

אם אתה יכול להרחיב תאים אך לא יכול להעביר אותם לגורל הנכון בזמן הנכון, התהליך שלך יתקע בשלב ההתמיינות. זהו הנקודה המרכזית כאן: מעגלים גנטיים סינתטיים נותנים לך שליטה בתוך התא על התחייבות, תזמון, זיכרון ותערובת שושלת, כאשר שינויים במדיה בלבד לעיתים קרובות משאירים אוכלוסיות הטרוגניות, חלקית מחויבות.
אם הייתי בונה תהליך עבודה להתמיינות בשר מתורבת, הייתי לוקח ארבע נקודות מהמאמר הזה מיד:
- התחל עם הרשת הטבעית, לא עם המבנה. השתמש ב- snRNA-seq, ניתוח מסלול, הסקת GRN ופרופילינג miRNA כדי למצוא היכן תאים נתקעים, נסחפים או מסתעפים לגורל הלא נכון.
-
התאם את סוג המעגל לבעיה בתהליך.
מתג toggle switch מתאים לנעילה, עיצוב feedforward או band-pass מתאים לשליטה בתזמון, שער לוגי logic gate מתאים לשער רב-אותות, ו- miSFITs מתאימים לפלט מדורג. - תכנן לדליפה נמוכה, רעש נמוך ובטיחות מהיום הראשון. חלקים אורתוגונליים, אוטורגולציה שלילית, iFFLs, טרנסגנים cm ומודול הרג או עצירת צמיחה מושרה הם חלק מהבנייה, לא מחשבה שנייה.
- אמת בתנאים רלוונטיים לקנה מידה מוקדם. מעגל שעובד ב-2D עשוי להשתנות ב- 3D, מיקרונשא או השעיה מעורבלת בגלל גרדיאנטים של משרה, מגבלות חמצן וגזירה.
המאמר גם מציין נקודה מעשית שחשובה לצוותי תהליך: שליטה בקו יחיד ושליטה ביחס הם עבודות שונות. קלטת Tet-On MyoD עשויה לדחוף כניסה מיוגנית, אך מוצרים שלמים דורשים שליטה על פרופורציות של שריר, שומן ו-ECM, שבדרך כלל משמעותם משוב, איתות פאראקריני וסינון שיבוטים כבד יותר.
כמה נתונים מעגנים את המסר הזה:
- התמיינות מיוגנית סטנדרטית יכולה להיעצר עם אינדקסי היתוך של בערך 50–60%
- GRNs מהונדסים ב-iPSCs הגדילו את התמיינות קו המטרה מ-52% ל-81%
- מעגלים סינתטיים ב-MSCs מותאמים הניעו התמיינות לבבית ל-76%
- חלק מקווי Tet-On-PAX7 חזיריים שמרו על פוטנציאל מיוגני גבוה מעבר ל-40 מעברים
- בערך 20% מתאי גזע פלוריפוטנטיים אנושיים עשויים לשאת מוטציות הקשורות לסרטן, ולכן מודולי בטיחות ניתנים להשראה חשובים
