

CRISPR משנה את ייצור הבשר המתורבת על ידי התמודדות עם אתגר מרכזי: לחץ תאי בביו-ריאקטורים תעשייתיים. כלי זה מאפשר עריכות גנטיות מדויקות לשיפור הישרדות התאים, הארכת התרבות והפחתת הזדקנות בתנאים קשים. לדוגמה, השבתת גנים כמו TP53 ו- PTEN האריכה את משך התרבות קווי תאים ראשוניים לעומת קווי תאים בני אלמוות מ-100 ל-200 ימים והגדילה את שפע התאים פי 1,000 תוך 30 ימים. עם זאת, שינויים אלו יכולים להשפיע על התמיינות, ולכן נדרשת אופטימיזציה זהירה.
תובנות מרכזיות מהמאמר כוללות:
- גורמי לחץ בביו-ריאקטורים: כוחות גזירה, חוסר איזון תזונתי ולחץ חמצוני מפחיתים את חיות התאים.
- אסטרטגיות CRISPR: השבתת גנים (TP53, PTEN) והפעלות (HIF1A) מכוונות לתגובות לחץ ספציפיות.
- אימות: תאים שעברו עריכה עוברים בדיקות גנומיות, פרוטאומיות ופונקציונליות כדי להבטיח ביצועים ופוטנציאל התמיינות.
-
הגדלה: מעבר לתנאי ביוריאקטור כולל מדיה וציוד מותאמים, עם פלטפורמות כמו
Cellbase המספקות משאבים מותאמים.
הדיוק של CRISPR מאפשר פיתוח קווי תאים עמידים ללחץ, אך איזון בין צמיחה והתמיינות נותר קריטי לייצור בשר מתורבת בקנה מידה.
מיפוי פרופילי לחץ ביוריאקטור לעיצוב גנטי
זיהוי גורמי לחץ מרכזיים בביוריאקטור
לפני תחילת עריכת CRISPR, חשוב למפות פרופילי לחץ ביוריאקטור כדי להנחות את העיצוב הגנטי. גורמי לחץ בביוריאקטורים מעוררים תגובות תאיות ספציפיות שיש להבין היטב כדי לבחור מטרות גנטיות מתאימות.
מתח מכני והידרודינמי הוא אחד האתגרים המיידיים ביותר. ביוריאקטורים עם מיכל מעורב יוצרים כוחות גזירה שיכולים לפגוע בממברנות התאים ולהפריע לנתיבי האיתות התאי [5][2]. מתחים תזונתיים ומטבוליים גם הם משחקים תפקיד מרכזי, לעיתים קרובות נובעים מנטילת תזונה לא אחידה. גרדיאנטים תזונתיים בשלדים תלת-ממדיים והצטברות אמוניה תורמים למתח מטבולי [3][5][6]. בנוסף, תנודות ב-pH וטמפרטורות מוגברות יכולות להפחית את שיעורי התרבות התאים ואפילו לדחוף תאים לכיוון התמיינות מוקדמת [3][2].
מתחים אחרים, כולל מתחים חמצוניים, מיטוכונדריאליים ומתחים של ה-ER, מאתגרים עוד יותר את חיות התאים. עקה חמצונית הופכת לחמורה במיוחד במהלך המעבר למדיה ללא סרום, שכן היעדר נוגדי חמצון טבעיים משאיר את התאים פגיעים יותר למיני חמצן תגובתיים [4]. ברמה התאית, עקה מיטוכונדריאלית ו-עקה של הרשתית התוך-פלזמית (ER) מתעוררות כאשר תנאי הביופרוסס חורגים מהטווחים האופטימליים שלהם [6]. שיאויאן גואו מהמכון למחלות נוירודגנרטיביות ב-UCSF מדגיש את הדינמיקה הזו:
"בנוכחות לחצים פיזיולוגיים וסביבתיים שונים, תאים מתחילים במהירות תגובות עקה כדי לשחזר את ההומאוסטזיס התאי." [6]
על ידי מיפוי יזום של גורמי עקה אלה, במקום להגיב לבעיות כשהן מתעוררות, חוקרים יכולים להגדיר מטרות מדויקות להנדסה גנטית.גישה שיטתית זו מבטיחה כי אסטרטגיות CRISPR מכוונות לפיתוח קווי תאים עמידים ללחץ בצורה יעילה.
