

Si puedes expandir células pero no puedes cambiarlas al destino correcto en el momento adecuado, tu proceso se detendrá en la diferenciación. Ese es el punto central aquí: los circuitos genéticos sintéticos te dan dentro de la célula control sobre el compromiso, el tiempo, la memoria y la mezcla de linajes, donde los cambios de medios por sí solos a menudo dejan poblaciones heterogéneas, parcialmente comprometidas.
Si estuviera construyendo un flujo de trabajo de diferenciación de carne cultivada, tomaría cuatro puntos de este artículo de inmediato:
- Comienza con la red nativa, no con la construcción. Usa análisis de trayectoria snRNA-seq, , inferencia de GRN y perfilado de miRNA para encontrar dónde las células se detienen, desvían o ramifican hacia el destino incorrecto.
-
Empareja el tipo de circuito con el problema del proceso.
Un interruptor de palanca es adecuado para el bloqueo, un diseño de retroalimentación o paso de banda es adecuado para el control de tiempo, una puerta lógica es adecuada para el control de múltiples señales, y miSFITs son adecuados para salida graduada. - Diseñar para baja fuga, bajo ruido y seguridad desde el primer día. Las partes ortogonales, la autorregulación negativa, los iFFLs, los transgenes cm y un módulo de muerte inducible o de detención del crecimiento son parte de la construcción, no una ocurrencia tardía.
- Validar bajo condiciones relevantes a escala temprano. Un circuito que funciona en 2D puede cambiar en 3D, microportador o suspensión agitada debido a gradientes de inductor, límites de oxígeno y cizallamiento.
El artículo también hace un punto práctico que importa para los equipos de proceso: el control de una sola línea y el control de proporción son trabajos diferentes. Un casete Tet-On MyoD puede impulsar la entrada miogénica, pero los productos de corte completo necesitan control de proporciones de músculo, grasa y ECM, lo que generalmente significa retroalimentación, señalización paracrina y una selección más rigurosa de clones.
Algunas cifras anclan ese mensaje:
- La diferenciación miogénica estándar puede detenerse con índices de fusión de aproximadamente 50–60%
- Los GRNs diseñados en iPSCs aumentaron la diferenciación de la línea objetivo de 52% a 81%
- Los circuitos sintéticos en MSCs modificados impulsaron la diferenciación cardíaca a 76%
- Algunas líneas porcina Tet-On-PAX7 mantuvieron un alto potencial miogénico más allá de 40 pasajes
- Alrededor del 20% de las células madre pluripotentes humanas pueden portar mutaciones vinculadas al cáncer, por lo que los módulos de seguridad inducibles son importantes
Flujo de Trabajo de Circuitos Genéticos Sintéticos para la Diferenciación de Carne Cultivada
Historias de Científicos: Michael Elowitz, Circuitos Genéticos en Células Vivas
Comparación rápida
| Tema | Lo que dice el artículo en términos sencillos |
|---|---|
| Cuello de botella principal | Diferenciación, no expansión |
| Por qué los circuitos ayudan | Agregan control de umbral y memoria de estado celular dentro de la célula |
| Mejor primer paso | Mapear el control del destino nativo con snRNA-seq y herramientas relacionadas |
| Opciones principales de circuitos | Conmutador, avance, paso de banda, puertas lógicas, miSFITs, CRISPRa/i |
| Ejemplo de miogénesis | Tet-On-MyoD para separar el crecimiento de la diferenciación terminal |
| Adipogénesis / control de ECM | miSFITs y diseños de corbata de lazo para salida graduada tipo PPARG/BMP4 |
| Desafío de corte completo | Control de proporción entre músculo, grasa y tejido conectivo |
| Riesgo de escalado | El comportamiento en 2D puede no mantenerse en 3D o en biorreactores |
| Opciones de integración | Lentivirus, transposones, CRISPR knock-in, vectores episomales |
| Punto regulatorio | Las líneas diseñadas necesitan un paquete de seguridad más amplio; se prefieren inductores seguros para alimentos como ácido vanílico sobre DOX cuando sea posible |
Entonces, en términos simples: esto no es solo un artículo de diseño de circuitos. Lo leería como una guía para vincular la arquitectura de circuitos, la biología de linaje, la selección de clones, el rendimiento del biorreactor y la documentación de seguridad del Reino Unido/UE en una estrategia de diferenciación.
