

CRISPR는 산업용 생물반응기에서 세포 스트레스를 해결하여 배양육 생산을 혁신하고 있습니다. 이 도구는 가혹한 조건에서 세포 생존을 개선하고 증식을 연장하며 노화를 줄이기 위해 정밀한 유전자 편집을 가능하게 합니다. 예를 들어, TP53 및 PTEN과 같은 유전자를 제거하면 일차 대 불멸화 세포주 배양 기간이 100일에서 200일로 연장되고 30일 만에 세포 풍부도가 1,000배 증가했습니다. 그러나 이러한 수정은 분화에 영향을 미칠 수 있으므로 신중한 최적화가 필요합니다.
기사의 주요 통찰은 다음과 같습니다:
- 생물반응기에서의 스트레스 요인: 전단력, 영양 불균형 및 산화 스트레스는 세포 생존력을 감소시킵니다.
- CRISPR 전략: 유전자 제거 (TP53, PTEN) 및 활성화 (HIF1A)는 특정 스트레스 반응을 목표로 합니다.
- 검증: 편집된 세포는 성능과 분화 잠재력을 보장하기 위해 유전체, 단백질체 및 기능적 테스트를 거칩니다.
-
확대: 바이오리액터 조건으로의 전환은 최적화된 배지와 장비를 포함하며,
Cellbase 와 같은 플랫폼은 맞춤형 자원을 제공합니다.
CRISPR의 정밀성은 스트레스 저항성 세포주 개발을 가능하게 하지만, 성장과 분화의 균형을 맞추는 것은 확장 가능한 배양육 생산에 여전히 중요합니다.
유전적 설계를 위한 바이오리액터 스트레스 프로파일 매핑
주요 바이오리액터 스트레스 요인 식별
CRISPR 편집을 시작하기 전에 유전적 설계를 안내하기 위해 바이오리액터 스트레스 프로파일을 매핑하는 것이 필수적입니다. 바이오리액터의 스트레스 요인은 특정 세포 반응을 유발하며, 적절한 유전적 목표를 선택하기 위해 잘 이해되어야 합니다.
기계적 및 유체역학적 스트레스는 가장 즉각적인 과제 중 하나입니다. 교반 탱크 생물반응기는 세포막을 손상시키고 세포 신호 경로를 방해할 수 있는 전단력을 생성합니다 [5][2]. 영양 및 대사 스트레스도 주요 역할을 하며, 종종 불균형한 영양소 흡수에서 비롯됩니다. 3D 스캐폴드에서의 영양소 구배와 암모니아 축적은 대사적 부담을 가중시킵니다 [3][5][6]. 또한, pH 변동과 높은 온도는 세포 증식 속도를 감소시키고 심지어 세포를 조기 분화로 밀어 넣을 수 있습니다 [3][2].
산화, 미토콘드리아 및 ER 스트레스를 포함한 기타 스트레스 요인은 세포 생존 가능성을 더욱 어렵게 만듭니다.산화 스트레스는 천연 항산화제가 없기 때문에 세포가 활성 산소종에 더 취약해져서 무혈청 배지로 전환할 때 특히 심각해집니다 [4]. 세포 수준에서, 미토콘드리아 스트레스와 소포체(ER) 스트레스 는 생물공정 조건이 최적 범위에서 벗어날 때 발생합니다 [6]. UCSF의 신경퇴행성 질환 연구소의 Xiaoyan Guo는 이 동태를 강조합니다:
"다양한 생리적 및 환경적 스트레스가 존재할 때, 세포는 세포 항상성을 재확립하기 위해 신속하게 스트레스 반응을 시작합니다." [6]
이러한 스트레스 요인을 사전에 매핑함으로써, 문제가 발생할 때 반응하는 대신, 연구자들은 정확한 유전공학 목표를 정의할 수 있습니다.이 체계적인 접근 방식은 CRISPR 전략이 스트레스에 강한 세포주 개발을 효과적으로 목표로 하도록 보장합니다.
