

리보솜 공학은 세포 수준에서 단백질 합성을 개선하여 배양육 생산을 혁신하고 있습니다. 리보솜은 세포의 단백질 공장으로, 육류의 질감과 영양 가치를 정의하는 액틴, 미오신 및 기타 단백질을 생산하는 데 중요합니다. 그러나 표준 세포주는 대규모 육류 배양에 필요한 높은 생산성을 위해 최적화되어 있지 않습니다.
주요 발전 사항은 다음과 같습니다:
- 최적화된 리보솜 RNA 변이체: 1.7 × 10⁷ 변이체를 포함한 스크리닝 라이브러리는 번역 활동 증가 가능성을 보여주었습니다.
- 직교 리보솜: 이러한 엔지니어링된 리보솜은 정상적인 세포 기능을 방해하지 않고 미오신과 같은 특정 단백질을 생산하는 데 특화되어 있습니다.
- 코돈 최적화: 리보솜의 선호도에 맞춰 mRNA 서열을 조정함으로써 최대 72배 높은 단백질 발현을 얻었습니다.
- 마이오카인 신호: IL-15 및 마이오넥틴과 같은 단백질은 근육 분화 동안 리보솜 생합성과 단백질 합성을 촉진합니다.
에너지 수요의 균형을 맞추고, 세포 안정성을 유지하며, 생산을 산업 수준으로 확장하는 데 여전히 도전 과제가 남아 있습니다. 예를 들어, 리보솜의 과도한 활동은 잘못 접힌 단백질이나 대사적 부담을 초래할 수 있으며, 생물 반응기에서의 영양소 확산 제한은 조직 성장을 200 μm 이상으로 제한합니다. 이러한 문제를 해결하려면 리보솜 공학을 고급 생물 처리 전략과 통합해야 합니다.
이 기사는 이러한 방법이 배양육의 미래를 어떻게 형성하고 있으며 상업적 실현 가능성을 달성하기 위해 극복해야 할 장애물에 대해 탐구합니다.
리보솜과 단백질 생합성: 기초
포유류 세포에서의 리보솜 구조와 기능
리보솜은 단백질 합성의 중심에 있으며, mRNA 서열을 기능적 단백질로 번역합니다.포유류 세포에서 리보솜은 80S 입자로 분류되며, 두 개의 소단위로 구성됩니다: mRNA를 해독하는 40S 소단위와 펩타이드 결합 형성을 촉진하는 60S 대단위입니다. 번역 과정은 세 가지 주요 단계로 이루어집니다: 개시, 시작 코돈이 인식되는 단계; 신장, 아미노산이 성장하는 폴리펩타이드 사슬에 순차적으로 추가되는 단계; 그리고 종결, 종결 코돈에 도달할 때 발생하는 단계.
대단위의 두 특정 영역은 엔지니어링 응용에 특히 중요합니다: 펩티딜 전이효소 중심 (PTC), 펩타이드 결합 형성을 촉진하는 영역과, 새로 합성된 폴리펩타이드가 빠져나가는 출구 터널, 입니다 [3].
이 핵심 메커니즘을 이해하는 것은 배양육 생산을 개선하기 위해 리보솜 성능을 최적화하는 방법을 탐구하는 데 필수적입니다.
배양육을 위한 단백질 생합성의 중요성
단백질 합성의 효율성은 특히 시험관 내 근육 형성 동안 배양육 개발에 있어 중요한 요소입니다. 이 과정은 근육 위성 세포(MSCs)를 액틴과 미오신 같은 수축성 단백질이 풍부한 다핵 근섬유로 변환합니다. 리보솜은 이 변환에서 중심적인 역할을 합니다 [4].
"전통적인 5,000 L 용량의 바이오리액터에서 1 kg의 단백질을 생산하기 위해 약 8조 개의 근육 세포가 필요합니다" [5]
이 엄청난 요구 사항은 리보솜 효율성의 작은 개선이 생산 수율을 크게 향상시킬 수 있으며, 이는 배양육의 상업적 실현 가능성에 직접적인 영향을 미친다는 것을 강조합니다.
