A camada de detecção lê o estado atual da célula, enquanto a camada de saída ajusta PPARG, CEBPA ou outros reguladores de linhagem. Essa separação ajuda a impedir que programas adipogênicos ou de tecido conjuntivo sejam ativados antes que as células alcancem o estágio de desenvolvimento correto.
3.3 Controle de proporção multi-linhagem e detecção de feedback
Circuitos de controle de proporção lidam com um problema diferente: não se a diferenciação acontece, mas se a mistura final da população permanece onde deveria. Para produtos inteiros, obter músculo, gordura e ECM nas proporções corretas é tão importante quanto fazer qualquer linhagem se diferenciar.
Esses sistemas incorporam controle de feedback nas próprias células. Promotores específicos de estado restringem a expressão de proteínas sinalizadoras a células que já se comprometeram com uma determinada linhagem.Módulos parácrinos sintéticos então permitem que células miogênicas comprometidas liberem um sinal inibitório que suprime o comprometimento adipogênico em células próximas. A lógica se assemelha à inibição lateral em sistemas de desenvolvimento Delta-Notch [1][6] . Onde o ramificação se torna mais complicada, portões de múltiplas entradas podem combinar sinais extracelulares com sinais de estado interno [9].
miSFITs também funcionam nesta etapa. Ao ajustar a força de saída de BMP4 ou outros morfógenos, as equipes podem alterar o equilíbrio de linhagem sem reescrever a lógica de decisão a montante. Em redes regulatórias gênicas projetadas, esse tipo de controle aumentou a eficiência de diferenciação da linhagem alvo de 52% nos controles para 81% em iPSCs projetadas [2]. Em células-tronco mesenquimais modificadas, circuitos sintéticos impulsionaram a eficiência de diferenciação cardíaca para 76% [2].
A tabela abaixo compara abordagens de linhagem única e controle de proporção usando os pontos que mais importam na produção.
| Recurso | Circuito de linhagem única | Circuito de controle de proporção |
|---|---|---|
| Complexidade | Baixa; tipicamente um único promotor e regulador induzível [4] | Alta; requer portas lógicas e sinalização parácrina [6][9] |
| Encargo de monitoramento | Baixo; geralmente segue um único repórter [4] | Alto; requer rastreamento de múltiplos marcadores de linhagem [5] |
| Robustez | Moderada; propensa a heterogeneidade e formação de células de reserva [3] | Alto; usa feedback e inibição lateral para equilibrar populações [1] |
| Valor de produção | Alto para biomassa; limitado para estrutura de tecido complexo [10] | Essencial para produtos inteiros que precisam de músculo, gordura e ECM [4] |
Circuitos de controle de proporção adicionam uma carga de validação mais pesada. Mas o feedback embutido é difícil de igualar apenas com o controle de processo, o que coloca mais pressão na seleção de clones e nos testes de processo.
4. Do constructo ao processo: validação, ampliação e adequação regulatória
4.1 Estratégia de integração e seleção de clones para desempenho estável
Após o design do circuito, a parte difícil começa: inserir esse circuito nas células de uma forma que permaneça estável durante a passagem em escala de produção.
Entrega lentiviral é frequentemente eficiente e fornece integradores estáveis rapidamente. Mas a integração é aleatória. Isso significa mais atenção regulatória, além de um risco de que a expressão diminua ao longo do tempo devido ao silenciamento. Sistemas de transposons como PiggyBac e Sleeping Beauty estão no meio. Eles podem manter o desempenho através de muitas passagens, mas ainda é necessário verificar o número de cópias e os locais de inserção.Sleeping Beauty , por exemplo, tem sido usado para imortalizar de forma estável células satélites bovinas por meio da superexpressão de TERT e CDK4, com linhas mantendo potencial miogênico além de 40 passagens [10] . CRISPR knock-in oferece o controle mais rigoroso sobre onde o constructo se insere e o ajuste genômico mais preciso, embora a seleção de clones seja mais lenta e o rendimento seja menor.