오믹스 데이터를 사용하여 스트레스 반응 유전자를 찾기
스트레스 환경을 특성화한 후, 다음 단계는 이러한 조건에 반응하는 유전자를 식별하는 것입니다. 전사체학 (RNA-seq) 및 단백질체학과 같은 도구는 세포가 건강한 초기 배양 상태에서 스트레스가 많은 후기 배양 조건으로 이동함에 따라 유전자 발현 및 단백질 풍부도의 변화를 추적하는 데 매우 유용합니다 [1] [6]. 그러나 이러한 방법은 하위 효과를 포착하는 반면, 이러한 변화를 유도하는 상위 조절자를 식별하는 데 종종 실패합니다 [6].
풀드 CRISPR 녹아웃 스크린은 이 격차를 메웁니다.수천 개의 유전자를 대규모 세포 집단에서 체계적으로 교란함으로써, 이러한 스크린은 스트레스 하에서 성장 이점을 제공하는 유전자 교란을 밝혀내어 중요한 조절 허브를 발견합니다 [1] [6]. 예를 들어, TP53 및 PTEN과 같은 유전자를 표적으로 삼는 것은 장기간의 배양 스트레스로 인한 분자 노화 서명을 역전시키는 것으로 나타났습니다. 이는 후기 통과 세포가 초기 통과 야생형 세포와 유사한 전사체 프로파일을 유지할 수 있게 합니다 [1] .
계층적 군집화, 를 사용하여 연구자들은 시간이 지남에 따라 발현 변화에 따라 유전자를 그룹화하여 세포 주기 진행 및 단백질 합성과 같은 과정과 관련된 모듈을 분리할 수 있습니다. 이러한 과정은 일반적으로 생물반응기 유도 노화가 진행됨에 따라 감소합니다 [1]. When combined with 경로 강화 분석 (via tools like gprofiler2), these modules can be linked to specific biological pathways, such as TGFβ signalling or chondrogenic differentiation, which may actively limit cell expansion [1].