세포가 성숙해짐에 따라, 그들의 리보솜 활동은 변화를 겪습니다.증식 단계 동안, MSCs는 빠른 분열을 우선시합니다. 그러나 분화가 시작된 지 3~5일 후에는 수축성 단백질의 성인형 동종체 합성과 세포가 근관으로 융합되는 것에 초점을 맞춥니다 [4]. 이 전환은 특정 신호 분자, 즉 마이오카인에 의해 조절됩니다.
예를 들어, 인터루킨‑15 (IL‑15)는 근육 발달 동안 주요 동화작용 인자로 작용하여 Myosin Heavy Chain (MyHC) 단백질의 축적을 촉진하고 단백질 분해를 감소시킵니다 [4]. 유사하게, 마이오넥틴은 PI3K/Akt/mTOR 신호 경로를 통해 단백질 합성을 강화하여 근육 성장을 지원합니다 [4]. 이러한 신호 경로가 리보솜 활동에 어떻게 영향을 미치는지를 이해하는 것은 생산 요구를 충족하는 확장 가능한 세포주를 설계하는 데 필수적입니다.이 통찰력은 후속 섹션에서 논의되는 엔지니어링 전략의 기초를 마련합니다.
리보솜 엔지니어링에 대한 현재 연구
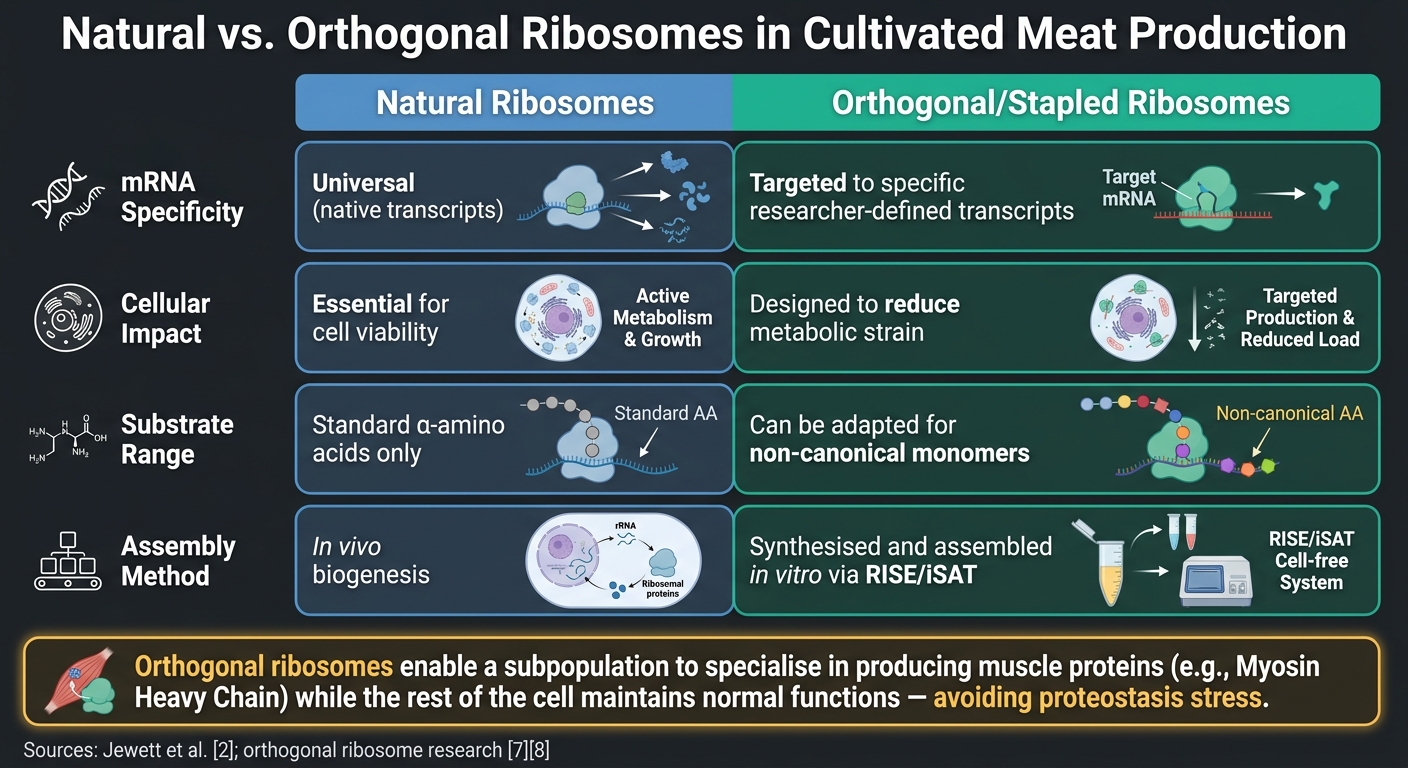
자연 리보솜 대 직교 리보솜: 배양육 생산에서의 역할
리보솜 생합성과 번역 조절
리보솜 생합성은 세포가 새로운 리보솜을 구성하는 과정으로, 매우 규제되고 에너지를 많이 소모하는 활동입니다. 포유류 세포에서는 세포 대사 산출의 큰 부분을 차지합니다. 번역만으로도 세포 전체 에너지 예산의 75% [8], 를 소비할 수 있어 가장 자원 소모가 많은 세포 과정 중 하나입니다.
