

אם אתה עורך קודם ובודק מאוחר יותר, תוכל לתקן שינוי לא מכוון לתוך השיבוט. הייתי שומר על תהליך עבודה פשוט: בחר בשיטת העריכה בעלת הסיכון הנמוך ביותר, שמור על חשיפה קצרה לעורך, ולאחר מכן בדוק גם אתרים מחוץ למטרה הצפויים וגם את יציבות השיבוט לפני השחרור.
עבור מהנדסי ביופרוסס, מדעני תרביות תאים וצוותי R&D של בשר מתורבת, הנקודה המרכזית היא פשוטה. מערכות CRISPR עדיין יכולות לחתוך באתרים קרובים, לעיתים עם 3–6 אי התאמות נסבלות, וטעויות אלו יכולות לעבור לשיבוטים של תאים בודדים מורחבים. המאמר מחלק את בקרת הסיכון לשלושה שלבים: לפני העריכה, במהלך העריכה, ואחרי העריכה.
הנה הרשימה המלאה במונחים פשוטים:
-
בחר את כלי העריכה בעל הסיכון הנמוך ביותר למשימה
- השתמש ב-עריכה בסיסית או עריכה ראשונית כאשר הם יכולים לבצע את העריכה ללא שבירה כפולה
- השתמש ב-מודולציה מבוססת dCas9 אם אתה זקוק רק לרגולציה גנטית
- אם אתה זקוק לנוקלאז, התחל עם גרסה Cas9 בעלת דיוק גבוה
-
נעל את חומר ההתחלה
- אשר זהות קו התאים
- בדוק מיקופלסמה
- רשום מספר מעבר
- רצף את המיקום המטרה בפועל בקו העבודה, לא רק את הגנום הייחוס
-
סנן מדריכים לפני עבודה רטובה
- השתמש בכלים מבוססי יישור ו-מבוססי ניקוד לאיתור מחוץ למטרה יחד
- העדף מדריך עם פרופיל מחוץ למטרה נקי יותר על פני אחד עם פעילות גבוהה יותר רק במטרה
- שים לב לאורך המדריך, תוכן GC של 40–60%, וריצות הומופולימריות
-
הגבל חשיפה בתוך התא
- השתמש ב-RNP או mRNA במקום מערכות פלסמיד או ויראליות כאשר אפשרי
- השתמש ב-מינון האפקטיבי המינימלי
- הימנע מהארכת התמדה של העורך רק כדי לכפות תוצאות טרנספקציה
-
הוסף בקרות נוספות למקרים בסיכון גבוה יותר
- שקול ניקאזות מצומדות
- השתמש במערכות מושרות, ספליט-Cas9, או מבוקרות אור כאשר התזמון חשוב
- הוסף חלבוני אנטי-CRISPR כשלב כיבוי כאשר נדרש
-
אמת כראוי לאחר עריכה
- אשר את העריכה המדויקת תחילה
- בדוק כל אתר מחוץ למטרה שנחזה עם NGS אמפליקון ממוקד
- עבור ל-GUIDE-seq, CIRCLE-seq, CAST-seq, UDiTaS, או WGS כאשר הסיכון בפרויקט גבוה יותר
- עבור עורכי בסיס או עורכי פריים , הוסף בדיקות ברמת RNA כאשר רלוונטי
-
אל תשחרר שיבוט יחיד על סמך רצף בלבד
- השווה 2–3 שיבוטים עצמאיים
- השתמש בבקרה הורית לא ערוכה
- הסר שיבוטים עם חוסר יציבות או שינוי פנוטיפי
- שחרר רק כאשר מצב העריכה, סריקת מחוץ למטרה, והרשומות כולם שלמים
דרך קצרה לחשוב על זה: לעצב כדי להימנע מחיתוכים לא מכוונים, לספק כדי להגביל את הזמן בתא, ואז לאמת בעומק שהסיכון בפרויקט מצדיק. זהו החוט שעובר דרך כל היצירה.