שימוש בנתוני אומיקס למציאת גנים תגובתיים ללחץ
לאחר אפיון סביבת הלחץ, השלב הבא הוא זיהוי הגנים המגיבים לתנאים אלו. כלים כמו טרנסקריפטומיקה (RNA-seq) ו-פרוטאומיקה הם בעלי ערך רב למעקב אחר שינויים בביטוי גנים ושפע חלבונים כאשר תאים עוברים ממצבים בריאים, מוקדמים, למצב לחוץ, מאוחר [1][6]. עם זאת, בעוד ששיטות אלו לוכדות את ההשפעות במורד הזרם, הן לעיתים קרובות נכשלות בזיהוי הרגולטורים במעלה הזרם המניעים שינויים אלו [6].
מסכי נוקאאוט CRISPR מאוגדים מגשרים על הפער הזה.על ידי שיבוש שיטתי של אלפי גנים באוכלוסיית תאים גדולה, מסכים אלו חושפים אילו שיבושי גנים מעניקים יתרון גדילה תחת לחץ, ומגלים מרכזי רגולציה קריטיים [1][6]. לדוגמה, מיקוד בגנים כמו TP53 ו-PTEN הוכח כמסוגל להפוך חתימות הזדקנות מולקולריות שנגרמות על ידי לחץ תרבות ממושך. זה מאפשר לתאים במעבר מאוחר לשמור על פרופיל תעתוקי דומה לתאים מסוג פראי במעבר מוקדם [1].
באמצעות אשכולות היררכיים, חוקרים יכולים לקבץ גנים על בסיס שינויים בביטוי שלהם לאורך זמן, ולבודד מודולים הקשורים לתהליכים כמו התקדמות מחזור התא וסינתזת חלבונים. תהליכים אלו בדרך כלל יורדים כאשר הזדקנות הנגרמת על ידי ביוריאקטור תופסת [1]. כאשר משולבים עם ניתוח העשרת מסלולים (באמצעות כלים כמו gprofiler2 ), ניתן לקשר את המודולים הללו למסלולים ביולוגיים ספציפיים, כגון איתות TGFβ או התמיינות כונדרוגנית, אשר עשויים להגביל באופן פעיל את התפשטות התאים [1].
הטבלה למטה מפרטת את התרומות של כל שיטה לבניית מפת לחץ מקיפה:
| שיטה | שימוש עיקרי | פלט מפתח |
|---|---|---|
| תעתוק (RNA-seq) | מדידת שינויים בביטוי mRNA | גנים מבוטאים באופן דיפרנציאלי (DEGs) בין תאים בלחץ וללא לחץ[1] |
| פרוטאומיקה | מדידת שפע חלבונים | פלטים תרגומיים ממופים לגורמי לחץ ספציפיים[6] |
| מסך CRISPR משולב | הפרעה פונקציונלית בגנים | מרכזי רגולציה עליונים וגנים קריטיים לכושר[1][6] |
| PCA & אשכול היררכי | הדמיית נתונים וקיבוץ | שינויים במצב התאי ונתיבי תגובה ללחץ המשותפים [1] |
sbb-itb-ffee270
הנדסת קווי תאים עם CRISPR-Cas9 - טיפים וטריקים למקסום הצלחה
אסטרטגיות CRISPR להנדסת קווי תאים עמידים ללחץ
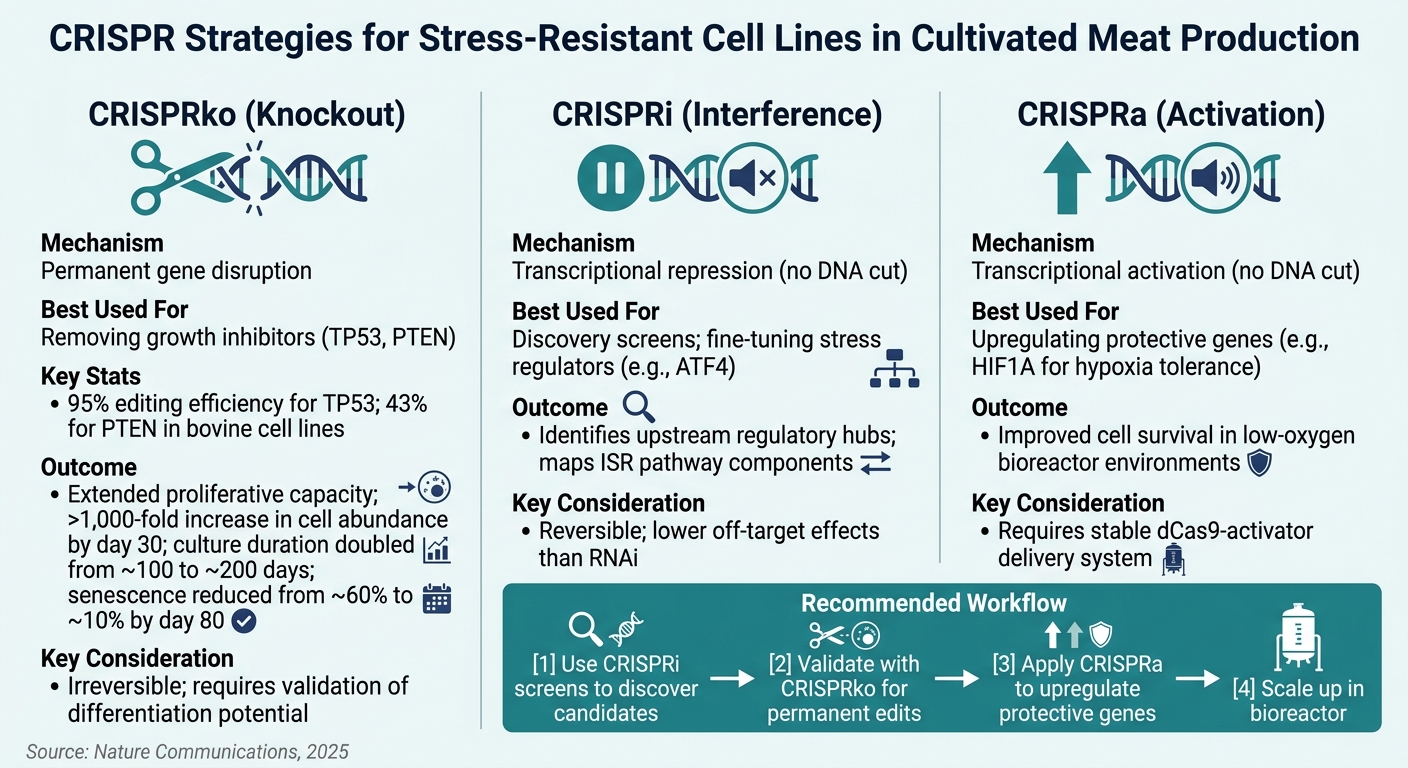
טכניקות CRISPR לקווי תאים עמידים ללחץ בבשר מתורבת
גנים ונתיבים מרכזיים לעמידות ללחץ
עם מפת לחץ מפורטת ביד, השלב הבא הוא לזהות גנים יעד לעריכה.בחירת המטרות תלויה בגורם הלחץ הראשי המשפיע על ביצועי התאים.
הזדקנות רפליקטיבית היא מכשול משמעותי בייצור בשר מתורבת, שכן היא מגבילה את התרבות התאים. כ-25% ממקורות התאים בתחום זה הם תאי גזע מזנכימליים (MSCs), אשר מתמודדים עם עצירת גדילה בלתי הפיכה לאחר מעבר חוזר [1] . נוק אאוט של TP53, הגן המקודד לחלבון מדכא הגידול p53, מתמודד ישירות עם בעיה זו. מחקר בתאי גזע מזנכימליים של בקר מראה כי נוק אאוט של TP53 מאריך משמעותית את יכולת ההתרבות של התאים, ומאפשר להם להתחלק הרבה מעבר למגבלות של קווים לא ערוכים [1] . באופן דומה, נוק אאוט של PTEN משפר את מסלול PI3K/AKT/mTOR, ומגביר את עמידות הלחץ [1] .