리보솜 할당이 비효율적일 때 - 예를 들어, 리보솜이 초기 코딩 영역에서 멈출 때 - 자유 리보솜의 가용성을 감소시키는 병목 현상이 발생하여 궁극적으로 단백질 생산을 제한합니다.계산 모델에 따르면, 단 100개의 유전자를 조작하여 이러한 병목 현상을 해결하면 효모(Saccharomyces cerevisiae)에서 리보솜 할당이 35% 개선되고 대장균( Escherichia coli)에서 57% 개선될 수 있습니다. [8]. 이러한 발견은 특히 에너지 효율성과 단백질 출력이 중요한 배양육 산업에서 포유류 세포의 리보솜 역학 최적화에 직접적인 영향을 미칩니다.
배양육 맥락에서의 리보솜 공학
리보솜 공학의 발전은 이제 리보솜 생합성에 대한 기초 지식을 바탕으로 배양육 생산에 적용되고 있습니다. 근육 세포에서 직접 수행되지 않은 연구조차도 배양육 세포주에 관련된 통찰력을 제공하고 있습니다.
2020년 12월, Hadas Zur와 Tamir Tuller는 텔아비브 대학교에서 리보솜 트래픽 엔지니어링 (RTE) 의 잠재력을 통해 성장률과 단백질 출력을 향상시킬 수 있음을 입증했습니다. CRISPR-Cas9 , 을 사용하여 RPO21과 CYS4 의 램프 영역(코돈 11–50)에 동의어 돌연변이를 도입했습니다. S. cerevisiae . 결과적으로 생성된 이중 돌연변이는 로그 단계 성장과 세포 밀도가 개선되었습니다. 그러나 연구자들은 번역 최적화와 성장률 간의 관계가 이원적 전환 및 정지 단계 동안 번역 이외의 요인이 속도 제한 요소가 되는 경우 감소한다고 경고했습니다 [8]. 이 통찰력은 배양육 생산에서 분화 프로토콜을 설계하는 데 특히 중요합니다.