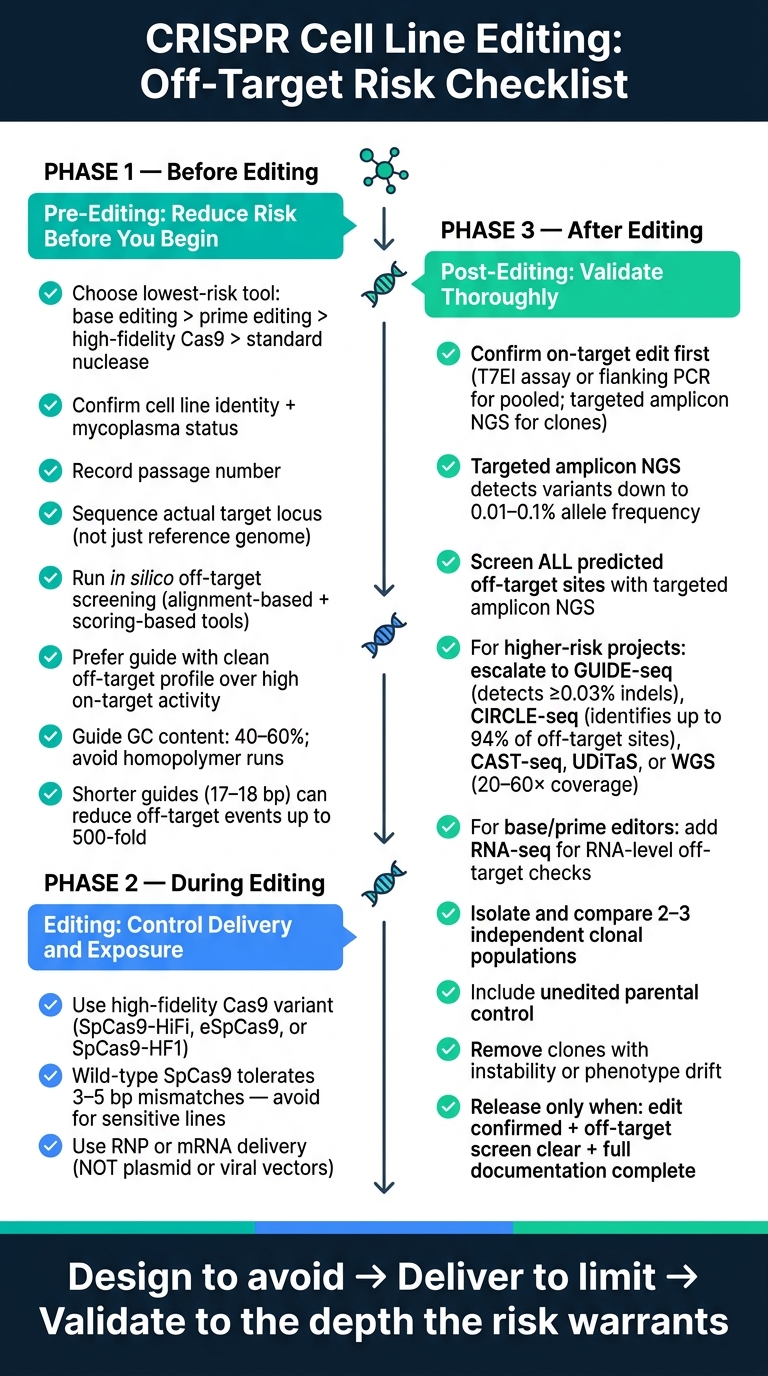
בקרת סיכון מחוץ למטרה של CRISPR: רשימת בדיקה בת 3 שלבים לעריכת קווי תאים
רשימת בדיקה לפני עריכה: הפחתת סיכון לפני תחילת העריכה
הגדר את מטרת העריכה ובחר את שיטת העריכה בעלת הסיכון הנמוך ביותר
לפני שאתה מזמין ריאגנט אחד, היה ברור מאוד מה העריכה אמורה לעשות. נוקאאוט, נוק-אין, שינוי נוקלאוטיד בודד ומודולציה טרנסקריפציונית אינם נושאים את אותו סיכון מחוץ למטרה. הם גם לא דורשים את אותו כלי.