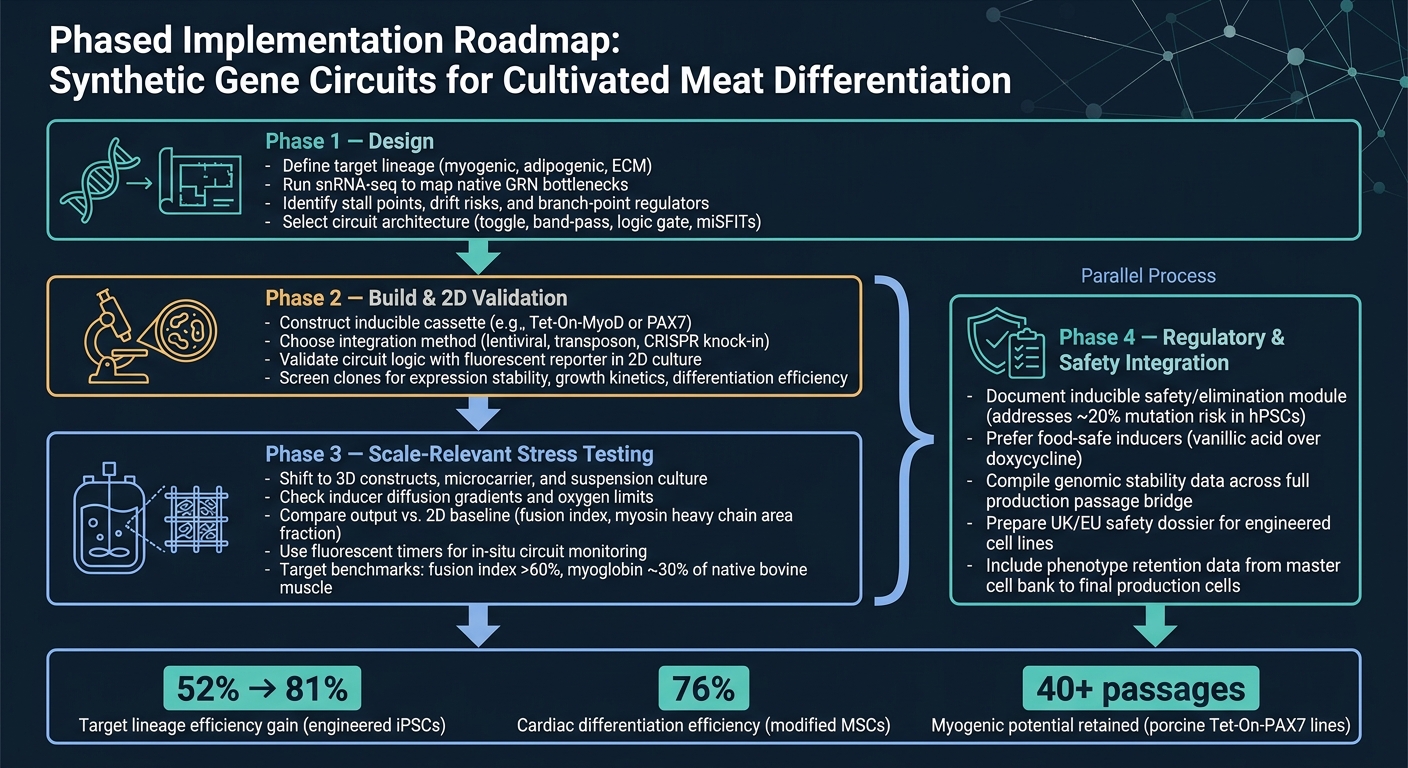
תהליך עבודה של מעגל גנים סינתטי להתמיינות בשר מתורבת
סיפורי מדענים: מייקל אלוויץ, מעגלים גנטיים בתאים חיים
השוואה מהירה
| נושא | מה שהמאמר אומר במילים פשוטות |
|---|---|
| צוואר הבקבוק העיקרי | הבדלה, לא התרחבות |
| למה מעגלים עוזרים | הם מוסיפים שליטה בסף וזיכרון מצב תא בתוך התא |
| הצעד הראשון הטוב ביותר | מפה שליטה בגורל טבעי עם snRNA-seq וכלים קשורים |
| אפשרויות המעגל העיקריות | מתג, הזנה קדימה, מעבר פס, שערי לוגיקה, miSFITs, CRISPRa/i |
| דוגמת מיוגנזה | Tet-On-MyoD להפרדת צמיחה מהתמיינות סופית |
| אדיפוגנזה / שליטת ECM | miSFITs ועיצובים בצורת פפיון לפלט מדורג מסוג PPARG/BMP4 |
| אתגר חיתוך שלם | שליטה ביחס בין שריר, שומן ורקמת חיבור |
| סיכון בהגדלה | התנהגות דו-ממדית עשויה לא להחזיק בשלושה ממדים או בביו-ריאקטורים |
| בחירות אינטגרציה | לנטיוירוס, טרנספוזונים, CRISPR knock-in, וקטורים אפיזומליים |
| נקודת רגולציה | קווים מהונדסים זקוקים לחבילת בטיחות רחבה יותר; משרים בטוחים למזון כמו חומצה ונילית מועדפים על פני DOX כאשר אפשר |
אז, במילים פשוטות: זה לא רק מאמר על עיצוב מעגלים. הייתי קורא את זה כמדריך לקישור ארכיטקטורת מעגלים, ביולוגיית שושלת, בחירת שיבוט, ביצועי ביוריאקטור ותיעוד בטיחות בבריטניה/האיחוד האירופי לאסטרטגיית הבחנה אחת.
המשך לקרוא אם אתה רוצה את המסלול המלא ממיפוי רשת מקומית לבחירת מבנה, בדיקות הגדלה והתאמה רגולטורית.
2. עקרונות עיצוב למעגלי שליטה בהבחנה
2.1 מיפוי רשת גורל התא המקומית לפני עיצוב המעגל
לפני שאתה מעצב מעגל, אתה צריך תמונה ברורה של מה שהתא כבר עושה.
ריצוף RNA של גרעין יחיד (snRNA-seq) הוא מקום טוב להתחיל בו. זה יכול להראות תת-אוכלוסיות שקטות, כולל תאי רזרבה המסומנים על ידי NOTCH2 ו-HEYL , ולהצביע על מטרות מסלול שעשויות לשפר הבחנה [3].