לטיפול בלחצים מטבוליים ומיטוכונדריאליים, תגובת הלחץ המשולבת (ISR) היא מסלול קריטי. גורם השעתוק ATF4 משחק תפקיד מרכזי בתיאום תגובות לחץ מיטוכונדריאליות, ומסכי CRISPR היו חיוניים במיפוי הרגולטורים העיליים שלו [6] . כפי שמסבירים שיאויאן גואו ומרטין קמפמן מאוניברסיטת קליפורניה, סן פרנסיסקו:
"מסכי גנטיקה בלתי מוטים המבוססים על כתב שעתוק או תרגום הם גישות חזקות לזיהוי גורמי רגולציה של תגובת לחץ ספציפית." [6]
גם מסלול TGFβ ראוי לתשומת לב, במיוחד להרחבת MSCs של בקר. מסכי CRISPR הראו כי התמיינות כונדרוגנית מונעת TGFβ מדכאת את התרבות התאים.דיכוי מסלול זה מסייע לשמור על תאים במצב בלתי ממויין וניתן להרחבה [1] . בתנאים היפוקסיים הנמצאים לעיתים קרובות בליבות הצפופות של פיגומים תלת-ממדיים, הפעלת HIF1A באמצעות CRISPRa משפרת את הישרדות התאים בסביבות דלות חמצן. שינויים אלה מציידים את התאים לשגשג בתנאים הדינמיים של ביוריאקטורים בקנה מידה תעשייתי.
עם זאת, חשוב לציין כי עריכות הממקסמות את התרבות התאים - כמו TP53 נוקאאוט - יכולות להפחית את יכולת התאים להתמיין לרקמת שריר או שומן. יש לאזן בזהירות את הפשרה בין פוטנציאל הצמיחה וההתמיינות בעת תכנון אסטרטגיית הנדסה [1] .
| גורם לחץ | מטרה גנטית מרכזית | אסטרטגיית CRISPR | תוצאה |
|---|---|---|---|
| הזדקנות רפליקטיבית | TP53 | נוקאאוט | יכולת פרוליפרטיבית מורחבת; עלייה בכמות התאים |
| לחץ תזונתי/גדילה | PTEN | נוקאאוט | הגברת איתות PI3K/AKT/mTOR; שיפור הישרדות |
| לחץ מיטוכונדריאלי | ATF4 | CRISPRi / כתב | זיהוי מסלולי רגולציה עיליים |
| היפוקסיה | HIF1A | CRISPRa (הפעלה) | הגברת הישרדות בסביבות ביוריאקטור עם חמצן נמוך |
| סטייה כונדרוגנית | מסלול TGFβ | נוקאאוט / דיכוי | תחזוקה של מצב לא ממויין, פרוליפרטיבי בתאי גזע מזנכימליים של בקר |
לאחר זיהוי הגנים המרכזיים, בחירת הטכניקה הנכונה של CRISPR הופכת לשלב הקריטי הבא.
השוואת טכניקות עריכת CRISPR
הבחירה בשיטת CRISPR קובעת את הדיוק והקביעות של השינויים הגנטיים. לכל גישה יש יתרונות ייחודיים בהתאם למטרה, אם היא שינוי קבוע, התאמה הפיכה או סריקה חקרית.
CRISPR knockout (CRISPRko) היא השיטה המועדפת להשבתה קבועה של גנים. היא אידיאלית למטרות כמו TP53 ו-PTEN, שבהן נדרש אובדן תפקוד מוחלט. מחקרי אימות הראו ש-CRISPRko משיגה יעילות עריכה של 95% עבור TP53 ו-43% עבור PTEN בקווי תאים של בקר [1] . שינויים אלו מדגישים את החשיבות של בדיקת יעילות ספציפית למטרה לפני המשך לעריכה בקנה מידה גדול.
CRISPR interference (CRISPRi) מציעה דיכוי גנים הפיך, מה שהופך אותה לאידיאלית לשלבי גילוי.זה גם מפחית את ההשפעות הלא מכוונות בהשוואה ל-RNAi [6]. מצד שני, הפעלת CRISPR (CRISPRa) פועלת על ידי ביטוי יתר של גנים מגנים, כמו אלה המעורבים בסבילות להיפוקסיה (HIF1A) או בהגנה נוגדת חמצון, כדי לשפר את עמידות הלחץ.