2020년 2월, 노스웨스턴 대학교의 Michael Jewett 팀은 RISE (Ribosome In vitro Synthesis and Evolution) 방법을 검증했습니다. 이 기술은 약 1.7 × 10⁷ 리보솜 RNA 변이 [2] . 의 라이브러리를 스크리닝하는 것을 포함합니다. RISE는 살아있는 세포 외부에서 완전히 작동하여, 생체 내에서 연구할 수 없는 치명적인 리보솜 돌연변이에 의해 부과되는 제약을 우회합니다.
"시험관 내 접근법은 세포 생존성 제약을 극복하여 치명적인 리보솜 돌연변이를 탐구할 수 있게 합니다." - Michael Jewett et al. [2]
배양육을 위한 또 다른 유망한 혁신은 직교 리보솜. 의 사용입니다. 이 엔지니어링된 리보솜-mRNA 쌍은 세포의 고유 번역 기계와 독립적으로 작동합니다.이는 연구자들이 필수적인 세포 과정에 간섭하지 않고 근육 질감에 중요한 Myosin Heavy Chain (MyHC) 이소폼과 같은 특정 표적에 리보솜 활동을 집중할 수 있게 합니다 [6]. 비교 연구는 자연 리보솜에 비해 직교 리보솜의 장점을 강조합니다:
| 특징 | 자연 리보솜 | 직교/스테이플 리보솜 |
|---|---|---|
| mRNA 특이성 | 보편적 (자연 전사체) | 특정 연구자가 정의한 전사체에 맞춤화됨 [6] |
| 세포 영향 | 생존에 필수적 | 대사 부담을 줄이도록 설계됨 [7] |
| 기질 범위 | 표준 α-아미노산 | 비정형 모노머에 맞게 조정 가능 [7] |
| 조립 | 생체 내 생합성 | RISE/iSAT [2]를 통해 시험관 내에서 합성 및 조립됨 |
여기서 중요한 점은 직교 리보솜이 리보솜의 하위 집단이 MyHC와 같은 근육 단백질을 생산하는 데 특화될 수 있도록 하면서 나머지 세포는 정상적인 기능을 유지할 수 있다는 것입니다.이는 전체 번역 시스템이 특정 단백질을 과잉 생산하도록 압박받을 때 발생할 수 있는 단백질 항상성 스트레스의 위험을 피합니다.
리보솜 성능 향상을 위한 전략
리보솜 생합성 증가
리보솜 수를 늘리는 것은 단백질 생산을 향상시키는 직접적인 방법이며, 두 가지 주요 방법이 주목받고 있습니다. 첫 번째 방법은 리보솜 RNA (rRNA) 유전자의 후성유전학적 상태를 수정하여 번역 능력을 증가시키는 것입니다.
"리보솜 RNA 유전자의 후성유전학적 공학은 단백질 생산을 향상시킵니다." - Santoro R., Lienemann P., Fussenegger M. [1]
두 번째 접근법은 PI3K/Akt/mTOR 신호 경로를 활용합니다. IL-15, 마이오넥틴, 아이리신과 같은 마이오카인은 이 경로를 활성화하여 근관 성숙 동안 리보솜 생합성을 촉진합니다.
그러나 리보솜 생산의 증가는 세포의 대사 능력과 신중하게 균형을 맞춰야 하며, 리보솜 합성은 살아있는 세포에서 가장 에너지를 많이 소모하는 과정 중 하나입니다 [1].
리보솜 수가 증가하면, 이들이 번역에 완전히 참여하도록 보장하는 데 초점이 맞춰집니다.
번역 개시 및 신장 개선
모든 리보솜의 활동을 최대화하는 것이 필수적이며, 성장 최적화된 세포에서도 15–20%의 리보솜이 비활성 상태로 남아 있습니다 [9]. 이는 배양육 세포주에서 활용되지 않은 상당한 예비 용량을 나타냅니다.