משם, ניתוח מסלול והסקת רשת רגולטורית גנטית (GRN) עוזרים למפות את סדר הפעלת הרגולטורים ולהדגיש היכן התאים נוטים להיתקע. במיוגנזה, המפל העיקרי עובר דרך MYOD1 ו-MYOG. באדיפוגנזה, הצמתים העיקריים הם PPARG ו-CEBPA, כאשר גורל פרוג'ניטור פיברו-אדיפוגני (FAP) פועל כנקודת סיכון עיקרית. הטבלה למטה מסכמת את הרגולטורים והצווארי בקבוק העיקריים.
| שושלת | רגולטורים ראשיים | מסלולי איתות קריטיים | צווארי בקבוק מזוהים |
|---|---|---|---|
| מיוגני | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | יצירת תאי רזרבה (מנוחה) |
| אדיפוגני | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | גורל תאי אב פיברו-אדיפוגניים (FAP) |
| פלוריפוטנטי | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | התמיינות ספונטנית / הטרוגניות |
שכבה שימושית נוספת היא פרופיל ביטוי miRNA. miRNAs אנדוגניים כמו miR-302a, הקשור לפלוריפוטנטיות, ו-miR-375, הקשור להתמיינות, יכולים לפעול כמסווגים פנימיים בעיצובים של חישה ותגובה. שמאפשר למעגל לקרוא את מצב התא האמיתי במקום להסתמך רק על גורמים חיצוניים [5].
הצווארי בקבוק הללו צריכים להנחות את בחירת המעגל. אם הבעיה העיקרית היא סחיפה, ייתכן שתצטרך נעילה. אם התזמון הוא הבעיה, פולס עשוי להתאים יותר. אם שליטה בגורל תלויה ביותר מרמז אחד, לוגיקה מרובת קלטים בדרך כלל הגיונית יותר.
2.2 בחר את ארכיטקטורת המעגל הנכונה
כאן מופיעים הפשרות בצורה ברורה. הארכיטקטורה הנכונה תלויה בשלוש נקודות מעשיות: כמה קבועה המחויבות צריכה להיות, כמה הדוק צריך להיות התזמון, וכמה מטען גנטי התא יכול לשאת ללא בעיות.
מתגים דו-יציבים מתאימים למקרים שבהם מחויבות השושלת צריכה להישאר נעולה. הבעיה העיקרית היא היפוך מצב ספונטני הנגרם על ידי רעש תעתוקי.
מסנני מעבר פס מתאימים למקרים שבהם גורם שעתוק נדרש רק בתוך חלון התפתחותי מוגדר. הבעיה היא שרמות המשרה צריכות להיות תחת שליטה הדוקה, אחרת התזמון מחליק.
שערים לוגיים מוסיפים ספציפיות על ידי דרישה ליותר מקלט אחד בו זמנית. לדוגמה, ניתן לאפשר התמיינות רק כאשר משרה חיצונית נוכחת והתא מראה את פרופיל ה-miRNA האנדוגני הנכון. זה עוזר להפחית את הסיכון להתחייבות לא מכוונת.
הטבלה למטה מתארת את הארכיטקטורות העיקריות ואת הפשרות הנלוות להן.
| ארכיטקטורה | הפיכות | דיוק זמני | מורכבות אינטגרציה | שימוש עיקרי | סיכון מרכזי |
|---|---|---|---|---|---|
| מתג דו-יציב | נמוך (לאחר נעילה) | בינוני | בינוני | מחויבות שושלת קבועה | היפוך ספונטני עקב רעש |
| מסנן מעבר פס | גבוה (תלוי בריכוז) | גבוה | גבוה | שלבי התפתחות חולפים | דורש שליטה הדוקה במשרה |
| שער לוגי (AND/OR/NOT) | משתנה | בינוני | בינוני–גבוה | הפעלה ספציפית לסוג תא | דליפה במצבי OFF |
| פפיון / קלט מרובה | גבוה | מתון | מתון | אינטגרציה של אותות מרובים | מבוסס על יציבות miRNA אנדוגני |
| miSFITs | גבוה | מתון | נמוך–מתון | כוונון פלט מדורג | טווח דינמי צר אם לא מכוון היטב |
"על ידי מזעור מספר השכבות החישוביות תוך שמירה על פונקציונליות, אסטרטגיה זו מתמודדת עם מחסומי מדרגיות בהנדסת מעגלי גנים." - Nature Communications [9]
כל שכבת רגולציה נוספת מגדילה את העומס ומשתמשת במשאבים תאיים. בפועל, אם שני עיצובים מבצעים את אותה עבודה, הפשוט יותר הוא בדרך כלל ההימור הטוב יותר כאשר הגודל חשוב.
לאחר שהארכיטקטורה נקבעת, המשימה הבאה היא לוודא שהיא תחזיק מעמד תחת דליפה נמוכה, דיכוי רעש ובקרה בטוחה.