סדר הסיכון הרחב הוא פשוט. נוקלאזות יוצרות DSB כמו Cas9 ו-Cas12 נמצאות בקצה העליון של הסיכון מכיוון שהן יכולות לגרום למחיקות גדולות, טרנסלוקציות ותגובות נזק ל-DNA [1] [7]. עורכי בסיס ועורכי פריים משתמשים בניקאזים, ולכן הם נמנעים מ-DSBs ומפחיתים את הסיכון לשינוי מבני [1][5]. למודולציה טרנסקריפציונית, עורכים אפיגנטיים כמו dCas9 שמחובר למודיפיקטורים טרנסקריפציוניים משאירים את רצף ה-DNA ללא שינוי [1].
הכלל המעשי הוא פשוט: השתמשו בשיטה הכי פחות גנוטוקסית שעדיין יכולה לספק את העריכה שאתם צריכים. לשינויים בנוקלאוטיד בודד, CBEs או ABEs מתאימים יותר מ-HDR, שעדיין יכול להכניס אינדלים [3] [1]. להחלפות והכנסות או מחיקות קטנות, עריכת פריים לעיתים קרובות מראה פעילות נמוכה יותר מחוץ למטרה מאשר CRISPR-Cas9 סטנדרטי [1]. אם עליך להשתמש בנוקלאז, בחר בגרסה בעלת דיוק גבוה כמו SpCas9-HiFi, eSpCas9, או SpCas9-HF1 [1] [6].
לאחר שהגישה לעריכה נקבעה, נעל את קו התאים העובד ואת רצף המטרה המדויק.
אשר את זהות קו התאים, ההיסטוריה ורצף אתר המטרה
אם קו התאים מזוהה באופן שגוי או מזוהם, שאר תהליך העבודה מתחיל להתערער. אפילו RNA מנחה מתוכנן היטב לא יציל חומר התחלה גרוע. בדוק את זהות קו התאים לפני תחילת כל עריכה. במקביל, אשר את מצב המיקופלסמה ורשום את מספר המעבר הנוכחי, שכן תאים במעבר גבוה יכולים לשנות את היציבות הגנומית ואת יעילות העריכה [1][6].
באותה מידה חשוב, אל תסתמך רק על גנום ייחוס.רצף את המיקום המדויק של היעד בקו התאים העובד. שלב זה עוזר לך לזהות SNPs או indels שעשויים לחסום את הקישור של המדריך או ליצור אתרים חדשים מחוץ ליעד [1] [6].
לאחר מכן, עבור לעיצוב המדריך.
בצע סינון מחוץ ליעד במחשב לפני בחירת חומרים
ברגע שמיקום היעד מאושר, סנן מדריכי RNA מועמדים במחשב לפני שאתה מתחייב לעבודה במעבדה רטובה. השתמש בכלים מבוססי יישור, כגון Cas-OFFinder או FlashFry, וכלים מבוססי ניקוד, כגון ניקוד CFD או DeepCRISPR . הקבוצה הראשונה עוזרת למצוא אתרים גנומיים עם הומולוגיה של רצף. השנייה עוזרת לדרג את האתרים הללו לפי הסתברות חיתוך חזויה [1][5].
כאשר מדריכים נבחרים לרשימה קצרה, פרופיל נקי מחוץ למטרה צריך לגבור על יעילות גולמית במטרה. מדריך עם 70% יעילות במטרה וללא תחזיות מחוץ למטרה הוא מקום בטוח יותר להתחיל ממנו מאשר אחד עם 90% יעילות וכמה אתרים בסיכון גבוה [6]. בכמה הגדרות, קיצור אורך המדריך מ-20 bp ל-17-18 bp יכול להפחית אירועים מחוץ למטרה עד פי 500 ללא אובדן משמעותי של דיוק במטרה [5]. שאפו לתוכן GC בין 40% ל-60%, והימנעו מרצפים של ארבעה או יותר בסיסים זהים [6][5].