아래 표는 포괄적인 스트레스 맵을 구성하는 각 방법의 기여도를 설명합니다:
| 방법 | 주요 사용 | 주요 출력 |
|---|---|---|
| 전사체학 (RNA-seq) | mRNA 발현 변화 측정 | 스트레스 받은 세포와 받지 않은 세포 간의 차등 발현 유전자 (DEGs) [1] |
| 프로테오믹스 | 단백질 풍부도 측정 | 특정 스트레스 요인에 매핑된 번역 출력 [6] |
| 통합 CRISPR 스크린 | 기능적 유전자 교란 | 상위 조절 허브 및 적합성-중요 유전자 [1][6] |
| PCA & 계층적 군집화 | 데이터 시각화 및 그룹화 | 세포 상태 변화 및 공동 조절 스트레스 반응 경로[1] |
sbb-itb-ffee270
세포주 공학 CRISPR-Cas9 - 성공을 극대화하기 위한 팁과 요령
스트레스 저항성 세포주 공학을 위한 CRISPR 전략
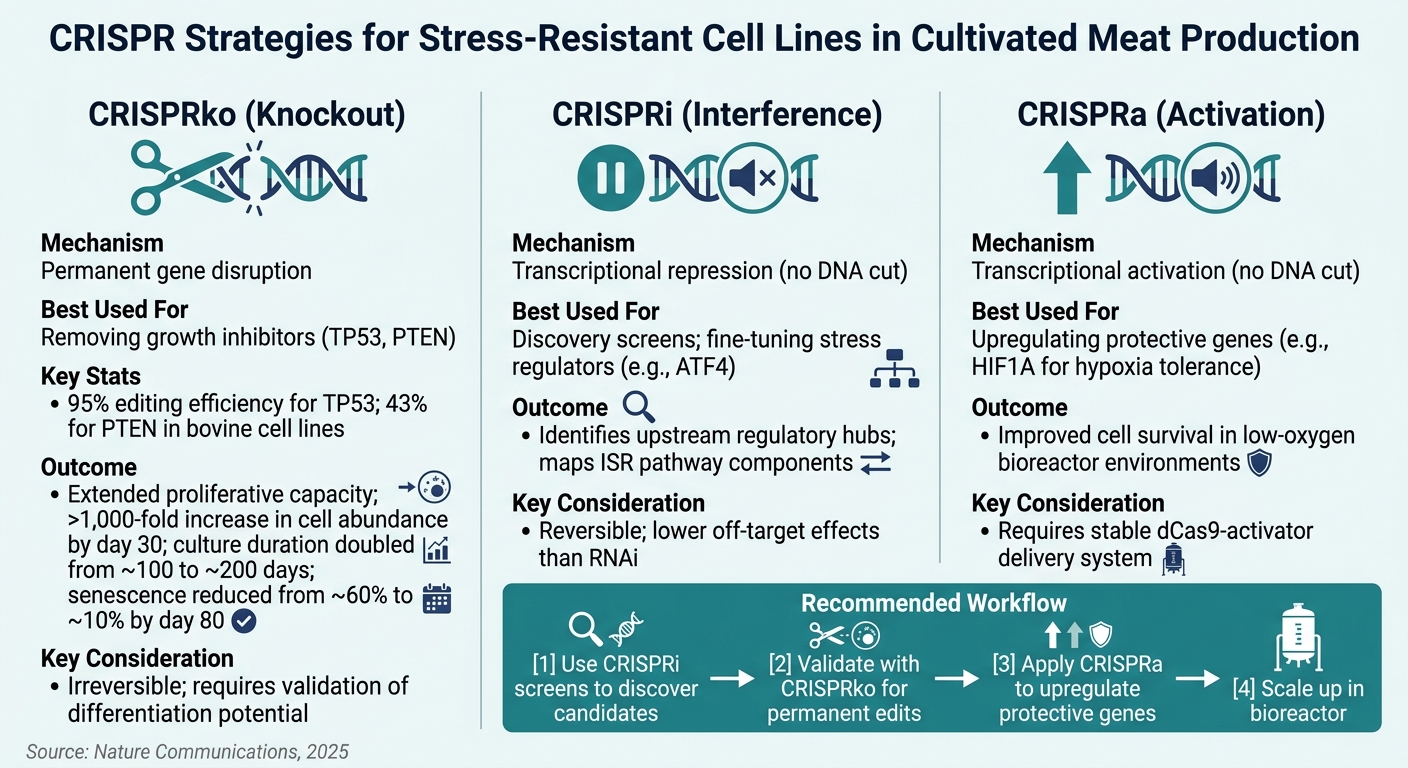
배양육에서 스트레스 저항성 세포주를 위한 CRISPR 기술
스트레스 저항성을 위한 주요 유전자 및 경로
상세한 스트레스 지도를 손에 들고, 다음 단계는 편집할 표적 유전자를 정확히 찾아내는 것입니다.표적 선택은 세포 성능에 영향을 미치는 주요 스트레스 요인에 따라 달라집니다.
복제 노화는 세포 증식을 제한하기 때문에 배양육 생산에서 주요 장애물입니다. 이 분야의 세포 소스 중 약 25%는 중간엽 줄기 세포(MSC)이며, 반복적인 계대 배양 후에는 비가역적인 성장 정지에 직면합니다 [1]. p53 종양 억제 단백질을 암호화하는 유전자인 TP53을 제거하면 이 문제를 직접적으로 해결할 수 있습니다. 소 MSC에 대한 연구에 따르면 TP53 제거는 세포의 증식 능력을 크게 확장하여 편집되지 않은 세포주보다 훨씬 더 많이 분열할 수 있게 합니다 [1]. 마찬가지로, PTEN을 제거하면 PI3K/AKT/mTOR 경로가 강화되어 스트레스 저항성이 향상됩니다 [1] .