Continúa leyendo si deseas conocer el camino completo desde el mapeo de la red nativa hasta la elección de construcción, las verificaciones de escalado y el ajuste regulatorio.
2. Principios de diseño para circuitos de control de diferenciación
2.1 Mapea la red de destino celular nativa antes de diseñar el circuito
Antes de diseñar un circuito, necesitas una imagen clara de lo que la célula ya está haciendo.
La secuenciación de ARN de núcleo único (snRNA-seq) es un buen punto de partida. Puede mostrar subpoblaciones quiescentes, incluidas las células de reserva marcadas por NOTCH2 y HEYL , y señalar objetivos de vías que pueden mejorar la diferenciación [3].
A partir de ahí, el análisis de trayectoria y la inferencia de la red reguladora de genes (GRN) ayudan a mapear el orden de activación de los reguladores y destacan dónde es más probable que las células se detengan. En la miogénesis, la cascada principal pasa por MYOD1 y MYOG . En la adipogénesis, los nodos principales son PPARG y CEBPA , con el destino de los progenitores fibro-adipogénicos (FAP) actuando como el principal punto de riesgo de ramificación. La tabla a continuación resume los principales reguladores y cuellos de botella.
| Línea genética | Reguladores maestros clave | Vías de señalización críticas | Cuellos de botella identificados |
|---|---|---|---|
| Miogénico | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Formación de células de reserva (quiescencia) |
| Adipogénico | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Destino de progenitores fibro-adipogénicos (FAP) |
| Pluripotente | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Diferenciación espontánea / heterogeneidad |
Otra capa útil es el perfil de expresión de miRNA. Los miRNAs endógenos como miR-302a, vinculados a la pluripotencia, y miR-375, vinculados a la diferenciación, pueden actuar como clasificadores internos en diseños de sentido y respuesta. Eso permite que el circuito lea el estado real de la célula en lugar de depender solo de inductores externos [5].
Esos cuellos de botella deberían guiar la elección del circuito. Si el problema principal es la deriva, puede necesitar un bloqueo. Si el problema es el tiempo, un pulso puede ser más adecuado. Si el control del destino depende de más de una señal, la lógica de entrada múltiple generalmente tiene más sentido.
2.2 Elija la arquitectura de circuito adecuada
Aquí es donde los compromisos se muestran a plena vista. La arquitectura adecuada depende de tres puntos prácticos: cuán permanente debe ser el compromiso, cuán estrictamente se necesita controlar el tiempo y cuánta carga genética puede llevar la célula sin problemas.
Interruptores de palanca biestables se ajustan a los casos donde el compromiso de linaje necesita permanecer bloqueado. El problema principal es el cambio espontáneo de estado causado por el ruido transcripcional.
Los filtros de paso de banda se adaptan a los casos en los que se necesita un factor de transcripción solo dentro de una ventana de desarrollo definida. El inconveniente es que los niveles de inductor necesitan un control estricto, o el tiempo se desliza.
Las puertas lógicas añaden especificidad al requerir más de una entrada al mismo tiempo. Por ejemplo, la diferenciación solo puede permitirse cuando hay un inductor exógeno presente y la célula muestra el perfil correcto de miARN endógeno. Eso ayuda a reducir el riesgo de compromiso fuera del objetivo.