עם זאת, לסינון במחשב יש מגבלות. הוא לא מתחשב היטב במצב הכרומטין, מחזור התא או הקשר ספציפי לתא [1][6][4]. חשבו על זה כמסנן, לא כהוכחה.זה מצמצם את השדה, אך לא מחליף אישור ניסיוני.
הבא את האתרים בעלי הסיכון הגבוה ביותר לתוכנית העריכה והאימות.
sbb-itb-ffee270
רשימת בדיקה לעריכה: בחירת עורך, משלוח וחשיפה
השתמש בעורכים בעלי ספציפיות גבוהה וב-RNA מדריך מדורג היטב
התחל עם רשימת הקצרה של אתרים מחוץ למטרה הצפויים והשתמש בה כדי לבחור את העורך. ברוב המקרים, גרסה בעלת דיוק גבוה של SpCas9 - SpCas9-HiFi, eSpCas9, או SpCas9-HF1 - היא ברירת מחדל טובה יותר מאשר SpCas9 מסוג פראי [6] [1]. SpCas9 מסוג פראי יכול לסבול עד שלושה עד חמישה אי-התאמות בזוגות בסיסים, במיוחד באזור PAM-distal, וזה יוצר סיכון משמעותי מחוץ למטרה בקווי תאים רגישים [3].
כלל פשוט עוזר כאן: השתמש ב-עורך בעל נאמנות גבוהה הכי פחות פעיל שעדיין מספק את העריכה המיועדת.
עבור עורכי בסיס, עקוב אחר עריכות לא מכוונות ו- השפעות לא מכוונות על RNA בנפרד מ-סיכון לא מכוון על DNA [1] [8] . אלה מצבי כשל שונים, והם זקוקים לבדיקות נפרדות. אם ניתן לבצע את העריכה ללא שבירות דו-גדיליות, עריכת בסיס או עריכה ראשונית עשויות להתאים יותר בתהליכי עבודה בסיכון גבוה יותר [1][8].
לאחר שנבחר העורך, המשימה הבאה היא לשמור על זמן השהייה שלו בתוך התא קצר ככל האפשר.
הגבל את התמדה של העורך עם אספקה זמנית ומינון אפקטיבי מינימלי
התמדה של העורך חשובה בדיוק כמו בחירת העורך.ככל שהעורך נשאר פעיל יותר בתא, יש לו יותר זמן לפעול באתרים בעלי סבירות נמוכה. זה הופך את פורמט המשלוח לנקודת בקרה מרכזית.
השתמש ב-משלוח חולף כמו RNPs או mRNA, והימנע מ- DNA פלסמידי או וקטורים ויראליים שמאריכים את ביטוי העורך [1] [5] . בפועל, משלוח RNP צריך להיות ברירת המחדל [6].
גם המינון חשוב. ריכוז גבוה של נוקלאז מגדיל את הסיכוי לחיתוך באתרים מחוץ למטרה בעלי רגישות נמוכה [5]. השתמש ב-מינון האפקטיבי המינימלי. אם יעילות הטרנספקציה נמוכה, אל תוסיף סתם עוד ריאגנט ותקווה לטוב ביותר. זה לעיתים קרובות משנה את הבעיה במקום לפתור אותה.
הוספת אמצעי הגנה מדויקים עבור תהליכים בסיכון גבוה יותר
חלק מהתהליכים דורשים אמצעי הגנה נוספים. זה נכון במיוחד עבור מטרות קרובות לאונקוגנים, מדכאי גידולים, או בקווי תאים רגישים ל-p53, שבהם אירוע מחוץ למטרה יכול לגרום לעלות גבוהה במיוחד [1][6][3].