2.3 בנייה לאמינות, דליפה נמוכה ובטיחות
מעגל חייב להישאר יציב לאורך תרבות ממושכת. פרץ קצר של ביצועים טובים אינו מספיק לשימוש בייצור.
חלקים אורתוגונליים הם ההגנה הראשונה. פרומוטורים, גורמי שעתוק ואלמנטים רגולטוריים שאינם מגיבים עם המערכת הטבעית עוזרים להגביל השפעות לא מכוונות ולהפחית את הסיכוי שאותות אנדוגניים יפעילו את המעגל או יכבו אותו.מקדמים בעלי אטימות גבוהה מותאמים כמו PCREm שימשו להפחתת דליפה בסיסית במערכות יונקים מושרות [6].
רגולציה עצמית שלילית גם היא ראויה להוספה כאשר אפשרי. זהו אחד המוטיבים הידועים יותר להפחתת רעש תעתוקי וליצירת תגובה לינארית יותר לריכוז המשרה [6]. לולאות הזנה קדמית לא עקביות (iFFLs) יכולות להוסיף שכבת בקרה נוספת על ידי סינון תנודות סטוכסטיות, כך שהתאים יגיבו לאותות מתמשכים ולא לפולסים קצרים של רעש.
גרסאות עם קודונים מותאמים (cm ) של גורמי תעתוק סינתטיים מקלות על האפיון גם כן. הן מאפשרות להפריד בין ביטוי מונע מעגל לבין ביטוי גנומי אנדוגני (g) במהלך האימות [1]. זה אולי נשמע כמו פרט קטן, אבל זה חוסך זמן כשאתם מנסים להבין אם קריאה מגיעה מהמעגל או מהגנום המארח.
נדרשים מודולי בטיחות. כ-20% מתאי גזע פלוריפוטנטיים אנושיים נושאים מוטציות הקשורות לסרטן [7]. לכן, אם מעגל נכנס לקו שמקורו בתאי גזע, עליו לכלול מודול עצירת צמיחה או חיסול שניתן להפעיל. חומצה ונילית היא משרה שימושית שיש להעדיף כאן מכיוון שהיא תוסף מזון מורשה, מה שעוזר במקרה של שימוש בה כטריגר למעגל בקווי תאים של בשר מתורבת [1].
"ביולוגיה סינתטית מספקת למהנדסים את האמצעים לפרוס מעגלים כדי לכוון בקלות ובדייקנות את הביטוי של גנים מרובים כדי... לחסל תופעות לוואי פוטנציאליות לא רצויות." - npj Systems Biology and Applications [6]
בחירות אלו מגדירות את המעגלים הספציפיים לשושלת בסעיף 3.
sbb-itb-ffee270
3. אסטרטגיות מעגלים להתמיינות מיוגנית, אדיפוגנית ובקרת יחס
3.1 מעגלים מיוגניים שמפרידים בין התרחבות להתמיינות סופית
לאחר שהארכיטקטורה של המעגל נקבעת, המשימה הבאה היא פריסה ספציפית לשושלת. עבור מיוגנזה, הבעיה העיקרית פשוטה להצהרה אך קשה לביצוע: תאים צריכים להתרבות תחילה, ואז לעבור להתמיינות כאשר מתבקשים, מבלי לסטות מוקדם מדי לכיוון כלשהו.
קסטה Tet-On-MyoD היא אחת הדרכים הישירות ביותר לעשות זאת. בהגדרה זו, תאים מתרבים בתנאים סטנדרטיים כאשר דוקסיציקלין (DOX) נעדר. הוסף DOX, והמעגל מניע התחייבות מיוגנית.חוקרים ממספר מוסדות סיניים השתמשו בגישה זו בפיברובלסטים עובריים של תרנגולות ודיווחו על היווצרות מיוטוב יעילה לאחר אינדוקציה [4].
מתג דו-יציב מציע שליטה הדוקה יותר במצב. מערכות שנבנו ממדכאים מעכבים הדדית כמו E-KRAB ו-Pip-KRAB יכולות להפוך פולס DOX קצר לתוכנית מיוגנית יציבה [6]. בפועל, המשמעות היא שהמצב המובדל נשאר נעול לאחר אינדוקציה במקום לדעוך לאחר שהאות מוסר. הוספת אוטורגולציה שלילית גם עוזרת להפחית רעש סטוכסטי ולהגביל התמיינות דולפת במהלך שלב ההתרחבות [6].