La tabla a continuación describe las principales arquitecturas y los compromisos que conllevan.
| Arquitectura | Reversibilidad | Precisión temporal | Complejidad de integración | Uso principal | Riesgo clave |
|---|---|---|---|---|---|
| Interruptor biestable | Bajo (una vez bloqueado) | Moderado | Moderado | Compromiso de linaje permanente | Volteo espontáneo debido al ruido |
| Filtro de paso de banda | Alto (dependiente de la concentración) | Alto | Alto | Etapas de desarrollo transitorias | Requiere control estricto del inductor |
| Puerta lógica (AND/OR/NOT) | Variable | Moderado | Moderado–alto | Activación específica de tipo celular | Fugas en estados OFF |
| Corbata de lazo / entrada múltiple | Alto | Moderado | Moderado | Integración de múltiples señales | Depende de la estabilidad endógena de miRNA |
| miSFITs | Alto | Moderado | Bajo–moderado | Ajuste de salida gradual | Rango dinámico estrecho si está mal ajustado |
"Al minimizar el número de capas computacionales mientras se mantiene la funcionalidad, esta estrategia aborda las barreras de escalabilidad en la ingeniería de circuitos genéticos." - Nature Communications [9]
Cada capa reguladora añadida aumenta la carga útil y consume recursos celulares. En la práctica, si dos diseños realizan la misma función, el más simple suele ser la mejor opción cuando la escala importa.
Una vez establecida la arquitectura, el siguiente trabajo es hacer que se mantenga bajo baja filtración, supresión de ruido y control a prueba de fallos.
2.3 Construir para fiabilidad, baja filtración y seguridad
Un circuito debe permanecer estable durante un cultivo prolongado. Un breve período de buen rendimiento no es suficiente para su uso en producción.
Las partes ortogonales son la primera defensa. Promotores, factores de transcripción y elementos reguladores que no reaccionan de forma cruzada con la maquinaria nativa ayudan a limitar los efectos fuera de objetivo y reducen la posibilidad de que las señales endógenas activen o desactiven el circuito.Promotores modificados de alta hermeticidad como PCREm se han utilizado para reducir la fuga basal en sistemas mamíferos inducibles [6].
La autorregulación negativa también es recomendable añadirla cuando sea posible. Es uno de los motivos más conocidos para reducir el ruido transcripcional y producir una respuesta más lineal a la concentración del inductor [6]. Los bucles de retroalimentación incoherente (iFFLs) pueden añadir otra capa de control al filtrar las fluctuaciones estocásticas, de modo que las células respondan a señales sostenidas en lugar de picos de ruido cortos.
Las versiones modificadas por codón (cm ) de factores de transcripción sintéticos también facilitan la caracterización. Permiten separar la expresión impulsada por circuitos de la expresión genómica endógena (g) durante la validación [1]. Eso puede parecer un detalle pequeño, pero ahorra tiempo cuando estás tratando de determinar si una lectura proviene del circuito o del genoma del huésped.
Se requieren módulos de seguridad. Alrededor del 20% de las células madre pluripotentes humanas llevan mutaciones asociadas al cáncer [7]. Por lo tanto, si un circuito va a una línea derivada de células madre, debe incluir un módulo de detención de crecimiento o eliminación inducible. El ácido vanílico es un inductor útil a priorizar aquí porque es un aditivo alimentario autorizado, lo que ayuda a justificar su uso como desencadenante de circuitos en líneas celulares de carne cultivada [1].
"La biología sintética proporciona a los ingenieros los medios para desplegar circuitos que ajusten fácil y precisamente la expresión de múltiples genes para... eliminar posibles efectos secundarios adversos fuera del objetivo." - npj Systems Biology and Applications [6]
Estas elecciones configuran los circuitos específicos de linaje en la Sección 3.
sbb-itb-ffee270
3. Estrategias de circuitos para la diferenciación miogénica, adipogénica y controlada por proporción
3.1 Circuitos miogénicos que separan la expansión de la diferenciación terminal
Una vez que se establece la arquitectura del circuito, el siguiente trabajo es el despliegue específico de linaje. Para la miogénesis, el problema principal es simple de enunciar pero difícil de ejecutar: las células necesitan proliferar primero, luego cambiar a diferenciación cuando se les pida, sin desviarse demasiado pronto en ninguna dirección.
Un casete Tet-On-MyoD es una de las formas más directas de hacer esto. En esta configuración, las células proliferan bajo condiciones estándar cuando la doxiciclina (DOX) está ausente. Agregue DOX, y el circuito impulsa el compromiso miogénico.Investigadores en múltiples instituciones chinas utilizaron este enfoque en fibroblastos embrionarios de pollo y reportaron una formación eficiente de miotubos después de la inducción [4].