אמצעי הגנה שימושיים כוללים:
- ניקאזות זוגיות, שדורשות שני חיתוכים קרובים. חיתוך יחיד מחוץ למטרה בדרך כלל מתוקן ללא מוטציה, כך שהסיכון מחוץ למטרה יורד משמעותית בהשוואה להגדרת נוקלאז סטנדרטית [4][1].
- מערכות Cas9 מפוצלות, ניתנות להשראה או נשלטות על ידי אור, שמסייעות לשמור על פעילות העורך בתוך חלון זמן צר כאשר המשלוח יעיל והחשיפה חייבת להישאר קצרה [1].
- חלבוני Anti-CRISPR (Acr), שפועלים כמתג כיבוי. חלבוני Acr טבעיים אלו יכולים לנטרל את קומפלקס CRISPR-Cas לאחר פרק זמן מוגדר, ומספקים לך בלם מולקולרי על פעילות העורך [1].
רשימת בדיקה לאחר עריכה: זיהוי אירועים מחוץ למטרה ואימות שיבוטים
בדוק אתרים מחוץ למטרה שנחזו עם ריצוף ממוקד
לאחר סיום העריכה, אשר את השינוי המיועד בלוקוס המטרה תחילה. לבדיקה מהירה ראשונית בתאים מאוגדים, ניתן להשתמש בבדיקת חיתוך אי-התאמה כמו T7 Endonuclease I, עיכול הגבלה, או PCR מקיף.רק היו זהירים עם הפרשנות: לכל אחת מהשיטות הללו יש מגבלות רגישות, במיוחד עבור עריכות נדירות או וריאנטים הומוזיגוטיים [9].
לאימות ברמת השיבוט, NGS אמפליקון ממוקד הוא הסטנדרט. הוא מספק מבט כמותי על תדירות האללים ויכול לזהות וריאנטים עד 0.01% עד 0.1% [3].
רצף כל אתר מחוץ למטרה שנחזה עם NGS אמפליקון ממוקד. זה צריך להיות שלב האימות ברירת המחדל.
הסלימו לבדיקות רחבות היקף או מבניות כאשר הסיכון בפרויקט גבוה יותר
סינון אתר-לפי-אתר אינו תמיד מספיק. אם העורך, אתר המטרה או קו התאים מציעים סיכון נסתר, עברו לבדיקה שיכולה לזהות אירועים שלא חזיתם מראש.
בדיקות גילוי רחבות היקף כמו GUIDE-seq ו-CIRCLE-seq אינן זקוקות לרשימות אתרים מחוץ למטרה מראש.GUIDE-seq יכול לזהות אתרים מחוץ למטרה עם תדירות אינדלים נמוכה עד 0.03% [2]. CIRCLE-seq יכול לזהות עד 94% מהאתרים מחוץ למטרה in vitro [3]. שיטות אלו שימושיות כאשר הקשר סוג התא עשוי להסתיר פעילות מחוץ למטרה.
אם אתה מודאג משינויים גדולים, קריאות אמפליקון סטנדרטיות עשויות להחמיץ את הבעיה העיקרית. מחיקות, היפוכים וטרנסלוקציות דורשות בדיקות שנבנו לשינויים מבניים, כגון CAST-seq ו-UDiTaS [1].
ריצוף גנום מלא (WGS) הוא האפשרות הרחבה ביותר. הוא יכול לזהות אינדלים, וריאציות מבניות ושינויים במספר העותקים ברחבי הגנום [1]. הפשרה היא עומק ועלות: בדרך כלל נדרש כיסוי של 20–60×, מה שהופך אותו להתאמה גרועה לסינון שגרתי של אוכלוסיות בתפזורת [1].
השתמש ב- NGS של אמפליקונים ממוקדים לאתרים חזויים. עבור לפרויקטים בסיכון גבוה יותר, השתמש בבדיקות גנום רחבות או מבניות. עבור עורכי בסיס או פריים, הוסף RNA-seq כדי לבדוק השפעות מחוץ למטרה ברמת ה-RNA.