זה חשוב מכיוון שהתמיינות מיוגנית סטנדרטית במבחנה לעיתים קרובות נעצרת באמצע הדרך. מדדי היתוך הם בדרך כלל רק סביב 50–60%, מה שמשאיר אוכלוסיית רזרבה גדולה שאינה מתמזגת [3]. הפעלה מונעת מעגל של MyoD יכולה לשפר את המחויבות, אך מחויבות לבדה אינה מבטיחה איחוי אחיד. כאשר מעגלי MyoD משולבים עם מודולציה של MEK, NOTCH ו-RXR, איחוי כמעט מלא בתרבות דו-ממדית הופך לאפשרי [3]. המעגל שולט בתזמון הכניסה לשושלת; המולקולות הקטנות דוחפות יציאה נקייה ואחידה יותר ממחזור התא.
3.2 שליטה אדיפוגנית ופיברוגנית עבור הרכב ומבנה
שריר לבדו אינו מספיק. ברגע שהשליטה המיוגנית קיימת, הבעיה הבאה היא הרכב: כמה שומן נוצר, כמה ECM מופקד ומתי התוכניות הללו מופעלות. כאן, שליטה בינארית ON/OFF היא לעיתים קרובות גסה מדי. מה שצוותים בדרך כלל צריכים הוא פלט מדורג, במיוחד סביב נקודת הפיצול האדיפוגנית-פיברוגנית.
miSFITs מספקים דרך מעשית לכוון ביטוי בשלבים.על ידי הצבת אתרי מטרה miRNA מוטנטים - לדוגמה, אתרים עבור miR-17 - ב-3′UTR של גנים כמו PPARG או BMP4, חוקרים יכולים לבחור רמות ביטוי מספריית וריאנטים [5]. זה הופך את האינדוקציה של תאי שומן ליותר כמו מתג דימר מאשר מתג אור. במקום לדחוף תאים לתגובה של הכל או כלום, צוותים יכולים לכוון את האדיפוגנזה בזהירות רבה יותר [5].
פיברובלסטים הם גם לא רק צופים כאן. הם מספקים חלבוני ECM שמעצבים את המרקם [10]. זה הופך את השליטה הפיברוגנית לחלק מעיצוב המוצר, ולא רק נושא צדדי. מעגלים יכולים לעזור לנהל את המעבר בין מצבים פיברוגניים ואדיפוגניים, ובפולטרי מתורבת, ייתכן שיהיה צורך בהפעלה ישירה של PPARG בפיברובלסטים כדי ליצור הפקדת שומן משמעותית [10].
ארכיטקטורת פפיון מתאימה לבעיה זו מכיוון שהיא שומרת על חישה ופלט בנפרד. שכבת החישה קוראת את מצב התא הנוכחי, בעוד ששכבת הפלט מתאימה את PPARG, CEBPA או רגולטורים אחרים של שושלת. ההפרדה הזו עוזרת למנוע מתוכניות אדיפוגניות או של רקמת חיבור להידלק לפני שהתאים הגיעו לשלב ההתפתחותי הנכון.
3.3 שליטה ביחס רב-שושלתי וחישת משוב
מעגלי שליטה ביחס מתמודדים עם בעיה שונה: לא האם מתרחשת התמיינות, אלא האם תערובת האוכלוסייה הסופית נשארת במקום שבו היא צריכה להיות. עבור מוצרים של חיתוך שלם, הכנסת שריר, שומן ו-ECM לפרופורציות הנכונות חשובה בדיוק כמו לגרום לכל שושלת להתמיין.
מערכות אלו בונות בקרת משוב בתוך התאים עצמם. מקדמים ספציפיים למצב מגבילים את ביטוי חלבוני האותות לתאים שכבר התחייבו לשושלת נתונה.מודולים פאראקריניים סינתטיים מאפשרים אז לתאים מיוגניים מחויבים לשחרר אות מעכב שמדכא התחייבות אדיפוגנית בתאים סמוכים. הלוגיקה דומה לעיכוב לטרלי במערכות התפתחות Delta-Notch [1][6] . כאשר הסתעפות הופכת למורכבת יותר, שערים מרובי קלט יכולים לשלב רמזים חוץ-תאיים עם אותות מצב פנימיים [9].
miSFITs פועלים גם בשלב זה. על ידי כיוון עוצמת הפלט של BMP4 או מורפוגנים אחרים, צוותים יכולים לשנות את איזון השושלת מבלי לשכתב את לוגיקת ההחלטה במעלה הזרם. ברשתות רגולציה גנטיות מהונדסות, סוג זה של שליטה הגדיל את יעילות ההתמיינות של שושלת היעד מ-52% בביקורות ל-81% ב-iPSCs מהונדסים [2]. בתאים מזנכימליים שעברו שינוי, מעגלים סינתטיים הניעו את יעילות ההתמיינות הקרדיאלית ל-76% [2].