להלן השוואה מהירה של הטכניקות:
| טכניקה | מנגנון | הכי טוב לשימוש עבור | שיקול מפתח |
|---|---|---|---|
| CRISPRko | שיבוש גנים קבוע | הסרת מעכבי גדילה (TP53, PTEN) | בלתי הפיך; דורש אימות של פוטנציאל התמיינות |
| CRISPRi | דיכוי תעתוק (ללא חיתוך DNA) | מסכי גילוי; כוונון עדין של רגולטורים | הפיך; פחות השפעות מחוץ למטרה מאשר RNAi |
| CRISPRa | הפעלת תעתוק (ללא חיתוך DNA) | הגברת גנים מגנים (HIF1A) | דורש מערכת אספקה יציבה של dCas9-activator |
עבור צוותים בשלבים הראשונים של זיהוי מטרות, סריקות CRISPRi משותפות מספקות דרך חסכונית לגלות גנים לעמידות בפני לחץ בקנה מידה גדול. ברגע שמועמדים מבטיחים מאומתים, ניתן להשתמש ב-CRISPRko לעריכות קבועות המתאימות לייצור. גישות אלו משלימות זו את זו, ושימוש בהן ברצף נחשב יותר ויותר כפרקטיקה מיטבית בתחום [1][6].
לצורך השגת ריאגנטים של CRISPR ואספקת ביוריאקטורים המותאמים למחקר בשר מתורבת, פלטפורמות כמו
יישום ואימות קווי תאים שעברו עריכה באמצעות CRISPR
תכנון והעברת עריכות CRISPR
לאחר שזיהית את הגנים המטרה, השלב הבא הוא תכנון והעברת עריכות CRISPR. כדי להבטיח שיבוש גנים יעיל, התמקד ביצירת RNAs חד-מדריכים (sgRNAs) המכוונים לאקסונים חיוניים. גישה זו מגדילה את הסבירות לשיבוש מלא של הגן במקום לייצר חלבון מקוצר וחלקית פונקציונלי.שימוש באסטרטגיית RNA כפולה יכול לשפר משמעותית את יעילות הנוקאאוט, להעלות אותה מכ-55% ליותר מ-95% [8].
שיטת המסירה שתבחר תלויה בסוג התא הספציפי. עבור קווי תאים של בשר מתורבת, ריבונוקלאופרוטאינים (RNPs) של Cas9 מורכבים מראש הם לעיתים קרובות האפשרות הטובה ביותר. RNPs אלה הם זמניים, כלומר הם מתפרקים במהירות לאחר המסירה, מה שעוזר למזער השפעות מחוץ למטרה ולהימנע מהסיכון של אינטגרציה של DNA פלסמידי [8]. במקרים שבהם מעורבים מסכים מאוגדים או קווי תאים ראשוניים שקשה להחדיר, טרנסדוקציה לנטיויראלית היא חלופה אמינה. כאשר משתמשים במערכות לנטיויראליות, חוקרים בדרך כלל שומרים על ריבוי זיהום (MOI) נמוך של כ-0.3 כדי להימנע מאינטגרציות מרובות, שעלולות לסבך את הניתוח בהמשך [1].
לתוצאות מיטביות, ודא שהתאים נמצאים בשלב הצמיחה הלוגריתמית ובכיסוי של 70–90% לפני הטרנספקציה. לאחר המסירה, יש לבודד שיבוטים בודדים באמצעות שיטות כמו דילול מגביל או מיון תאים מופעל פלואורסצנטי (FACS) כדי להבטיח אימות ברור וחד משמעי. לבסוף, יש לאמת את העריכות ברמות הגנומיות, הפרוטאומיות והתפקודיות כדי לאשר הצלחה.
