

För forskare inom odlat köttproduktion är det viktigt att minimera apoptos för att förbättra cellernas livskraft och produktivitet i bioreaktorer. Stressfaktorer som näringsbrist, osmotiska obalanser och avfallsansamling utlöser ofta celldöd, vilket minskar avkastningen. Anti-apoptotiska gener kan mildra dessa utmaningar genom att förlänga cellernas livslängd under odling. Här är en snabb översikt över de främsta generna och deras roller:
- BCL-2: Förhindrar bildandet av mitokondriella porer, vilket blockerar apoptos vid dess initiering. Effektiv för odifferentierade celler men kräver noggrann balans med pro-apoptotiska proteiner.
- BCL-xL: Skyddar celler under differentiering och stöder energimetabolism. Idealisk för högstressfaser i bioreaktorer.
- MCL-1: Erbjuder snabb respons på näringsförändringar och förblir stabil under differentiering. Fungerar bra i kombination med andra gener.
- BIRC5 (Survivin) : Hämmar kaspaser för att blockera apoptos nedströms. Stödjer proliferation i snabbt delande celler.
- XIAP: En potent kaspashämmare effektiv under extrema stressförhållanden, såsom högdensitetskulturer. Övervakning av dessa förhållanden kräver val av sensorer för bioreaktorer för odlat kött för att spåra näringsnivåer och avfallsackumulering i realtid.
Snabb Jämförelse
| Gen | Nyckelroll | Stabilitet under Differentiering | Bästa Användningsfall |
|---|---|---|---|
| BCL-2 | Blockerar tidig apoptos (BAX/BAK) | Stabil | Bevara odifferentierade celler |
| BCL-xL | Förhindrar kaspasaktivering, stöder metabolism | Steg-specifik | Differentierande celler under stress |
| MCL-1 | Snabb respons på näringsförändringar | Stabil | Överlevnad i flera steg |
| BIRC5 | Hämmar kaspaser nedströms | Minskar med differentiering | Snabbt delande celler |
| XIAP | Bred kaspashämning | Stabil | Högstress bioreaktorvillkor |
1.BCL-2
BCL-2 är en välstuderad anti-apoptotisk gen som spelar en nyckelroll i den intrinsiska (mitokondriella) apoptosvägen. Denna väg är en viktig mekanism för celldöd, ofta utlöst i odlade köttceller under bioreaktorstress som näringsbrist eller låga syrenivåer.
BCL-2 fungerar genom att binda till och neutralisera pro-apoptotiska proteiner som BAX och BAK. Denna åtgärd förhindrar bildandet av mitokondriella porer, stoppar frisättningen av cytokrom c och avbryter den nedströms kaskaden av apoptos. Denna mekanism är avgörande för att förlänga cellernas livslängd i produktionen av odlat kött. Som Rønning SB et al. förklarar:
"Förhållandet mellan Bcl-2 och Bax bestämmer cellernas känslighet för att genomgå apoptos."[5]
Utöver sin mitokondriella roll finns BCL-2 också i det endoplasmatiska retiklet (ER).Här minskar det kalciumnivåerna och hämmar IP3-receptor-medierad kalciumfrisättning, vilket minskar kalciuminducerad apoptos – ett vanligt problem i högdensitetsbioreaktorkulturer[4]. Att hantera dessa skalningsutmaningar är ett primärt fokus för branschen. Denna dubbla lokalisering gör det möjligt för BCL-2 att skydda celler från flera apoptosutlösare.
Den molekylära strukturen av BCL-2, bestående av ett åtta-alfa-helix-bunt och fyra väldefinierade BH-domäner, gör det till en e
Det finns dock en kritisk varning: balansen mellan BCL-2 och pro-apoptotiska proteiner som BAX måste hanteras noggrant. Även höga nivåer av BCL-2-uttryck kan misslyckas med att förhindra apoptos om pro-apoptotiska signaler blir för starka[2]. Att övervaka denna balans är avgörande för att uppnå optimal cellviabilitet.
sbb-itb-ffee270
2. BCL-xL
BCL-xL, kodad av BCL2L1 genen, spelar en central roll i BCL-2-familjen genom att lokaliseras till det yttre mitokondriemembranet och förhindra apoptos. Den uppnår detta genom att motverka pro-apoptotiska proteiner som BAX och BAK.Dessutom hämmar det klyvt kaspas-3 (CASP3), vilket är viktigt för att stoppa celldöd. Denna mekanism är särskilt värdefull i högdensitetsbioreaktorkulturer , där metabolisk stress kan hota cellviabiliteten.