הטבלה למטה משווה בין גישות של שושלת יחידה ושליטה ביחס תוך שימוש בנקודות החשובות ביותר בייצור.
| תכונה | מעגל חד-שושלתי | מעגל בקרת יחס |
|---|---|---|
| מורכבות | נמוכה; בדרך כלל פרומוטר משרה יחיד ורגולטור [4] | גבוהה; דורש שערי לוגיקה וסיגנלינג פאראקריני [6][9] |
| עומס ניטור | נמוך; בדרך כלל עוקב אחר מדווח יחיד [4] | גבוה; דורש מעקב אחר מספר סמני שושלת [5] |
| חוסן | בינוני; נוטה להטרוגניות ולהיווצרות תאי רזרבה [3] | גבוה; משתמש במשוב ועיכוב לטרלי לאיזון אוכלוסיות [1] |
| ערך ייצור | גבוה עבור ביומסה; מוגבל עבור מבנה רקמות מורכב [10] | חיוני למוצרים שלמים שדורשים שריר, שומן ו-ECM [4] |
מעגלי שליטה ביחס מוסיפים עומס אימות כבד יותר.אבל המשוב המובנה שלהם קשה להתאים עם בקרת תהליך בלבד, מה שמפעיל יותר לחץ על בחירת שיבוט ובדיקת תהליך.
4. מהמבנה לתהליך: אימות, הגדלה והתאמה רגולטורית
4.1 אסטרטגיית אינטגרציה ובחירת שיבוט לביצועים יציבים
לאחר תכנון המעגל, החלק הקשה מתחיל: הכנסת המעגל לתאים בצורה שנשארת יציבה דרך מעבר בקנה מידה ייצור.
העברה באמצעות וירוס לנטי היא לעיתים קרובות יעילה ומספקת אינטגרנטים יציבים במהירות. אבל האינטגרציה היא אקראית. זה אומר יותר תשומת לב רגולטורית, בנוסף לסיכון שהביטוי יירד עם הזמן בגלל השתקה. מערכות טרנספוזון כמו PiggyBac ו-Sleeping Beauty נמצאות באמצע. הן יכולות לשמור על ביצועים לאורך מעברים רבים, אבל עדיין צריך לסנן עבור מספר עותקים ואתרי הכנסה. היפהפייה הנרדמת, לדוגמה, שימשה להאלמות יציבה של תאי לוויין בקר על ידי ביטוי יתר של TERT ו-CDK4, עם קווים ששומרים על פוטנציאל מיוגני מעבר ל-40 מעברים [10] . הכנסה באמצעות CRISPR נותנת את השליטה ההדוקה ביותר על מיקום הנחיתה של המבנה וההגדרה הגנומית המדויקת ביותר, אם כי בחירת השיבוטים איטית יותר והתפוקה נמוכה יותר.
| שיטת אינטגרציה | בקרת החדרה | יציבות | יכולת הרחבה | שיקולים רגולטוריים |
|---|---|---|---|---|
| העברה באמצעות וירוס לנטי | נמוך (אינטגרציה אקראית) | גבוה, אך נוטה להשתקה | גבוה | פיקוח רגולטורי מוגבר בשל החדרה אקראית ושאריות ויראליות |
| טרנספוזונים (PiggyBac/SB) | בינוני | גבוה לאורך מעברים רבים | גבוה | דורש סינון למספר עותקים ואתרי החדרה |
| CRISPR Knock-in | גבוה (ממוקד לאתר ספציפי) | גבוה מאוד | בינוני | מועדף; מפחית את הסיכון להפרעה בגנים אנדוגניים |
| וקטורים אפיזומליים | ללא (חוץ כרומוזומלי) | נמוך; יכול ללכת לאיבוד במהלך חלוקה | נמוך | מחסום אינטגרציה נמוך יותר, אך לא מתאים להתרחבות לטווח ארוך |
סינון שיבוטים צריך לעשות יותר מאשר לאשר שהמבנה קיים. זה צריך לעקוב אחר נדידת ביטוי, פרופיל החדרה, קינטיקה של צמיחה, יעילות התמיינות ושימור פנוטיפ במספר מעבר גבוה. snRNA-seq שימושי כאן כי הוא יכול לשלול שיבוטים מועשרים בתאי Pax7⁺/Ki-67⁻ רזרבה - תאים שיוצאים ממחזור התא מבלי להתמיין - לפני הגדלה [3]. תאי EPSC חזיריים עם מעגל Tet-On-PAX7 שמרו על התמיינות שריר גבוהה בתרבות מיקרונשאים ותליה תלת-ממדית מעל 40 מעברים [8].