סינון ואימות קווי תאים ערוכים
אימות יסודי הוא חיוני כאשר מעבירים קווי תאים ערוכים לתנאי ביוריאקטור. תהליך זה כולל סינון בשלוש רמות: גנומית, פרוטאומית ותפקודית. דילוג על כל אחד מהשלבים הללו מגדיל את הסיכון לבחירת קווי תאים שעלולים להיכשל בתנאי ייצור.
ברמה הגנומית, ניתן לבצע סינון ראשוני באמצעות מבחני אי התאמה כמו T7E1 או Surveyor, המספקים הערכה מהירה של תדירות העריכה בבריכת התאים.לאישור מדויק, יש לעקוב עם ריצוף סנגר או ריצוף מהדור הבא (NGS) כדי לזהות שיבוטים עם אינדלים משבשים ביאלליים [7][8]. אימות פרוטאומי, שבדרך כלל מבוצע באמצעות ניתוח Western blot, מבטיח את היעדרות המלאה של חלבון המטרה. לדוגמה, מחקר שנערך בשנת 2025 הראה כי נוקאאוט של TP53 הוביל לעלייה של פי 1,000 בשפע התאים עד היום ה-30 של מסך תחרותי, מה שהכפיל למעשה את משך התרבות מ-100 לכ-200 ימים [1].
אימות פונקציונלי חשוב באותה מידה. ניתן להעריך את החיוניות המטבולית וקצב ההתרבות באמצעות מבחני Alamar Blue, בעוד מעקב אחר זמן הכפלת האוכלוסייה (PDT) לאורך תקופות ממושכות - עד 200 ימים - מסייע לזהות קווי תאים שהתמודדו עם הזדקנות רפליקטיבית [1]. עבור קווי תאים שהונדסו לעמוד בלחץ היפוקסי או מיטוכונדריאלי, מבחני דיווח מבוססי FACS יכולים לאשר שהתאים מגיבים כראוי בתנאים של חוסר חמצן או מגבלות תזונתיות [6]. בנוסף, יש לבדוק קווי תאים עם נוקאאוטים של TP53 או PTEN עבור יכולתם לשמור על פוטנציאל התמיינות. ציטומטריה בזרימה עבור סמני תאי גזע מזנכימליים (MSC) כגון CD29 ו-CD44 יכולה לאמת שהתאים הללו שומרים על תכונות הגזע שלהם [1].
| רמת אימות | שיטה | מטרה |
|---|---|---|
| גנומית | ריצוף סנגר / NGS | אישור אינדלים משבשים ביאלליים[7][8] |
| פרוטאומית | בלוט מערבי | אימות היעדר מוחלט של חלבון המטרה[7][8] |
| פנוטיפית | ציטומטריית זרימה (CD29/CD44) | בדיקת שמירה על סמני MSC וסטמנס[1] |
| פונקציונלית | Alamar Blue / מעקב PDT | הערכת קינטיקה של צמיחה ובריאות מטבולית[1] |
| לחץ | מבחני דיווח מבוססי FACS | בדוק את התנהגות התגובה ללחץ בתנאים מאתגרים[6] |
לפני הגדלת קו תאים ערוך, בצע פרופיל STR כדי לאשר את זהות התאים ובצע בדיקת מיקופלסמה כדי לשלול זיהום[7]. יצירת קו תאים נוקאאוט מאומת בדרך כלל דורשת כשלושה חודשים, עם אפשרות לחזור על שלבים מסוימים בתהליך העבודה.
הגדלת היקף: העברת קווי תאים עמידים ללחץ לייצור
מעבר קווי תאים ערוכים לתנאי ביוריאקטור
לאחר האימות, קווי תאים ערוכים חייבים לעבור מתרביות מעבדה בקנה מידה קטן למערכות השעיה כמו ביוריאקטורים עם מערבל, ריאקטורים עם הרמת אוויר, או כלים עם קיר מסתובב - כל אחד מהם מסוגל לתמוך בייצור בשר מתורבת בקנה מידה תעשייתי [2].
עבור תאים התלויים בהידבקות כמו תאי גזע מזנכימליים של בקר (bMSCs), שימוש במיקרונשאים מצופים בלמינין-511 מציע דרך מעשית לתרבות השעיה [3]. במהלך המעבר הזה, חשוב לעקוב אחר סמני MSC כמו CD29 ו-CD44 כדי להבטיח שהתאים שומרים על הפוטנציאל שלהם להתמיינות [1].