대사 및 미토콘드리아 스트레스를 해결하기 위해 통합 스트레스 반응 (ISR)은 중요한 경로입니다. 전사 인자 ATF4는 미토콘드리아 스트레스 반응을 조정하는 데 중심적인 역할을 하며, CRISPR 스크린은 상위 조절자를 매핑하는 데 중요한 역할을 했습니다 [6] . 캘리포니아 대학교 샌프란시스코의 Xiaoyan Guo와 Martin Kampmann은 다음과 같이 설명합니다:
"전사 또는 번역 리포터를 기반으로 한 편향 없는 유전적 스크린은 특정 스트레스 반응의 조절 인자를 식별하는 강력한 접근 방식입니다." [6]
TGFβ 경로 또한 주목할 만하며, 특히 소 MSC를 확장하는 데 중요합니다. CRISPR 스크린은 TGFβ에 의해 유도된 연골 형성이 세포 증식을 억제한다는 것을 보여주었습니다.이 경로를 억제하면 세포를 미분화되고 확장 가능한 상태로 유지하는 데 도움이 됩니다 [1]. 3D 스캐폴드의 밀집된 코어에서 종종 발견되는 저산소 조건에서는 CRISPRa를 사용하여 , HIF1A를 활성화하면 저산소 환경에서 세포 생존이 개선됩니다. 이러한 수정은 산업 규모의 생물 반응기의 동적 조건에서 세포가 번성할 수 있도록 합니다.
그러나 세포 증식을 극대화하는 편집 - 예를 들어 TP53 노크아웃 - 은 세포가 근육이나 지방 조직으로 분화할 수 있는 능력을 감소시킬 수 있다는 점을 주의해야 합니다. 성장과 분화 잠재력 간의 이러한 균형은 엔지니어링 전략을 설계할 때 신중하게 조정되어야 합니다 [1].
| 스트레스 요인 | 주요 유전자 표적 | CRISPR 전략 | 결과 |
|---|---|---|---|
| 복제 노화 | TP53 | 노크아웃 | 증가된 증식 능력; 세포 풍부도 증가 |
| 영양/성장 스트레스 | PTEN | 노크아웃 | PI3K/AKT/mTOR 신호 강화; 생존율 향상 |
| 미토콘드리아 스트레스 | ATF4 | CRISPRi / 리포터 | 상위 조절 경로 식별 |
| 저산소증 | HIF1A | CRISPRa (활성화) | 저산소 바이오리액터 환경에서 생존율 증가 |
| 연골 형성 이동 | TGFβ 경로 | 노크아웃 / 억제 | 소 MSC에서 미분화, 증식 상태의 유지 |
핵심 유전자가 식별되면, 적절한 CRISPR 기술을 선택하는 것이 다음 중요한 단계가 됩니다.
CRISPR 편집 기술 비교
CRISPR 방법의 선택은 유전적 수정의 정밀성과 영구성을 결정합니다. 각 접근 방식은 영구적인 변화, 가역적인 조정 또는 탐색적 스크리닝이 목표인지에 따라 고유한 강점을 가지고 있습니다.
CRISPR 노크아웃 (CRISPRko)는 유전자를 영구적으로 비활성화하는 데 가장 적합한 방법입니다. TP53 및 PTEN, 과 같은 완전한 기능 상실이 필요한 표적에 이상적입니다. 검증 연구에 따르면 CRISPRko는 소 세포주에서 TP53에 대해 95%의 편집 효율성과 PTEN에 대해 43%의 효율성을 달성했습니다 [1]. 이러한 변동은 대규모 편집을 진행하기 전에 표적 특이적 효율성을 테스트하는 것이 중요함을 강조합니다.