| Método de Integração | Controle de Inserção | Estabilidade | Escalabilidade | Considerações Regulatórias |
|---|---|---|---|---|
| Entrega Lentiviral | Baixa (integração aleatória) | Alta, mas propensa a silenciamento | Alta | Maior escrutínio regulatório devido à inserção aleatória e remanescentes virais |
| Transposons (PiggyBac/SB) | Moderada | Alta em muitas passagens | Alta | Requer triagem para número de cópias e locais de inserção |
| CRISPR Knock-in | Alta (específica para o local) | Muito alta | Moderada | Favorável; reduz o risco de interromper genes endógenos |
| Vetores Episomais | Nenhum (extracromossômico) | Baixo; pode ser perdido durante a divisão | Baixo | Menor obstáculo de integração, mas inadequado para expansão a longo prazo |
A triagem de clones precisa fazer mais do que confirmar a presença do constructo.Deve rastrear a deriva de expressão, perfil de inserção, cinética de crescimento, eficiência de diferenciação e retenção de fenótipo em número de passagem elevado. snRNA-seq é útil aqui porque pode descartar clones enriquecidos para células de reserva Pax7⁺/Ki-67⁻ - células que saem do ciclo celular sem se diferenciar - antes da ampliação [3]. EPSCs suínas com um circuito Tet-On-PAX7 mantiveram alta diferenciação muscular em cultura de microcarregador 3D e suspensão por mais de 40 passagens [8].
4.2 Como o comportamento do circuito muda em cultura 3D, microcarregador e biorreator
Uma vez que você tem um clone, o próximo teste é se ele se comporta da mesma forma fora do 2D. Em muitos casos, não se comporta. O desempenho em 2D raramente se transfere de forma limpa para suspensão, microcarregador ou cultura de suporte porque gradientes de difusão, limites de oxigênio e cisalhamento alteram a saída do circuito.
Um dos primeiros testes é a difusão do indutor. Em biorreatores de tanque agitado, indutores de pequenas moléculas precisam alcançar as células de maneira uniforme. Na prática, podem se formar gradientes, especialmente em culturas densas de microcarregadores e dentro de agregados ou núcleos de suportes. A cultura em suspensão geralmente é mais adequada para carne cultivada em larga escala porque suporta maior densidade celular e oferece um controle de processo mais rigoroso.
Monitorar o estado das células também se torna mais difícil à medida que o sistema escala. Sinais de repórteres fluorescentes que são fáceis de ler por microscopia em 2D podem se tornar difíceis de resolver em construções opacas em 3D. Temporizadores fluorescentes - sondas que mudam a emissão de azul para vermelho à medida que uma proteína amadurece - podem fornecer dados de ativação de circuito em tempo real in situ [1]. A rota sensata é a validação em etapas: primeiro em 2D, depois em construções 3D, e então sob as condições finais do biorreator [3][8] .
4.3 Caracterização, documentação de segurança alimentar e considerações do Reino Unido/UE
Após os testes de processo, a caracterização deve mostrar que a função do circuito, o fenótipo e a segurança ainda se mantêm. O conjunto de dados principal deve incluir citometria de fluxo, qPCR com sequências cm, cursos de tempo de RNA-seq e leituras funcionais, como fração de área da cadeia pesada de miosina e expressão de mioglobina [1]. Mídias de diferenciação otimizadas sem soro demonstraram trazer a expressão de mioglobina para aproximadamente 30% dos níveis encontrados no músculo bovino nativo [3]. Isso dá às equipes um ponto de referência claro em vez de um alvo vago.
Você também precisa documentar perfis de proteína, aminoácidos e gorduras, juntamente com características sensoriais [10][3].
Do ponto de vista regulatório, o Reino Unido e a UE traçam uma linha clara entre linhagens celulares imortalizadas espontaneamente (não-OGM) e linhagens geneticamente modificadas. Estas últimas precisam de dossiês de segurança mais amplos [10][3]. Os pacotes de estabilidade devem mostrar a retenção do fenótipo e a estabilidade genômica ao longo de toda a ponte de produção - desde o banco de células mestre até as células de produção final - e os registros de rastreabilidade precisam contabilizar cada passagem intermediária [10]. Se o circuito depender de um indutor químico, um aditivo seguro para alimentos ou licenciado, como o ácido vanílico, é preferível à doxiciclina [1].