4.2 כיצד משתנה התנהגות המעגל בתרבות תלת-ממדית, מיקרונשאים וביוראקטור
ברגע שיש לך שיבוט, המבחן הבא הוא האם הוא מתנהג באותו אופן מחוץ ל-2D. במקרים רבים, זה לא. ביצועים ב-2D לעיתים רחוקות עוברים בצורה נקייה לתליה, מיקרונשאים או תרבות פיגום כי גרדיאנטים של דיפוזיה, מגבלות חמצן וגזירה כולם משנים את תפוקת המעגל.
אחת הבדיקות הראשונות היא דיפוזיה של משרה. בביו-ריאקטורים עם מיכל מעורב, משרות מולקולות קטנות צריכות להגיע לתאים באופן אחיד. בפועל, יכולים להיווצר גרדיאנטים, במיוחד בתרביות מיקרוקרייר צפופות ובתוך אגרגטים או ליבות שלד. תרבית השעיה בדרך כלל מתאימה יותר לבשר מתורבת בקנה מידה גדול מכיוון שהיא תומכת בצפיפות תאים גבוהה יותר ומספקת שליטה הדוקה יותר בתהליך.
מעקב אחר מצב התאים גם הופך לקשה יותר ככל שהמערכת מתרחבת. אותות דיווח פלואורסצנטיים שקל לקרוא במיקרוסקופיה ב-2D יכולים להיות קשים לפענוח במבנים אטומים ב-3D. טיימרים פלואורסצנטיים - גששים שמשנים פליטה מכחול לאדום כשהחלבון מתבגר - יכולים לספק נתוני הפעלת מעגל בזמן אמת במקום [1]. המסלול ההגיוני הוא אימות בשלבים: תחילה ב-2D, לאחר מכן במבנים תלת-ממדיים, ולבסוף בתנאי הביוראקטור הסופיים [3][8] .
4.3 אפיון, תיעוד בטיחות מזון ושיקולים בבריטניה/האיחוד האירופי
לאחר בדיקות התהליך, האפיון צריך להראות כי פונקציית המעגל, הפנוטיפ והבטיחות עדיין מתקיימים. ערכת הנתונים המרכזית צריכה לכלול ציטומטריה של זרימה, qPCR עם רצפי cm, קורסי זמן של RNA-seq, ותוצאות פונקציונליות כמו חלק שטח שרשרת כבדה של מיוזין וביטוי מיוגלובין [1]. הוכח כי מדיה אופטימלית להבדלה ללא סרום מביאה את ביטוי המיוגלובין לכ-30% מהרמות שנמצאו בשריר בקר טבעי [3]. זה נותן לצוותים מדד ברור במקום יעד מעורפל.
אתה גם צריך לתעד פרופילים של חלבון, חומצות אמינו ושומנים, יחד עם תכונות חושיות [10][3].
מבחינה רגולטורית, בריטניה והאיחוד האירופי משרטטים קו ברור בין קווי תאים שהפכו לאלמותיים באופן ספונטני (לא מהונדסים גנטית) לבין קווים מהונדסים גנטית. האחרונים זקוקים לתיקים בטיחותיים רחבים יותר [10][3]. חבילות יציבות צריכות להראות שמירה על פנוטיפ ויציבות גנומית לאורך כל גשר הייצור - מבנק התאים הראשי ועד לתאי הייצור הסופיים - ורשומות המעקב צריכות לכלול כל מעבר ביניהם [10]. אם המעגל תלוי במפעיל כימי, תוסף בטוח למזון או מורשה כמו חומצה ונילית עדיף על דוקסיציקלין [1].
מעקב גנומי שגרתי הוא הכרחי, ויש לתעד מתג התאבדות או חיסול מושרה כאמצעי שליטה בסיכון מרכזי [7]. יש לכלול את תפקודו גם בתיק הבטיחות, במיוחד כאשר כללי בריטניה והאיחוד האירופי לבשר מתורבת ממשיכים להתגבש.
5. מפת דרכים מעשית ומסקנה
5.1 מפת דרכים ליישום מדורג לצוותי בשר מתורבת
הדרך הנקייה ביותר מהקונספט לייצור היא תהליך עבודה בשלבים.
שלב 1 הוא תכנון. התחילו בהגדרת השושלת המטרה, ואז השתמשו ב-snRNA-seq כדי לאשר את צווארי הבקבוק העיקריים לפני שתבחרו ארכיטקטורת מעגל. שלב זה חשוב כי מעגל יכול לפתור רק את המגבלות שזיהיתם בפועל.
שלב 2 הוא בנייה ואימות ב-2D. בנו את המבנה ובדקו שהמעגל מתנהג כמתוכנן ב-2D, תוך שימוש בקריאת דיווח פשוטה.בשלב זה, המטרה היא פשוטה: לאשר שהלוגיקה עובדת לפני המעבר לדגמים קשים ויקרים יותר.