번역 신장의 속도는 두 가지 요인에 따라 달라집니다: 리보솜의 고유한 속도와 번역에 적극적으로 참여하는 리보솜의 비율 [9]. 이를 최적화하기 위해 배양 배지에서 높은 아미노산 수준을 유지하는 것이 중요합니다.또한, 리보솜 단백질을 안정화하기 위해 세포주를 엔지니어링하는 것은 rRNA의 오접힘과 분해로부터 보호하여, 성장 조건이 최고조에 달할 때 rRNA의 일반적인 10% 손실을 줄이는 데 도움이 됩니다 [9].
리보솜 활동이 최대화되면, 단백질 합성을 더욱 가속화하기 위해 mRNA 서열을 정제하는 것이 다음 단계가 됩니다.
mRNA 최적화 및 코돈 사용
리보솜의 성능은 그들이 처리하는 mRNA의 품질에 크게 의존합니다. 코돈 최적화는 목표 단백질의 코딩 서열을 숙주 종에 특정한 tRNA 풀과 일치하도록 조정합니다 - 예를 들어, 소, 돼지, 또는 어류와 같은. 이러한 정렬은 신장 중 리보솜의 정지를 방지하고 MyoD 및 Myf5와 같은 중요한 근원성 단백질의 처리량을 증가시킵니다.
코돈 최적화 외에도, 전사 조정은 세포 내 rRNA와 mRNA 수준 간의 적절한 균형을 보장합니다.이러한 구성 요소 간의 불일치는 병목 현상을 초래하여 전체 효율성을 감소시킬 수 있습니다 [1].
실용적인 응용을 위해, 통합 합성, 조립 및 번역 (iSAT) 시스템은 귀중한 도구를 제공합니다. 이러한 시스템은 세포 추출물과 형광 기반 분석을 사용하여 최적화된 mRNA를 시험관 내에서 프로토타입으로 만든 후 안정적인 세포주에 통합합니다. 이 반복적인 접근 방식은 연구자들이 코돈 최적화된 변형체를 빠르게 비교할 수 있게 하여 필수적인 근육 형성 단백질의 수율을 개선하고 배양육 생산의 확장성을 강화합니다 [1].
절충: 성장, 분화 및 제품 품질
리보솜 성능을 최적화하는 것은 단백질 합성을 증대시키는 것과 세포 성장 및 분화에 미치는 영향을 관리하는 것 사이의 미세한 균형을 포함합니다.
대사 부담과 단백질 항상성 스트레스
단백질 생산을 증대시키기 위해 리보솜을 조작하는 것은 ATP와 아미노산을 다른 중요한 세포 기능에서 빼앗아 에너지 수요를 증가시킵니다. 리보솜 합성은 이미 세포 내에서 가장 에너지를 많이 소모하는 과정 중 하나이며, 추가적인 증폭은 이러한 에너지 문제를 악화시킬 수 있습니다.
이러한 강화된 활동은 단백질 품질에도 영향을 미칠 수 있습니다. 과활성화된 리보솜은 세포 내 샤페론을 압도하여 잘못 접힌 단백질을 생성하고 비접힌 단백질 반응(UPR)을 활성화시킬 수 있습니다. 이러한 스트레스는 성장을 억제하거나 심지어 세포 사멸을 초래할 수 있습니다. 소나 양과 같은 가축 종의 성체 줄기세포는 자연적으로 증식 능력이 제한되어 있어, 이러한 추가적인 스트레스는 노화가 시작되기 전에 유효한 세포 분열 횟수를 크게 줄일 수 있습니다 [5].
배양육 생산에서 조직 두께는 영양소 확산의 제약으로 인해 200 μm를 거의 초과하지 않으며, 이는 더 큰 조직 집합체의 중심에서 세포 사멸을 초래할 수 있습니다 [5]. 에너지 소비를 증가시키는 전략은 일관된 단백질 합성이 필수적인 이러한 중요한 영역에서 영양소 고갈을 가속화할 위험이 있습니다. 또한, 대사적 부담이 증가하면 근육 분화에 필요한 정밀한 신호 경로에 방해가 될 수 있습니다.