Un interruptor de palanca biestable ofrece un control de estado más estricto. Los sistemas construidos a partir de represores mutuamente inhibitorios como E-KRAB y Pip-KRAB pueden convertir un pulso corto de DOX en un programa miogénico estable [6]. En la práctica, eso significa que el estado diferenciado permanece bloqueado después de la inducción en lugar de desvanecerse una vez que se elimina la señal. Agregar autorregulación negativa también ayuda a reducir el ruido estocástico y limitar la diferenciación filtrante durante la fase de expansión [6].
Esto es importante porque la diferenciación miogénica estándar in vitro a menudo se detiene a mitad de camino. Los índices de fusión suelen ser solo alrededor del 50–60%, lo que deja una gran población de reserva no fusionante [3]. La activación de MyoD impulsada por circuitos puede mejorar el compromiso, pero el compromiso por sí solo no garantiza una fusión uniforme. Cuando los circuitos de MyoD se combinan con la modulación de MEK, NOTCH y RXR, se hace posible una fusión casi completa en cultivo 2D [3]. El circuito controla el momento de la entrada en la línea; las pequeñas moléculas impulsan una salida del ciclo celular más limpia y uniforme.
3.2 Control adipogénico y fibrogénico para la composición y estructura
El músculo por sí solo no es suficiente. Una vez que el control miogénico está en su lugar, el siguiente problema es la composición: cuánto grasa se forma, cuánto ECM se deposita y cuándo se activan esos programas. Aquí, el control binario ON/OFF a menudo es demasiado brusco. Lo que los equipos suelen necesitar es una salida graduada, especialmente alrededor del punto de ramificación adipogénico-fibrogénico.
miSFITs ofrecen una forma práctica de ajustar la expresión en pasos.Al colocar sitios diana de miRNA mutados, por ejemplo, sitios para miR-17, en la 3′UTR de genes de salida como PPARG o BMP4, los investigadores pueden seleccionar niveles de expresión de una biblioteca de variantes [5]. Eso hace que la inducción de adipocitos sea más como un regulador de intensidad que un interruptor de luz. En lugar de empujar a las células a una respuesta de todo o nada, los equipos pueden titular la adipogénesis con más cuidado [5].
Los fibroblastos tampoco son solo espectadores aquí. Proporcionan proteínas ECM que dan forma a la textura [10]. Eso hace que el control fibrogénico sea parte del diseño del producto, no solo un problema secundario. Los circuitos pueden ayudar a gestionar el cambio entre estados fibrogénicos y adipogénicos, y en aves de corral cultivadas, puede ser necesario activar directamente PPARG en fibroblastos para generar una deposición de grasa significativa [10].
Una arquitectura de pajarita se adapta bien a este problema porque mantiene separados la detección y la salida. La capa de detección lee el estado actual de la célula, mientras que la capa de salida ajusta PPARG, CEBPA u otros reguladores de linaje. Esa separación ayuda a evitar que los programas adipogénicos o de tejido conectivo se activen antes de que las células hayan alcanzado la etapa de desarrollo adecuada.
3.3 Control de proporción multi-linaje y detección de retroalimentación
Los circuitos de control de proporción abordan un problema diferente: no si ocurre la diferenciación, sino si la mezcla final de la población se mantiene donde debería. Para productos de corte completo, obtener músculo, grasa y ECM en las proporciones correctas es tan importante como lograr que cualquier linaje se diferencie.
Estos sistemas incorporan control de retroalimentación en las propias células. Los promotores específicos de estado restringen la expresión de proteínas de señalización a las células que ya se han comprometido con un linaje dado.Los módulos paracrinos sintéticos permiten entonces que las células miogénicas comprometidas liberen una señal inhibitoria que suprime el compromiso adipogénico en las células cercanas. La lógica se asemeja a la inhibición lateral en los sistemas de desarrollo Delta-Notch [1] [6]. Donde la ramificación se complica más, las puertas de entrada multi-input pueden combinar señales extracelulares con señales de estado interno [9].