שלב 3 הוא בדיקות מאמץ בקנה מידה רלוונטי. מעבר למערכות תלת-ממד ותנאים רלוונטיים לביוראקטור, ואז השוואת התפוקה מול הבסיס הדו-ממדי. זה המקום שבו עיצובים רבים מתחילים להראות את נקודות התורפה שלהם, במיוחד כאשר העברת מסה, גזירה והשפעות מטריצה נכנסות לפעולה.
שלב 4 הוא שילוב רגולטורי ובטיחותי, והוא צריך להתבצע במקביל לשלב 3. עבודת הבטיחות והרגולציה לא צריכה להמתין עד הסוף. יש להריץ אותה לצד ההגדלה, כולל תיעוד עבור כל מודול בטיחות ניתן להשראה.
5.2 השגת כלים וחומרים מאפשרים דרך Cellbase
ברגע שהזרימה נקבעת, ההשגה לעיתים קרובות הופכת לשלב המגביל את הקצב.
- קווי תאים
- מדיה ללא סרום ומוגדרת כימית
- פיגומים
- רכיבי ביוריאקטור
- חיישנים
- ציוד אנליטי
גישה אמינה לחומרים תואמים בכל שלב משפיעה ישירות על מהירות האפיון של התנהגות המעגל בתנאים רלוונטיים להיקף.
5.3 נקודות מפתח
מעגלי גנים סינתטיים מעניקים לצוותי בשר מתורבת שליטה ניתנת לתכנות על תזמון, ספים ואיזון שושלת שלא ניתן להשיג בפרוטוקולים מבוססי מדיה בלבד. בחירת הארכיטקטורה מעצבת הפיכות, דליפות ובטיחות.מערכות הניתנות להשראה בדרך כלל מועדפות מכיוון שהן מספקות שליטה מותנית ועומס מטבולי נמוך [6].
"ערכת הכלים של הביולוגיה הסינתטית יכולה לשמש להקמת קווי תאים עם ביטוי גנים הניתן לכוונון, אשר, כאשר משולבים עם PAT ודוגמנות חישובית, יכולים לאפשר מערכות בקרה בלולאה סגורה לספק תפוקה ואיכות מוצר אופטימליים." - npj Systems Biology and Applications [6]
הפריסה המוצלחת אינה בעיה ביולוגית בלבד. היא תלויה בקישור הדוק בין הנדסת מעגלים, תכנון תהליכים ביולוגיים, תיעוד רגולטורי ורכש.
שאלות נפוצות
כיצד מעגלי גנים סינתטיים משפרים את עקביות ההבחנה?
מעגלי גנים סינתטיים יכולים להפוך את ההבחנה לעקבית יותר מכיוון שהם נותנים לך שליטה מתוכנתת על התנהגות התאים והתחייבות השושלת.בפועל, זה אומר שימוש בפעולות לוגיות מודולריות כדי לכוון את הביטוי של גנים וגורמי שעתוק עם תזמון מדויק.
התזמון הזה חשוב. הוא עוזר לתאים לעבור דרך שינויים במצב מוגדרים בסדר הנכון, במקום להיסחף למצבים מעורבים או לא רצויים. זה גם מפחית התמיינות לא מכוונת ומפחית רעש בתרבות.
התוצאה היא פשוטה: אוכלוסיות תאים אחידות, יציבות ובוגרות יותר לייצור בשר מתורבת.
איזה עיצוב מעגל מתאים לשליטה מיוגנית או אדיפוגנית?
במחקר בשר מתורבת, אותם פיברובלסטים של עוף יכולים להיות מונעים לכל אחד מהקווים. מיוגנזה עוקבת אחר סט אחד של פרוטוקולי השראה, בעוד ש-אדיפוגנזה יכולה להיות מופעלת על ידי חשיפת התאים לקלטים כמו סרום עוף או חומצות שומן.
משם, ניתן לשלוט בגורלות התאים הללו שלב אחר שלב בתוך פיגומי הידרוג'ל תלת-ממדיים כדי לבנות מבני בשר עם יחסי שומן וקולגן מוגדרים.
מדוע מעגלי גנים מתנהגים לעיתים קרובות בצורה שונה בתרבות תלת-ממדית?
ב-תרבות תלת-ממדית , מעגלי גנים מתנהגים לעיתים קרובות בצורה שונה מכיוון שהתאים מתמודדים עם קלטים פיזיים ומבניים שפשוט לא קיימים בשכבות דו-ממדיות. קלטים אלו כוללים מתח מכני, לחץ גזירה, קשיחות המטריצה וצפיפות תאים מקומית.
רמזים אלו יכולים לשנות מסלולי איתות כמו Notch. הם יכולים גם לשנות את האופן שבו מעגלים סינתטיים מזהים כוח ומתאמים תגובות במורד הזרם, כולל הידבקות תא-תא ומורפוגנזה של רקמות.