근육 분화 및 단백질 구성에 미치는 영향
리보솜 공학에 의해 도입된 스트레스는 대사를 넘어 근육 발달을 방해할 수 있습니다.근형성, 근육 형성 과정은 전사 인자의 엄격하게 조절된 순서에 의존합니다: Pax7은 줄기세포가 정지 상태를 유지하도록 하고, Myf5는 근모세포의 증식을 촉진하며, MyoD 는 분화를 촉발합니다 [5] . 단백질 합성의 변화는 이 순서를 방해하여 분화를 지연시키거나 비정상적인 근섬유 구성을 초래할 수 있습니다. 이는 배양육에서 바람직한 질감과 풍미를 얻기 위한 핵심 요소인 근내 지방 침착이 감소할 수 있습니다 [5].
따라서 공학 과정 전반에 걸쳐 근형성 마커의 발현을 모니터링하여 엄격한 품질 관리를 유지하는 것이 적절한 근육 발달과 제품 품질을 보장하는 데 필수적입니다.
sbb-itb-ffee270
연구 격차 및 미래 방향
리보솜 공학의 발전은 유망하지만, 상업적인 배양육 생산에의 적용에는 여전히 상당한 장애물이 있습니다. 이러한 격차를 해소하기 위해 연구자들은 장기 생산의 요구를 견딜 수 있는 고급 분자 프로파일링 기술과 확장 가능한 생물 공정 전략에 집중해야 합니다.
멀티 오믹스 및 장기 안정성 연구
주요 도전 과제는 엔지니어링된 세포주에 대한 장기 안정성 데이터의 부족에 있습니다. 시간이 지남에 따라 이러한 세포는 자발적인 돌연변이를 축적하여 잠재적으로 그들의 표현형을 변경할 수 있습니다. 노비사드 대학교의 이바나 파친은 이 문제를 강조합니다: 불멸화된 세포는 "장기 배양 중 잠재적인 자발적 돌연변이로 인해 항상 1차 배양을 대표하지는 않습니다" [13]. 리보솜 엔지니어링 라인의 경우, 리보솜 구성 요소의 돌연변이는 즉각적인 감지 없이 번역 효율성을 저해할 수 있어 위험이 더욱 큽니다.
멀티오믹스 접근법은 이러한 문제를 해결할 수 있는 방법을 제공합니다. 전사체학, 단백질체학, 대사체학을 통합함으로써 연구자들은 Pax7, MyoD, 및 Myogenin, 과 같은 중요한 근원성 마커와 MyHC 아이소폼의 변화를 모니터링할 수 있습니다. 그런 다음, 게놈 규모의 대사 모델은 이러한 통찰력을 매체 구성의 실행 가능한 변화로 번역하여 엔지니어링된 리보솜의 고유한 요구를 충족시킬 수 있습니다 [5][11]. 배양육의 경우, 장기간의 주기 동안 일관된 단백질 생산을 보장하는 것이 필수적입니다. 이러한 장기 모니터링 없이는 지속 가능한 개선과 단기적인 효과를 구분하기 어렵습니다.
유전적 및 대사적 안정성 외에도 이러한 혁신을 산업 수준으로 확장하는 것은 자체적인 과제를 제시합니다.
생물공정 통합 및 규모 확장
리보솜이 조작된 세포를 작은 플라스크에서 산업용 생물반응기로 확장하는 것은 쉬운 일이 아닙니다. 5,000 L 교반 탱크 생물반응기에서 단 1kg의 단백질을 생산하려면 약 8조 개의 근육 세포가 필요합니다 [5]. 이러한 밀도에서는 영양소 구배가 중요한 문제가 됩니다. 산소 및 기타 영양소의 200 μm 확산 한계는 3D 조직 구조의 중심에 있는 세포가 특히 단백질 합성이 활발할 때 자원 수요가 최고조에 달할 때 기아에 직면할 수 있음을 의미합니다.