CRISPR 간섭 (CRISPRi) 는 가역적인 유전자 억제를 제공하여 발견 단계에 이상적입니다.또한 RNAi에 비해 비표적 효과를 줄입니다 [6]. 반면에, CRISPR 활성화 (CRISPRa)는 저산소 내성 (HIF1A ) 또는 항산화 방어에 관여하는 보호 유전자를 과발현하여 스트레스 저항성을 향상시킵니다.
다음은 기술의 간단한 비교입니다:
| 기술 | 메커니즘 | 최적의 사용 용도 | 주요 고려사항 |
|---|---|---|---|
| CRISPRko | 영구적인 유전자 파괴 | 성장 억제제 제거 (TP53, PTEN) | 비가역적; 분화 잠재력 검증 필요 |
| CRISPRi | 전사 억제 (DNA 절단 없음) | 발견 스크린; 조절자 미세 조정 | 가역적; RNAi보다 낮은 비표적 효과 |
| CRISPRa | 전사 활성화 (DNA 절단 없음) | 보호 유전자 상향 조절 (HIF1A) | 안정적인 dCas9-활성화 전달 시스템 필요 |
초기 단계에서 목표를 식별하는 팀에게는, 풀드 CRISPRi 스크린이 대규모로 스트레스 저항 유전자를 발견하는 비용 효율적인 방법을 제공합니다.유망한 후보가 검증되면, CRISPRko는 생산에 적합한 영구적인 편집에 사용될 수 있습니다. 이러한 접근 방식은 서로를 보완하며, 순차적으로 사용하는 것이 이 분야에서 점점 더 최선의 관행으로 여겨지고 있습니다 [1][6].
배양육 연구에 맞춘 CRISPR 시약 및 바이오리액터 공급품을 조달하기 위해,
CRISPR 편집된 세포주 구현 및 검증
CRISPR 편집 설계 및 전달
표적 유전자를 식별한 후 다음 단계는 CRISPR 편집을 설계하고 전달하는 것입니다. 효과적인 유전자 파괴를 보장하기 위해 필수 엑손을 표적으로 하는 단일 가이드 RNA(sgRNA)를 만드는 데 중점을 두십시오. 이 접근 방식은 유전자를 완전히 제거할 가능성을 높이며, 잘린 부분적으로 기능하는 단백질을 생성하는 대신에 유전자를 완전히 제거할 가능성을 높입니다.듀얼 가이드 RNA 전략을 사용하면 녹아웃 효율성을 크게 향상시킬 수 있으며, 약 55%에서 95% 이상으로 증가시킬 수 있습니다 [8] .
선택한 전달 방법은 특정 세포 유형에 따라 달라집니다. 배양육 세포주에 대해서는 사전 조립된 Cas9 리보뉴클레오프로테인(RNP)이 종종 최선의 선택입니다. 이러한 RNP는 일시적이며, 전달 후 빠르게 분해되어 비표적 효과를 최소화하고 플라스미드 DNA 통합의 위험을 피할 수 있습니다 [8]. 풀드 스크린 또는 전이하기 어려운 1차 세포주가 관련된 경우, 렌티바이러스 전이는 신뢰할 수 있는 대안입니다. 렌티바이러스 시스템을 사용할 때, 연구자들은 일반적으로 약 0.3의 낮은 감염 다중성(MOI)을 유지하여 여러 통합을 피하고, 이는 다운스트림 분석을 복잡하게 만들 수 있습니다 [1].
최적의 결과를 위해, 세포가 로그 성장 단계에 있으며 형질감염 전에 70–90% 융합 상태에 있는지 확인하십시오. 전달 후, 제한 희석법이나 형광 활성 세포 분류(FACS)와 같은 방법을 사용하여 개별 클론을 분리하여 명확하고 모호하지 않은 검증을 보장하십시오. 마지막으로, 성공을 확인하기 위해 유전체, 단백질체, 기능적 수준에서 편집을 검증해야 합니다.