בחר מספר שיבוטים עצמאיים ותעד קריטריוני שחרור
לאחר בדיקות הרצף, בדוק את הפנוטיפ ביותר משיבוט אחד.
אל תתקדם עם שיבוט ערוך יחיד. בידד והרחב לפחות שתי עד שלוש אוכלוסיות שיבוטיות עצמאיות והשווה אותן עם בקרת הורה לא ערוכה [4][9] . הסר שיבוטים שמראים חוסר יציבות או שינוי פנוטיפי [3]. לאחר מכן, אשר את העריכה המיועדת במצב האלל הנדרש, בין אם הטרוזיגוטי או הומוזיגוטי, באמצעות NGS של אמפליקון ממוקד [9].
תיעוד אינו עבודה מנהלית בסוף. זהו חלק משחרור השיבוט. רשום את רקע הקו ההורי, עיצוב ה-sgRNA, גרסת הנוקלאז, שיטת המסירה, וכל תוצאות ה-QC [3]. שיבוט צריך להתקדם רק כאשר העריכה המיועדת מאושרת, אתרי ה-off-target החזויים ברורים, וכל התיעוד במקומו.
עריכת גנום עם CRISPR: כיצד למזער ביעילות את ההשפעות הלא מכוונות
סיכום: רשימת תיוג בשלושה שלבים לעריכות קו תאים נקיות יותר
ביחד, רשימת התיוג מתייחסת לשליטה ב-off-target כתהליך מדורג, לא כבדיקת QC חד פעמית. המטרה פשוטה: לצמצם סיכון מוקדם, להגביל את פעילות העורך, ואז לאמת את התוצאה.
עומק האימות צריך להתאים לסיכון. שחררו רק שיבוטים עצמאיים מרובים שאושרו במצב האלל המיועד.
שאלות נפוצות
למה לא להסתמך על שיבוט אחד?
הסתמכות על שיבוט יחיד היא מסוכנת. עריכת CRISPR אינה ספציפית באופן מושלם, ולכן היא יכולה להכניס מוטציות לא מכוונות מחוץ למטרה.
זו הסיבה שצוותים בדרך כלל מרחיבים אוכלוסיות שיבוטיות מרובות. פעולה זו מקלה על מציאת קו שנושא את העריכה המיועדת במטרה ללא שינויים מזיקים מחוץ למטרה.
יש סיבה נוספת: קווי תאים יכולים להראות הטרוגניות גנטית. ריצוף שיבוטים מרובים עוזר לאשר שהשינוי המיועד נוקאאוט הומוזיגוטי או שינוי באתר המטרה אחר קיים בכל אתרי המטרה.
מתי NGS מבוסס אמפליקון מספיק?
ריצוף דור הבא מבוסס אמפליקון הוא לעיתים קרובות מספיק כאשר אתה זקוק לדרך ממוקדת וחסכונית לאשר אתרים פוטנציאליים מחוץ למטרה שסומנו על ידי כלים חישוביים או שיטות סינון אחרות.
ריצוף גנום מלא הוא עדיין הדרך היחידה לכמת באופן מלא את ההשפעות מחוץ למטרה. אבל עבור יישומים רבים, רמת הניתוח הזו פשוט לא נדרשת.
איך אני בוחר את העורך הבטוח ביותר?
בחר את הגרסה הפחות פעילה של נוקלאז CRISPR שעדיין חותכת את האתר שלך במדויק.
אי אפשר לבחור את הגרסה הטובה ביותר רק על סמך תחזיות. הדרך האמינה היחידה היא להריץ סקר קטן על פני גרסאות נוקלאז נבחרות ולקרוא את העריכה עם ריצוף דור הבא.
עבור בשר מתורבת R& D, זה נותן לך דרך מעשית קדימה: התחל עם רשימה קצרה של וריאנטים, ואז בדוק את החלשים יותר צעד אחר צעד עד שתמצא את האופציה הפחות פעילה שעדיין עורכת את האתר המטרה ביעילות.