A vigilância genômica de rotina é essencial, e um interruptor de suicídio ou eliminação induzível deve ser documentado como uma medida central de controle de risco [7]. Sua função também deve ser incluída no dossiê de segurança, especialmente à medida que as regras do Reino Unido e da UE para carne cultivada continuam a se formar.
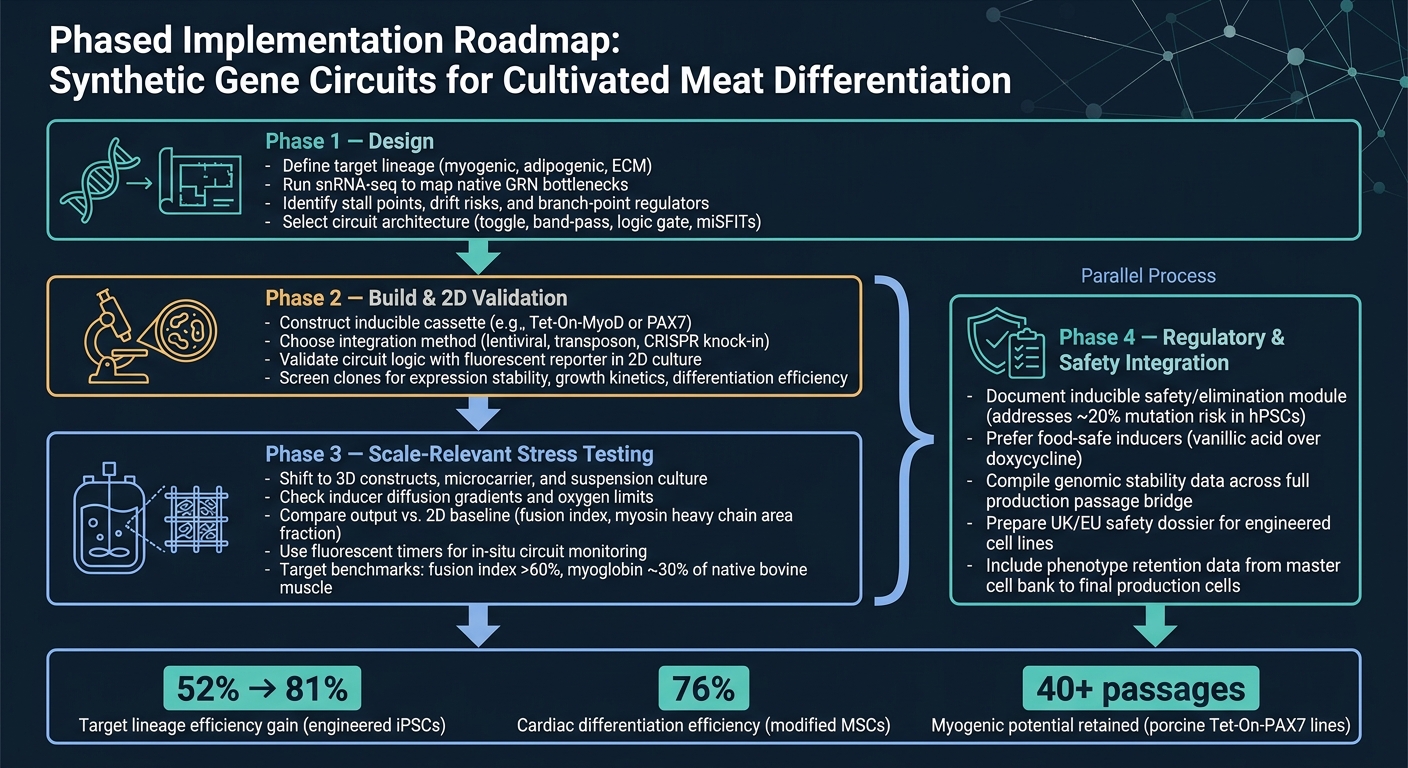
5. Roteiro prático e conclusão
5.1 Um roteiro de implementação em fases para equipes de carne cultivada
A rota mais limpa do conceito à produção é um fluxo de trabalho em etapas.
Fase 1 é o design. Comece definindo a linhagem alvo, depois use snRNA-seq para confirmar os principais gargalos antes de escolher uma arquitetura de circuito. Esse passo é importante porque um circuito só pode resolver as restrições que você realmente identificou.