편집된 세포주 스크리닝 및 검증
편집된 세포주를 바이오리액터 조건. 으로 전환할 때 철저한 검증이 필수적입니다. 이 과정은 유전체, 단백질체, 기능적 세 가지 수준에서 스크리닝을 포함합니다. 이러한 단계 중 어느 하나라도 건너뛰면 생산 조건에서 실패할 수 있는 세포주를 선택할 위험이 증가합니다.
유전체 수준에서 초기 스크리닝은 T7E1 또는 Surveyor와 같은 불일치 분석을 사용하여 세포 풀에서 편집 빈도를 빠르게 추정할 수 있습니다.정확한 확인을 위해, Sanger 시퀀싱 또는 차세대 시퀀싱(NGS)을 통해 이중 대립 유전자 파괴 인델을 가진 클론을 식별하십시오 [7][8]. 단백질체 검증은 일반적으로 Western blot 분석을 사용하여 수행되며, 목표 단백질의 완전한 부재를 보장합니다. 예를 들어, 2025년에 수행된 연구에서는 TP53을 제거하면 경쟁 스크린의 30일째에 세포 수가 1,000배 이상 증가하여 배양 기간이 100일에서 약 200일로 두 배가 되는 것으로 나타났습니다 [1].
기능적 검증도 마찬가지로 중요합니다. 대사 생존력 및 증식률은 Alamar Blue 분석을 사용하여 평가할 수 있으며, 최대 200일 동안의 인구 두 배 시간(PDT)을 추적하여 복제 노화를 극복한 세포주를 식별하는 데 도움이 됩니다 [1]. 저산소 또는 미토콘드리아 스트레스에 견딜 수 있도록 조작된 세포주에 대해, FACS 기반 리포터 분석은 세포가 저산소 또는 영양 제한 조건에서 올바르게 반응하는지 확인할 수 있습니다 [6]. 또한, TP53 또는 PTEN 녹아웃이 있는 세포주는 분화 잠재력을 유지할 수 있는지 테스트해야 합니다. CD29 및 CD44와 같은 중간엽 줄기세포(MSC) 마커에 대한 유세포 분석은 이러한 세포가 줄기세포성을 유지하는지 확인할 수 있습니다 [1] .
| 검증 수준 | 방법 | 목적 |
|---|---|---|
| 유전체 | Sanger 시퀀싱 / NGS | 이중 대립 유전자 파괴 인델 확인[7][8] |
| 단백질체 | 웨스턴 블롯 | 표적 단백질의 완전한 부재 확인[7][8] |
| 표현형 | 유세포 분석 (CD29/CD44) | MSC 마커 및 줄기성 유지 확인[1] |
| 기능적 | Alamar Blue / PDT 추적 | 성장 동력학 및 대사 건강 평가[1] |
| 스트레스 | FACS 기반 리포터 분석 | 도전적인 조건에서 스트레스 반응 행동을 테스트하십시오 [6] |
편집된 세포주를 확장하기 전에 STR 프로파일링을 수행하여 세포의 정체성을 확인하고, 오염을 방지하기 위해 마이코플라스마 검사를 실시하십시오 [7]. 유효한 녹아웃 세포주를 생성하는 데는 일반적으로 약 3개월이 소요되며, 워크플로의 특정 단계를 반복할 가능성이 있습니다.
확대: 스트레스 저항성 세포주를 생산으로 이동
편집된 세포주를 생물반응기 조건으로 전환
유효성이 검증된 후, 편집된 세포주는 실험실 규모의 부착 배양에서 교반 탱크 생물반응기, 공기 리프트 반응기 또는 회전 벽 용기와 같은 현탁 시스템으로 전환해야 하며, 각각은 산업 규모의 배양육 생산을 지원할 수 있습니다 [2].
소의 중간엽 줄기세포(bMSC)와 같은 부착 의존 세포의 경우, 라미닌-511 코팅 마이크로캐리어를 사용하는 것이 현탁 배양으로의 실용적인 경로를 제공합니다 [3]. 이 전환 과정에서 CD29 및 CD44와 같은 MSC 마커를 모니터링하여 세포가 분화 잠재력을 유지하는 것이 중요합니다 [1].