שלב קריטי בהגדלת היקף כולל שינוי נוסחת המדיה. מדיה מבוססת סרום צריכה להיות מוחלפת בנוסחאות מוגדרות כימית, ללא סרום, מועשרות בליפידים, חומצות אמינו לא חיוניות ונוגדי חמצון כדי לשמור על חיות התאים בתנאים בקנה מידה גדול [4]. במיוחד, קווי תאים שעברו עריכה באמצעות CRISPR עם נוקאאוטים של TP53 ו-PTEN מצוידים טוב יותר למעבר זה. מחקר שפורסם ב-Nature Communications (2025) הראה כי עריכות אלו האריכו את תוחלת החיים הפרוליפרטיבית של bMSCs מכ-100 ימים ליותר מ-200 ימים, תוך הפחתת הזדקנות מכ-60% ל-10% בלבד עד היום ה-80 [1].
"נוקאאוטים של TP53 ו-PTEN הגדילו משמעותית את שיעורי הפרוליפרציה ועיכבו את ההזדקנות." - Nature Communications [1]
במהלך המעבר, כלים כמו מבחני Alamar Blue ו-qRT-PCR הם חיוניים למעקב אחר חיות התאים ולהבטחת יציבות השינויים הגנטיים. קווי התאים הבקריים שנערכו באמצעות CRISPR הראו שיפור ממוצע של 12% בקצב הכפלה, כאשר חלקם הגיעו לעלייה של 50% עד היום ה-50 [1] . ברגע שהתאים מראים ביצועים יציבים בתנאי ביוריאקטור, ניתן להתמקד ברכישת הציוד המיוחד הנדרש להגדלה.
רכישת ציוד וחומרים להגדלה
הגדלה לריצות ביוריאקטור ברמת ייצור מציגה אתגרים משמעותיים ברכש. לאחר אישור התאמת התאים, רכישת החומרים והציוד הנדרשים הופכת לעדיפות.פריטים כמו ביוריאקטורים חד-פעמיים עם מיכל ערבוב, מיקרונשאים מאומתים, רכיבי מדיה ללא סרום ומערכות FACS למעקב מתמשך אחר שיבוטים הם מאוד מתמחים ולעיתים קרובות אינם זמינים מספקי מעבדות כלליים.
פלטפורמות כמו
סיכום
טכנולוגיית CRISPR עברה מכלי מחקר לשיטה מעשית להנדסת קווי תאים בייצור בשר מתורבת. על ידי מיקוד ברגולטורים מרכזיים כמו TP53 ו-PTEN, חוקרים האריכו משמעותית את התרבות התאים, והכפילו למעשה את משך התרבות הטיפוסי [1]. התקדמות זו דוחפת את גבולות הייצור בקנה מידה גדול לבשר מתורבת.
עם זאת, המסע מקווי תאים ערוכים לייצור בקנה מידה מלא דורש אימות יסודי בכל שלב. הבטחת שמירה על יכולת ההנדסה של התאים להתמיין לרקמות שריר ושומן היא קריטית בדיוק כמו השגת התרבות מהירה. ללא זאת, אפילו קווי התאים הצומחים במהירות לא יהיו בעלי כדאיות מסחרית [1]. זה מדגיש את הצורך בתהליכי אימות מחמירים כדי לאשר שהתרבות משופרת מתורגמת לתוצאות ייצור משמעותיות.
Nature Communications מחזקת גישה זו, ומציינת:
"ממצאים אלה מדגימים את התועלת של סריקת CRISPR לאופטימיזציה של תכונות תאי גזע בקר ומציעים דרך לייצור בשר מתורבת בקנה מידה גדול יותר בעתיד." [1]
למרות ההתקדמות הזו, אתגרים מעשיים כמו רכש יכולים לעכב את ההתקדמות. תלות בספקים כלליים לספריות sgRNA, ביוריאקטורים חד-פעמיים ומדיה ללא סרום לעיתים קרובות מציגה בעיות תאימות ועיכובים. פלטפורמות כמו
הזמינות של חומרים מתאימים חשובה לא פחות מההנדסה הגנטית עצמה. כפי שצוין על ידי Nature Communications, בעוד שבשר מתורבת מציג חלופה מבטיחה לבשר קונבנציונלי, יכולת ההגדלה ויעילות העלות נשארות מכשולים משמעותיים. הנדסה מבוססת CRISPR, כאשר היא משולבת עם עיצוב ביופרוסס ממושמע ורכש יעיל דרך פלטפורמות כמו
שאלות נפוצות
אילו לחצים של ביוריאקטור עלי לפרופיל לפני בחירת מטרות CRISPR?