Los miSFITs también funcionan en esta etapa. Al ajustar la fuerza de salida de BMP4 u otros morfógenos, los equipos pueden cambiar el equilibrio de linaje sin reescribir la lógica de decisión ascendente. En redes reguladoras de genes diseñadas, este tipo de control aumentó la eficiencia de diferenciación del linaje objetivo del 52% en controles al 81% en iPSCs diseñadas [2]. En células madre mesenquimales modificadas, los circuitos sintéticos impulsaron la eficiencia de diferenciación cardíaca al 76% [2].
La tabla a continuación compara los enfoques de línea única y control de proporción utilizando los puntos que más importan en la producción.
| Característica | Circuito de una sola línea | Circuito de control de proporción |
|---|---|---|
| Complejidad | Baja; típicamente un solo promotor inducible y regulador [4] | Alta; requiere puertas lógicas y señalización paracrina [6][9] |
| Carga de monitoreo | Baja; usualmente sigue un solo reportero [4] | Alta; requiere seguimiento de múltiples marcadores de línea [5] |
| Robustez | Moderada; propensa a la heterogeneidad y formación de células de reserva [3] | Alto; utiliza retroalimentación e inhibición lateral para equilibrar poblaciones [1] |
| Valor de producción | Alto para biomasa; limitado para estructura de tejido complejo [10] | Esencial para productos de corte completo que necesitan músculo, grasa y ECM [4] |
Los circuitos de control de proporciones añaden una carga de validación más pesada. Pero su retroalimentación incorporada es difícil de igualar solo con el control del proceso, lo que pone más presión en la selección de clones y las pruebas de proceso.
4. Del constructo al proceso: validación, escalado y ajuste regulatorio
4.1 Estrategia de integración y selección de clones para un rendimiento estable
Después del diseño del circuito, comienza la parte difícil: introducir ese circuito en las células de una manera que se mantenga estable a través del pasaje a escala de producción.
La entrega lentiviral a menudo es eficiente y proporciona integraciones estables rápidamente. Pero la integración es aleatoria. Eso significa más atención regulatoria, además de un riesgo de que la expresión disminuya con el tiempo debido al silenciamiento. Los sistemas de transposones como PiggyBac y Sleeping Beauty se sitúan en el medio. Pueden mantener el rendimiento a través de muchos pasajes, pero aún es necesario examinar el número de copias y los sitios de inserción.Sleeping Beauty , por ejemplo, se ha utilizado para inmortalizar de manera estable células satélite bovinas mediante la sobreexpresión de TERT y CDK4, con líneas que retienen el potencial miogénico más allá de 40 pasajes [10]. CRISPR knock-in ofrece el control más estricto sobre dónde aterriza el constructo y el entorno genómico más preciso, aunque la selección de clones es más lenta y el rendimiento es menor.
| Método de Integración | Control de Inserción | Estabilidad | Escalabilidad | Consideraciones Regulatorias |
|---|---|---|---|---|
| Entrega Lentiviral | Baja (integración aleatoria) | Alta, pero propensa a silenciarse | Alta | Mayor escrutinio regulatorio debido a la inserción aleatoria y restos virales |
| Transposones (PiggyBac/SB) | Moderada | Alta en muchos pasajes | Alta | Requiere cribado para número de copias y sitios de inserción |
| Inserción CRISPR | Alta (específica del sitio) | Muy alta | Moderada | Favorable; reduce el riesgo de interrumpir genes endógenos |
| Vectores Episomales | Ninguno (extracromosómico) | Bajo; puede perderse durante la división | Bajo | Menor obstáculo de integración, pero inadecuado para la expansión a largo plazo |
La selección de clones necesita hacer más que confirmar que el constructo está presente.Debería rastrear la deriva de expresión, el perfil de inserción, la cinética de crecimiento, la eficiencia de diferenciación y la retención del fenotipo a un número alto de pasajes. snRNA-seq es útil aquí porque puede descartar clones enriquecidos para células de reserva Pax7⁺/Ki-67⁻ - células que salen del ciclo celular sin diferenciarse - antes de la ampliación [3]. Las EPSCs porcinas con un circuito Tet-On-PAX7 retuvieron una alta diferenciación muscular en microportadores 3D y cultivo en suspensión durante más de 40 pasajes [8].