확대의 중요한 단계는 배지를 재구성하는 것입니다. 혈청 기반 배지는 화학적으로 정의된 혈청이 없는 배지로 대체되어야 하며, 대규모 조건에서 세포 생존력을 유지하기 위해 지질, 비필수 아미노산 및 항산화제가 풍부하게 포함되어야 합니다 [4]. 특히, TP53 및 PTEN이 제거된 CRISPR 편집 세포주는 이 전환에 더 잘 적응할 수 있습니다. Nature Communications (2025)에 발표된 연구에 따르면 이러한 편집은 bMSC의 증식 수명을 약 100일에서 200일 이상으로 연장했으며, 80일째에 노화가 약 60%에서 10%로 감소했습니다 [1].
TP53 및 PTEN의 제거는 증식 속도를 크게 증가시키고 노화를 지연시켰습니다." - Nature Communications [1]
전환 과정에서 Alamar Blue 분석 및 qRT-PCR과 같은 도구는 세포 생존율을 추적하고 유전자 변형의 안정성을 보장하는 데 필수적입니다. 이러한 CRISPR 편집된 소 세포주는 평균 12%의 배가율 향상을 보였으며, 일부는 50일째에 50% 증가에 도달했습니다 [1]. 세포가 생물 반응기 조건에서 안정적인 성능을 보이면, 초점은 확장을 위한 특수 장비 소싱으로 전환될 수 있습니다.
확장을 위한 장비 및 재료 소싱
생산 수준의 생물 반응기 실행으로 확장하는 것은 조달에 상당한 도전을 도입합니다. 세포 적응을 확인한 후 필요한 재료와 장비를 확보하는 것이 우선 과제가 됩니다.단일 사용 교반 탱크 생물 반응기, 검증된 미세 운반체, 무혈청 배지 구성 요소 및 지속적인 클론 모니터링을 위한 FACS 시스템과 같은 항목은 매우 전문화되어 있으며 일반 실험실 공급업체에서는 종종 구할 수 없습니다.
육류 배양 산업을 위해 특별히 설계된
결론
CRISPR 기술은 연구 도구에서 배양육 생산에서 세포주를 조작하는 실용적인 방법으로 전환되었습니다. TP53 및 PTEN과 같은 주요 조절자를 표적으로 하여 연구자들은 세포 증식을 크게 연장하여 일반적인 배양 기간을 효과적으로 두 배로 늘렸습니다 [1] . 이러한 진전은 배양육의 대량 생산 가능성의 경계를 확장합니다.
그러나 편집된 세포주에서 대량 생산으로의 여정은 모든 단계에서 철저한 검증이 필요합니다. 조작된 세포가 근육과 지방 조직으로 분화할 수 있는 능력을 유지하는 것은 빠른 증식을 달성하는 것만큼이나 중요합니다. 그렇지 않으면 가장 빠르게 성장하는 세포주조차도 상업적 실현 가능성이 부족할 것입니다 [1] . 이것은 개선된 증식이 의미 있는 생산 결과로 이어진다는 것을 확인하기 위한 엄격한 검증 프로세스의 필요성을 강조합니다.
Nature Communications는 다음과 같이 이 접근 방식을 강화합니다:
"이러한 발견은 소 줄기 세포 특성을 최적화하기 위한 CRISPR 스크리닝의 유용성을 입증하며, 미래에 더 확장 가능한 배양육 생산으로 가는 길을 제공합니다." [1]
이러한 발전에도 불구하고, 조달과 같은 실질적인 문제는 진전을 방해할 수 있습니다. sgRNA 라이브러리, 일회용 생물 반응기 및 무혈청 배지에 대한 일반 공급업체에 대한 의존은 종종 호환성 문제와 지연을 초래합니다.