생물반응기 교반으로 인한 전단 응력은 또 다른 복잡성을 추가합니다. 변형되지 않은 세포는 이 난류를 견딜 수 있지만 번역 기계가 수정된 조작된 세포는 더 취약할 수 있습니다.스트레스는 세포 경로를 방해할 뿐만 아니라 이미 대사적 부담을 받고 있는 세포에 물리적 손상을 줄 수 있습니다 [13]. 이 문제를 해결하려면 실시간 데이터를 디지털 생물 제조 모델과 통합하여 대규모 용기 내 다양한 미세 환경을 더 잘 이해하고 예측하기 위해 전산 유체 역학 시뮬레이션을 포함해야 합니다 [10]. 수확과 같은 다운스트림 프로세스도 주의가 필요합니다 - 트립신을 포함한 효소적 방법은 엔지니어링된 세포의 표면 프로테옴을 변경할 수 있습니다 [14], 리보솜 엔지니어링의 이점을 잠재적으로 무효화할 수 있습니다.
| 확대 요인 | 주요 병목 현상 | 리보솜 공학과의 관련성 |
|---|---|---|
| 영양소 확산 | 200 μm 침투 한계 [5] | 3D 조직에서 단백질 합성 요구가 높은 세포를 굶주리게 할 수 있음 |
| 유전적 안정성 | 자발적 돌연변이 [13] | 시간이 지남에 따라 공학적으로 설계된 번역 효율성을 저해할 수 있음 |
| 전단 응력 | 교반 탱크 난류 [13] | 공학적으로 설계된 세포 경로를 방해할 위험이 있음 |
| 수확 방법 | 트립신에 의한 단백질 분해 손상 [14] | 단백질 품질의 개선을 가리고 단백질체를 변화시킬 수 있음 |
이러한 확장 문제를 해결하는 것은 리보솜 공학을 실험실에서 상업 생산으로 전환하는 데 필수적입니다.각 전략은 산업 조건에서 신뢰할 수 있는 단백질 수율, 안정성 및 안전성을 보장하기 위해 철저히 테스트되어야 합니다.
결론: 배양육에서 리보솜 공학의 필요성
5,000 L 바이오리액터에서 1 kg의 단백질을 생산하려면 무려 8조 개의 근육 세포가 필요합니다 [5]. 이는 배양육 생산을 확장하는 데 있어 엄청난 도전을 강조합니다. 리보솜 공학은 단순히 세포 수를 늘리는 대신 개별 세포의 단백질 출력을 개선함으로써 해결책을 제공합니다.
리보솜 공학을 적용할 때 타이밍이 중요합니다. 잘못된 단계에서 번역을 강화하면 근형성에 방해가 되어 MyHC와 같은 주요 수축 단백질의 생산에 영향을 미칠 수 있습니다 [5]. 번역과 근형성 사이의 적절한 균형을 이루는 것은 공학 자체만큼이나 중요합니다.
"고품질 CBM과 높은 수율의 생산을 달성하기 위해서는 상업적 생산을 위한 우수한 실험실 관행을 달성하기 위해 분자적 측면에 대한 철저한 검토가 필요합니다." - Asim Azhar et al., Frontiers in Food Science and Technology [5]
번역 개시 인자(eIF3i 및 eIF3c)의 과발현, 코돈 최적화, mRNA 수정 타겟팅과 같은 여러 기술이 이미 재조합 단백질 출력 증가에 유망한 결과를 보여주었습니다 [15]. 그러나 이러한 방법은 대사 부담, 단백질 항상성 스트레스, 장기적인 유전적 불안정성과 같은 문제를 피하기 위해 주의 깊게 적용해야 합니다. 분자 최적화는 필수적이지만, 영양소 확산 한계, 전단 응력 민감성, 수확 중 단백질체 붕괴와 같은 문제를 완전히 해결할 수는 없습니다.이러한 장애물은 생물공정 설계의 동시 발전을 필요로 합니다.