4.2 Cómo cambia el comportamiento del circuito en cultivo 3D, microportador y biorreactor
Una vez que tienes un clon, la siguiente prueba es si se comporta de la misma manera fuera de 2D. En muchos casos, no lo hace. El rendimiento en 2D rara vez se traslada limpiamente a suspensión, microportador o cultivo en andamio porque los gradientes de difusión, los límites de oxígeno y el cizallamiento cambian la salida del circuito.
Uno de los primeros controles es la difusión del inductor. En los biorreactores de tanque agitado, los inductores de moléculas pequeñas necesitan alcanzar las células de manera uniforme. En la práctica, pueden formarse gradientes, especialmente en cultivos densos de microportadores y dentro de agregados o núcleos de andamios. El cultivo en suspensión suele ser más adecuado para la producción de carne cultivada a gran escala porque soporta una mayor densidad celular y ofrece un control de proceso más preciso.
El monitoreo del estado celular también se vuelve más difícil a medida que el sistema escala. Las señales de reporteros fluorescentes que son fáciles de leer por microscopía en 2D pueden volverse difíciles de resolver en construcciones opacas en 3D. Los temporizadores fluorescentes - sondas que cambian la emisión de azul a rojo a medida que una proteína madura - pueden proporcionar datos de activación de circuitos en tiempo real in situ [1]. La ruta sensata es la validación escalonada: primero en 2D, luego en construcciones 3D, y finalmente bajo las condiciones finales del biorreactor [3] [8].
4.3 Caracterización, documentación de seguridad alimentaria y consideraciones del Reino Unido/UE
Después de las pruebas del proceso, la caracterización debe mostrar que la función del circuito, el fenotipo y la seguridad siguen siendo válidos. El conjunto de datos principal debe incluir citometría de flujo, qPCR con secuencias cm, cursos de tiempo de RNA-seq y lecturas funcionales como la fracción de área de la cadena pesada de miosina y la expresión de mioglobina [1]. Se ha demostrado que los medios de diferenciación optimizados sin suero llevan la expresión de mioglobina a aproximadamente el 30% de los niveles encontrados en el músculo bovino nativo [3]. Eso proporciona a los equipos un punto de referencia claro en lugar de un objetivo vago.
También necesita documentar los perfiles de proteínas, aminoácidos y grasas, junto con las características sensoriales [10][3].
Desde el punto de vista regulatorio, el Reino Unido y la UE trazan una línea clara entre las líneas celulares inmortalizadas espontáneamente (no OGM) y las líneas genéticamente modificadas. Estas últimas necesitan expedientes de seguridad más amplios [10][3]. Los paquetes de estabilidad deben mostrar la retención del fenotipo y la estabilidad genómica a lo largo de todo el puente de producción - desde el banco maestro de células hasta las células de producción final - y los registros de trazabilidad deben dar cuenta de cada paso intermedio [10]. Si el circuito depende de un inductor químico, un aditivo seguro para alimentos o licenciado, como el ácido vanílico, es preferible a la doxiciclina [1].
La vigilancia genómica rutinaria es imprescindible, y un interruptor de suicidio o eliminación inducible debe documentarse como una medida central de control de riesgos [7]. Su función también debe incluirse en el expediente de seguridad, especialmente a medida que las normas del Reino Unido y la UE para la carne cultivada continúan tomando forma.
5. Hoja de ruta práctica y conclusión
5.1 Una hoja de ruta de implementación por fases para equipos de carne cultivada
La ruta más limpia desde el concepto hasta la producción es un flujo de trabajo escalonado.