적합한 자재의 가용성은 유전자 공학 자체만큼이나 중요합니다. Nature Communications에 따르면, 배양육은 전통적인 고기에 대한 유망한 대안을 제시하지만, 확장성과 비용 효율성은 여전히 중요한 장애물로 남아 있습니다. CRISPR 기반의 공학은 엄격한 생물 공정 설계 및 플랫폼을 통한 간소화된 조달과 결합될 때
자주 묻는 질문
CRISPR 타겟을 선택하기 전에 어떤 바이오리액터 스트레스를 프로파일링해야 하나요?
배양육 생산에서 스트레스 저항성 세포주를 개발하기 위해 CRISPR 타겟을 선택할 때, 세포 성장과 생존에 영향을 미치는 주요 바이오리액터 스트레스를 평가하는 것이 중요합니다. 이러한 스트레스에는 다음이 포함됩니다:
- 전단 응력: 바이오리액터의 세포는 혼합 및 공기 공급으로 인한 기계적 힘에 자주 노출됩니다. 장기간의 전단 응력은 세포막을 손상시키고 성장을 저해할 수 있습니다.
- 산소 수준: 최적의 산소 농도를 유지하는 것이 중요합니다. 산소가 너무 적으면 에너지 생산이 제한될 수 있으며, 과도한 산소는 산화 스트레스를 초래할 수 있습니다.
- 영양소 가용성: 세포는 일관된 영양소 공급이 필요합니다. 불균형이나 고갈은 증식과 생산성을 저해할 수 있습니다.
- pH 변동: 세포는 좁은 pH 범위 내에서 번성합니다. 편차는 대사 과정과 효소 활동을 방해할 수 있습니다.
- 온도 변화: 온도의 약간의 변화도 세포 기능에 영향을 미쳐 스트레스나 생존 가능성을 감소시킬 수 있습니다.
- 폐기물 축적: 대사 부산물이 효율적으로 제거되지 않으면 독성이 되어 세포 성장을 억제할 수 있습니다.
이러한 스트레스 요인을 철저히 이해함으로써 연구자들은 중요한 스트레스 반응 경로를 정확히 찾아낼 수 있습니다. 이 지식은 CRISPR을 사용한 표적 유전자 변형을 가능하게 하여 세포주 회복력을 향상시키고 생물 반응기 조건에서 더 강력한 성능을 보장합니다.
빠른 성장 편집을 근육과 지방의 차별화와 어떻게 균형을 맞출 수 있습니까?
배양육 생산에서 빠른 성장을 근육과 지방의 차별화와 균형을 맞추기 위해서는 유전학과 배양 조건의 세심한 관리가 필요합니다. CRISPR 기술은 여기서 중심적인 역할을 하며, TP53 및 PTEN과 같은 유전자의 표적 수정이 가능합니다.. 이러한 조정은 세포 증식을 촉진하면서 세포가 근육과 지방 조직으로 분화할 수 있는 능력을 유지할 수 있게 합니다.
배양 조건의 미세 조정과 유전자 발현 조절도 원하는 균형을 달성하는 데 매우 중요합니다.
바이오리액터 규모 확장을 위한 최소한의 검증은 무엇인가요?
바이오리액터로 이동하기 전에, 유전자 변형 세포주가 향상된 성장률, 스트레스 내성, 분화 능력과 같은 안정적이고 바람직한 특성을 유지하는지 확인하는 것이 중요합니다. 이 검증 과정은 유전적 안정성을 평가하고 생물공정 조건에서 일관된 성능을 보장해야 합니다. 다중 오믹스 분석 및 스트레스 반응 프로파일링에서 지원 데이터를 확보하는 것이 이 평가의 핵심입니다. 고처리량 CRISPR 스크리닝을 사용하면 세포 증식과 수명을 향상시키는 유전자 편집을 정확히 찾아낼 수 있으며, 이러한 세포주는 대규모 배양육 생산에 더 적합합니다.