Fase 2 é construção e validação 2D. Construa o projeto e verifique se o circuito se comporta conforme o esperado em 2D, usando uma leitura simples de repórter.Neste estágio, o objetivo é simples: confirmar que a lógica funciona antes de avançar para modelos mais difíceis e custosos.
Fase 3 é o teste de estresse relevante para escala. Mude para sistemas 3D e condições relevantes para biorreatores, depois compare a saída com a linha de base 2D. É aqui que muitos designs começam a mostrar seus pontos fracos, especialmente quando efeitos de transferência de massa, cisalhamento e matriz entram em jogo.
Fase 4 é a integração regulatória e de segurança, e deve ocorrer em paralelo com a Fase 3. O trabalho de segurança e regulamentação não deve esperar até o final. Execute-o junto com a ampliação, incluindo documentação para qualquer módulo de segurança induzível.
5.2 Aquisição de ferramentas e materiais habilitadores através de Cellbase
Uma vez que o fluxo de trabalho está definido, a aquisição muitas vezes se torna o passo limitante.
- linhas celulares
- meios sem soro e quimicamente definidos
- andaimes
- componentes de biorreatores
- sensores
- equipamentos analíticos
Acesso confiável a materiais compatíveis em cada estágio tem um efeito direto na rapidez com que o comportamento do circuito pode ser caracterizado em condições relevantes para escala.
5.3 Principais conclusões
Circuitos genéticos sintéticos dão às equipes de carne cultivada controle programável sobre o tempo, os limiares e o equilíbrio de linhagem que os protocolos apenas de meios não conseguem igualar. A escolha da arquitetura molda a reversibilidade, a vazão e a segurança.Sistemas induzíveis são geralmente preferidos porque oferecem controle condicional e uma carga metabólica menor [6].
"O kit de ferramentas de biologia sintética pode ser usado para estabelecer linhas celulares com expressão gênica ajustável, que, quando acopladas com PAT e modelagem computacional, podem permitir sistemas de controle em malha fechada para entregar rendimento e qualidade de produto ótimos." - npj Systems Biology and Applications [6]
O sucesso da implantação não é apenas um problema de biologia. Depende de um acoplamento estreito entre engenharia de circuitos, design de bioprocessos, documentação regulatória e aquisição.
Perguntas Frequentes
Como os circuitos genéticos sintéticos melhoram a consistência da diferenciação?
Os circuitos genéticos sintéticos podem tornar a diferenciação mais consistente porque oferecem controle programado sobre o comportamento celular e o compromisso de linhagem.Na prática, isso significa usar operações lógicas modulares para ajustar a expressão de genes e fatores de transcrição com um tempo preciso.
Esse tempo é importante. Ele ajuda as células a passarem por mudanças de estado definidas na ordem correta, em vez de derivarem para destinos mistos ou indesejados. Também reduz a diferenciação fora do alvo e diminui o ruído em toda a cultura.
O benefício é direto: populações celulares mais uniformes, estáveis e maduras para a produção de carne cultivada.
Qual design de circuito é adequado para controle miogênico ou adipogênico?
Na pesquisa de carne cultivada, os mesmos fibroblastos de frango podem ser direcionados para qualquer linhagem. Miogênese segue um conjunto de protocolos de indução, enquanto adipogênese pode ser ativada expondo as células a insumos como soro de frango ou ácidos graxos.
A partir daí, esses destinos celulares podem ser controlados passo a passo dentro de andaimes de hidrogel 3D para construir estruturas de carne com proporções definidas de gordura e colágeno.
Por que os circuitos genéticos frequentemente se comportam de maneira diferente em cultura 3D?
Em cultura 3D, os circuitos genéticos frequentemente se comportam de maneira diferente porque as células estão lidando com entradas físicas e estruturais que simplesmente não existem em monocamadas 2D. Essas entradas incluem tensão mecânica, estresse de cisalhamento, rigidez da matriz e densidade celular local.
Esses sinais podem alterar vias de sinalização como Notch. Eles também podem mudar como os circuitos sintéticos detectam força e coordenam respostas a jusante, incluindo adesão célula-célula e morfogênese tecidual.