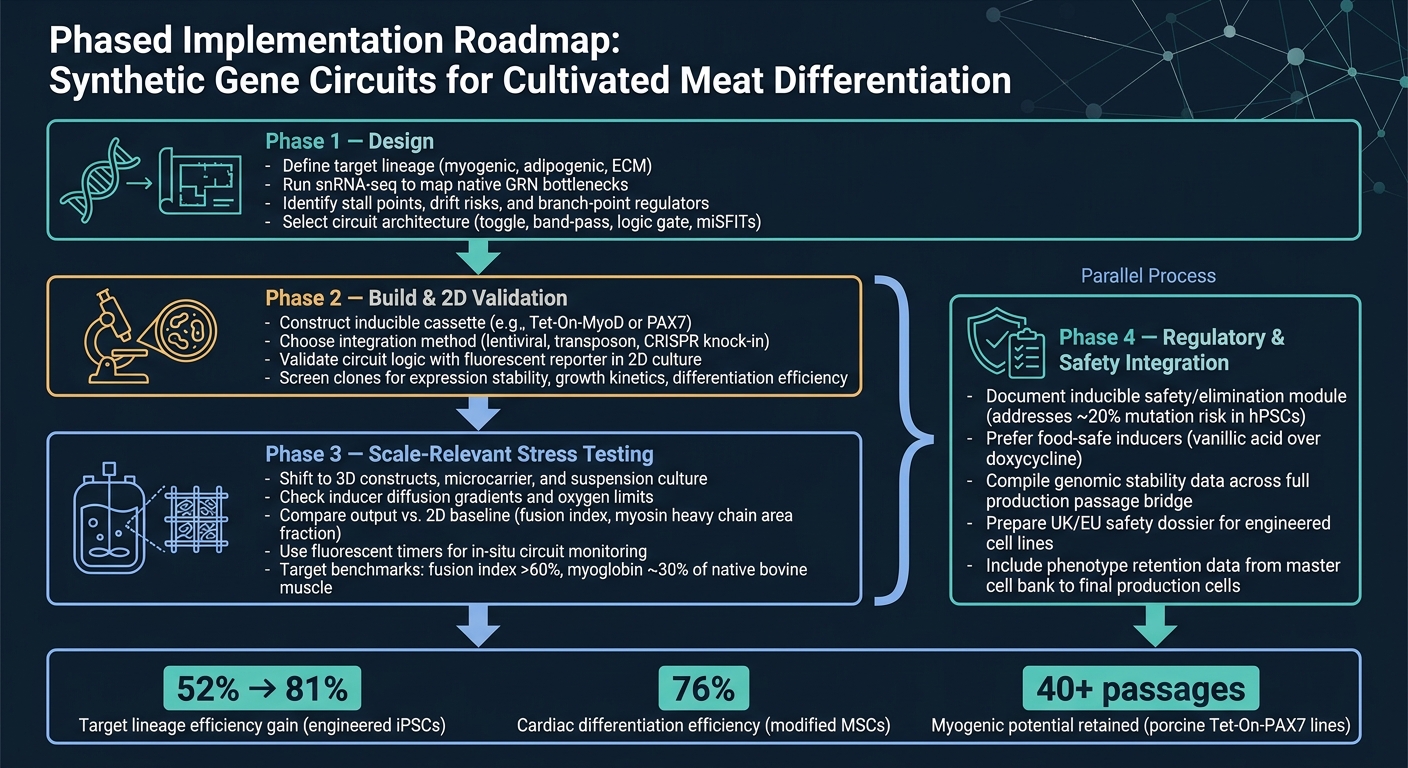
Fase 1 es diseño. Comience definiendo la línea objetivo, luego use snRNA-seq para confirmar los principales cuellos de botella antes de elegir una arquitectura de circuito. Ese paso es importante porque un circuito solo puede resolver las limitaciones que realmente ha identificado.
Fase 2 es construcción y validación 2D. Construya el constructo y verifique que el circuito se comporte como se espera en 2D, utilizando una lectura de reportero simple.En esta etapa, el objetivo es sencillo: confirmar que la lógica funciona antes de pasar a modelos más difíciles y costosos.