בעת בחירת מטרות CRISPR לפיתוח קווי תאים עמידים ללחץ בייצור בשר מתורבת, חשוב להעריך את הלחצים העיקריים של הביוריאקטור המשפיעים על צמיחת התאים והישרדותם. לחצים אלו כוללים:
- לחץ גזירה: תאים בביוריאקטורים נחשפים לעיתים קרובות לכוחות מכניים מערבוב ואוורור. לחץ גזירה ממושך יכול לפגוע בממברנות התאים ולפגוע בצמיחה.
- רמות חמצן: שמירה על ריכוזי חמצן אופטימליים היא חיונית. מעט מדי חמצן יכול להגביל את ייצור האנרגיה, בעוד שחמצן מוגזם עלול להוביל ללחץ חמצוני.
- זמינות חומרים מזינים: תאים דורשים אספקה עקבית של חומרים מזינים. כל חוסר איזון או דלדול יכולים לעכב את ההתרבות והפרודוקטיביות.
- תנודות ב-pH: תאים משגשגים בטווח pH צר. סטיות יכולות לשבש תהליכים מטבוליים ופעילות אנזימטית.
- שינויים בטמפרטורה: אפילו שינויים קלים בטמפרטורה יכולים להשפיע על תפקודי התאים, לגרום ללחץ או להפחית את החיוניות.
- הצטברות פסולת: תוצרי לוואי מטבוליים, אם לא מוסרים ביעילות, יכולים להפוך לרעילים ולעכב את גדילת התאים.
על ידי הבנה מעמיקה של גורמי הלחץ הללו, חוקרים יכולים לזהות מסלולי תגובה ללחץ קריטיים. ידע זה מאפשר שינויים גנטיים ממוקדים באמצעות CRISPR, משפר את עמידות קווי התאים ומבטיח ביצועים חזקים יותר בתנאי ביוריאקטור.
איך אני מאזן עריכות צמיחה מהירה עם הבחנה בין שריר לשומן?
איזון בין צמיחה מהירה להבחנה בין שריר לשומן בייצור בשר מתורבת דורש שליטה קפדנית בגנטיקה ובתנאי התרבות. טכנולוגיית CRISPR משחקת תפקיד מרכזי כאן, ומאפשרת שינויים ממוקדים בגנים כמו TP53 ו-PTEN. שינויים אלו יכולים לקדם את התרבות התאים תוך שמירה על יכולת התאים להבחין לשריר ולרקמת שומן.
כיוונון עדין של תנאי התרבות ורגולציה של ביטוי גנים הם קריטיים באותה מידה להשגת האיזון הרצוי. משאבים כמו
מהי האימות המינימלי הנדרש לפני הגדלת קנה המידה של הביוראקטור?
לפני המעבר לביוראקטורים, חשוב לאשר שקווי תאים מהונדסים גנטית שומרים על תכונות יציבות ורצויות, כגון שיעורי גידול משופרים, סבילות ללחץ ויכולת התמיינות. תהליך האימות הזה צריך להעריך את היציבות הגנטית ולהבטיח ביצועים עקביים בתנאי ביופרוסס. נתונים תומכים מניתוח מולטי-אומיקס ופרופיל תגובת לחץ הם מפתח להערכה זו. שימוש בסריקת CRISPR בתפוקה גבוהה יכול לזהות עריכות גנטיות שמשפרות את התרבות התאים ותוחלת החיים, מה שהופך את קווי התאים הללו ליותר מתאימים לייצור בשר מתורבת בקנה מידה גדול.