배양육의 잠재적인 환경적 이점은 막대합니다. 이는 전통적인 가축 사육에 비해 온실가스 배출을 78%–96% 줄이고, 토지 사용을 99% 줄이며, 물 사용을 82%–96% 줄일 수 있습니다 [12]. 이러한 이점을 대규모로 달성하려면 현재의 세포 배양 생산성과 경제적 타당성 간의 격차를 해소해야 합니다. 리보솜 공학은 이 격차를 해소하는 데 강력한 도구가 될 수 있지만, 분자 생물학, 생물공정 혁신 및 포괄적인 다중 오믹스 모니터링을 포함한 보다 넓고 통합된 접근 방식의 일부가 되어야 합니다. 이러한 노력을 결합해야만 배양육의 모든 가능성을 실현할 수 있습니다.
어떻게 Cellbase 배양육 연구를 지원하는지
분자 최적화에서 대규모 생산으로 나아가는 배양육은 모든 단계에서 정밀한 도구와 재료가 필요합니다.
세포주 최적화 작업을 하는 팀에게
생산을 확대할 때,
자주 묻는 질문
배양육 세포주에 가장 유망한 리보솜 공학 접근법은 무엇인가요?
배양육을 위한 리보솜 공학 연구는 단백질 생합성을 향상시키고 세포 운명 결정을 영향을 미치는 것을 목표로 합니다. 유망한 접근법 중 하나는 리보솜 풀 공학, 으로, 번역 효율성을 개선하기 위해 리보솜 RNA 오페론을 수정하는 것입니다. iSAT 및 RISE와 같은 도구는 시험관 내 리보솜 진화를 위한 플랫폼을 제공하여 기능이 향상된 리보솜 개발을 가능하게 합니다. 또한,
번역 속도를 높이면서 잘못 접힌 단백질이나 세포 스트레스를 유발하지 않으려면 어떻게 해야 합니까?
번역 속도를 전반적으로 가속화하는 대신 번역 과정을 미세 조정하는 데 중점을 둠으로써 단백질 오접힘이나 세포 스트레스를 유발하지 않고 번역 속도를 개선할 수 있습니다. 주요 접근 방식은 다음과 같습니다:
- 느리게 번역되는 코돈 사용 : 이들은 번역 속도를 단백질 접힘의 자연스러운 과정과 일치시켜 적절한 구조 형성을 보장합니다.
- 5' 코딩 영역에서 자유 접힘 에너지 감소: 이 조정은 세포 건강을 유지하면서 단백질 생산 효율성을 향상시킬 수 있습니다.
다른 기술로는 저유도 체제, 온도 하향 이동, 및 SINEUP RNA. 와 같은 고급 합성 도구가 포함됩니다. 이러한 전략은 세포에 과부하를 주지 않고 더 높은 단백질 수율을 가능하게 합니다.
특수 재료를 다루는 분들을 위해,
200 µm 이상의 리보솜-엔지니어드 근육 조직을 지원하기 위해 생물반응기에 어떤 변화가 필요합니까?
200 µm보다 두꺼운 근육 조직을 성장시키기 위해서는 생물반응기가 영양소, 산소, pH 확산과 관련된 문제를 극복해야 합니다. 이는 3차원 구조에서 세포 생존에 중요한 요소입니다. 교반 탱크 생물반응기는 균일한 조건을 유지하면서 세포에 해를 끼칠 수 있는 전단 응력을 줄이기 위해 정밀한 조정이 필요합니다. 많은 경우, 관류 기반 시스템은 특히 밀집된 조직에서 안정적인 환경을 조성하는 데 중요한 역할을 합니다. 특수 생물반응기와 재료를 다루는 분들을 위해,