Intressant nog överensstämmer BCL-xL:s aktivitet med specifika stadier av differentiering. Under vissa faser ökar dess uttryck, medan andra anti-apoptotiska proteiner, såsom BCL-2 och MCL-1, förblir oförändrade. Detta understryker dess betydelse för att upprätthålla cellöverlevnad under differentiering. Som noterat i Cell Death & Disease:
"BCL-xL/BCL2L1 är ett kritiskt anti-apoptotiskt protein som främjar överlevnaden av differentierande... celler." [2]
Utöver sin roll i apoptos stödjer BCL-xL cellens energimetabolism. Det förbättrar både glykolys och oxidativ fosforylering, vilket säkerställer hög metabolisk aktivitet.Hämning av BCL-xL har visat sig minska uttrycket av metabola gener och sänka både basal och maximal mitokondriell respiration. Denna funktion är särskilt viktig för odlade köttceller, som är beroende av en hållbar metabolisk produktion.
BCL-xL är mycket kompatibel med genredigeringsstrategier som vanligtvis används i forskning om odlat kött. Tekniker som lentiviral transduktion möjliggör stabil integration av BCL2L1-genen, medan doxycyklin-inducerbara CRISPR/Cas9-system ger exakt tidsmässig kontroll över dess uttryck [2] [6]. Denna nivå av precision hanteras ofta genom avancerad bioprocesskontrollprogramvara. Dessa egenskaper gör BCL-xL till en stark kandidat för att förbättra cellinjeviabiliteten i produktionen av odlat kött.
För differentieringsstadier med höga metabola krav kan BCL-xL vara mer effektiv än BCL-2.Forskare kan använda hämmaren WEHI-539 för att testa en cellinjes beroende av BCL-xL innan de går vidare med permanenta genetiska modifieringar [2]. Dessutom kan samuttryck av BCL-xL med MCL-1 ytterligare förbättra cellöverlevnaden, eftersom dessa proteiner har observerats arbeta synergistiskt i vissa resistenta celltyper [6].
3. MCL-1
MCL-1 (Myeloid Cell Leukaemia-1) spelar en nyckelroll i regleringen av den intrinsiska apoptotiska vägen. Belägen på det yttre mitokondriemembranet, förhindrar den apoptos genom att binda och sekvestrera de pro-apoptotiska proteinerna BAX och BAK, vilket stoppar deras oligomerisering och efterföljande membranpermeabilisering. Denna åtgärd blockerar frisättningen av cytokrom c, vilket stoppar den apoptotiska kaskaden innan den når exekveringsfasen [8] . Dessutom binder MCL-1 BH3-endast proteiner - såsom Bim, PUMA och NOXA - med hög affinitet [8]. Likt BCL-2 och BCL-xL är MCL-1 avgörande för att motverka apoptotiska signaler, särskilt under bioreaktorstress.
En av MCL-1:s unika egenskaper är dess korta halveringstid, vilket gör dess uttryck mycket känsligt för näringstillgänglighet och metaboliska signaler, särskilt genom AMPK/mTOR-vägen. Studier indikerar att en 25% minskning av kaloriintaget kan minska MCL-1-översättning med cirka 39% ± 10% [7]. Denna känslighet är särskilt relevant för odlad köttproduktion, där fluktuationer i tillväxtmediets sammansättning eller näringsbrist under storskaliga suspensionkulturer (som kräver noggrann produktionsskaleplanering) kan avsevärt sänka MCL-1-nivåerna.Sådana minskningar komprometterar cellens livskraft, vilket undergräver förbättringarna i IVCC (integral viable cell concentration) som uppnåtts genom anti-apoptotiska strategier. För att mildra detta är serumfria medieformuleringar som stödjer robust mTORC1-aktivitet väsentliga [7] .