Fase 3 es la prueba de estrés relevante para la escala. Cambiar a sistemas 3D y condiciones relevantes para biorreactores, luego comparar la salida con la línea base 2D. Aquí es donde muchos diseños comienzan a mostrar sus puntos débiles, especialmente cuando entran en juego la transferencia de masa, el cizallamiento y los efectos de la matriz.
Fase 4 es la integración regulatoria y de seguridad, y debe ejecutarse en paralelo con la Fase 3. El trabajo de seguridad y regulación no debe esperar hasta el final. Ejecútelo junto con la ampliación, incluida la documentación de cualquier módulo de seguridad inducible.
5.2 Obtención de herramientas y materiales habilitadores a través de Cellbase
Una vez que el flujo de trabajo está establecido, la obtención a menudo se convierte en el paso limitante de la velocidad.
- líneas celulares
- medios sin suero y químicamente definidos
- andamios
- componentes de biorreactores
- sensores
- equipos analíticos
El acceso confiable a materiales compatibles en cada etapa tiene un efecto directo en la rapidez con la que se puede caracterizar el comportamiento del circuito bajo condiciones relevantes a escala.
5.3 Puntos clave
Los circuitos genéticos sintéticos brindan a los equipos de carne cultivada un control programable sobre el tiempo, los umbrales y el equilibrio de linaje que los protocolos solo de medios no pueden igualar. La elección de la arquitectura moldea la reversibilidad, la fuga y la seguridad.Los sistemas inducibles suelen ser preferidos porque proporcionan control condicional y una carga metabólica más baja [6].
"El conjunto de herramientas de biología sintética se puede utilizar para establecer líneas celulares con expresión génica ajustable, que, cuando se combinan con PAT y modelado computacional, pueden permitir sistemas de control en bucle cerrado para ofrecer un rendimiento y calidad óptimos del producto." - npj Systems Biology and Applications [6]
El despliegue exitoso no es solo un problema de biología. Depende de una estrecha conexión entre la ingeniería de circuitos, el diseño de bioprocesos, la documentación regulatoria y la adquisición.
Preguntas Frecuentes
¿Cómo mejoran los circuitos genéticos sintéticos la consistencia de la diferenciación?
Los circuitos genéticos sintéticos pueden hacer que la diferenciación sea más consistente porque te dan control programado sobre el comportamiento celular y el compromiso de linaje.En la práctica, eso significa usar operaciones lógicas modulares para ajustar la expresión de genes y factores de transcripción con una sincronización precisa.
Esa sincronización importa. Ayuda a las células a pasar por cambios de estado definidos en el orden correcto, en lugar de derivar hacia destinos mixtos o no deseados. También reduce la diferenciación fuera del objetivo y disminuye el ruido en toda la cultura.
El beneficio es directo: poblaciones celulares más uniformes, estables y maduras para la producción de carne cultivada.
¿Qué diseño de circuito se adapta al control miogénico o adipogénico?
En la investigación de carne cultivada, los mismos fibroblastos de pollo pueden ser dirigidos hacia cualquiera de las dos líneas. La miogénesis sigue un conjunto de protocolos de inducción, mientras que la adipogénesis se puede activar exponiendo las células a insumos como suero de pollo o ácidos grasos.
A partir de ahí, estos destinos celulares pueden ser controlados paso a paso dentro de andamios de hidrogel 3D para construir estructuras de carne con proporciones definidas de grasa y colágeno.
¿Por qué los circuitos genéticos a menudo se comportan de manera diferente en cultivos 3D?
En cultivos 3D , los circuitos genéticos a menudo se comportan de manera diferente porque las células están lidiando con entradas físicas y estructurales que simplemente no existen en monocapas 2D. Estas entradas incluyen tensión mecánica, estrés de cizallamiento, rigidez de la matriz y densidad celular local.
Estas señales pueden cambiar vías de señalización como Notch. También pueden cambiar cómo los circuitos sintéticos detectan la fuerza y coordinan respuestas posteriores, incluyendo la adhesión célula-célula y la morfogénesis tisular.