En annan anmärkningsvärd egenskap hos MCL-1 är dess stabilitet under differentiering. I pankreatiska progenitormodeller förblev MCL-1-uttrycket stabilt under ett 17-dagars differentieringsprotokoll, till skillnad från BCL-xL, som visade stegberoende variation [2]. Denna stabilitet gör MCL-1 särskilt fördelaktig för odlad köttapplikationer, där celler behöver överleva flera mognadsstadier utan att kräva exakt tidsbestämda interventioner.&
Genredigeringsverktyg kan användas för att modifiera MCL-1, precis som andra anti-apoptotiska gener, vilket gör det till ett mångsidigt mål för cellinjeutveckling.
När det används i kombination med andra anti-apoptotiska gener, erbjuder MCL-1 ytterligare fördelar. Till exempel har kombinationen av MCL-1 med BCL-xL visat synergistiska effekter - samtidig hämning av båda proteinerna minskade EC50 för överlevnadsdroger från cirka 10 μM till mindre än 20 nM [6]. Denna metod kan avsevärt förbättra cellöverlevnad under de högstressiga faserna av odlat köttproduktion.
4. BIRC5 (Survivin)
BIRC5, ofta kallad Survivin, är en medlem av Inhibitor of Apoptosis (IAP) proteinfamiljen [2]. Till skillnad från BCL-2 familjeproteiner, som verkar vid mitokondriemembranet för att förhindra initiering av apoptos, verkar BIRC5 längre nedströms. Det blockerar kaspaserna som ansvarar för att utföra apoptos, vilket effektivt fungerar som en sista försvarslinje mot programmerad celldöd [10].
I suspensionskulturer kan stressfaktorer som näringsbrist, ansamling av metaboliskt avfall och mekanisk skjuvspänning utlösa apoptos. Genom att hämma kaspasaktivitet i detta senare skede hjälper överuttryck av BIRC5 till att förlänga cellens livskraft och produktivitet. Detta resulterar i en förbättring av tidsintegralen av livskraftig cellkoncentration - en nyckelmetrik för att optimera cellkulturens prestanda [9] . Eric Baek, en forskare vid KAIST, förklarar:
"Att förbättra tidsintegralen av livskraftig cellkoncentration genom att övervinna celldöd, nämligen apoptos, är en av de mest använda strategierna för effektiv produktion av terapeutiska proteiner [och celler]." [9]
Denna nedströmsintervention har visat sig förbättra bioreaktorns avkastning i odlade köttcellinjer, inklusive porcina satellitceller och bovint myoblaster.
Den mest effektiva strategin involverar kombinatorisk ingenjörskonst, som parar BIRC5 med mitokondriella skyddare som BCL-2 eller BCL-xL. Professor Michael Betenbaugh från Johns Hopkins University lyfter fram detta tillvägagångssätt:
"Strategier som blockerar celldöd vid flera punkter längs kaskaden kan begränsa förstärkningen av dessa apoptossignaler." [10]
Genom att kombinera BIRC5:s kaspashämning med uppströms mitokondriellt skydd kan forskare etablera ett flerskiktat försvar mot apoptos.
BIRC5 integreras också sömlöst i genredigeringsarbetsflöden.CRISPR/Cas9 är den ledande metoden för att skapa stabila cellinjer med överuttryck [9], även om zinkfingernukleaser erbjuder ett precist alternativ. siRNA kan användas för validering av vägar innan man åtar sig genomisk integration [9].
5. XIAP
XIAP (X-linked inhibitor of apoptosis) erkänns som den mest potenta caspashämmaren inom IAP (inhibitor of apoptosis protein) familjen. Tillsammans med gener som BCL-2 och MCL-1 spelar XIAP en kritisk roll i att rikta in sig på apoptos i dess exekveringsfas. Som framhävs i Genes & Development :
"XIAP anses vara den mest potenta caspashämmaren in vitro." [12]
XIAP använder två distinkta mekanismer för att hämma apoptos. Först blockerar dess BIR2-domän och länkområde effektorcaspaserna-3 och -7.Andra, dess BIR3-domän hämmar kaspas-9, vilket effektivt stoppar den intrinsiska mitokondriella apoptotiska vägen. Dessutom underlättar dess C-terminala RING-domän ubiquitinering och efterföljande proteasomal nedbrytning av mål-kaspaser [11]. Genom att ingripa i både intrinsiska och extrinsiska apoptotiska vägar, visar XIAP sig vara mycket effektiv i att hantera apoptosutlösare som näringsbrist, metaboliska biprodukter och mekanisk stress - faktorer som ofta förekommer i odlade köttproduktionssystem. Dess funktionalitet förbättras ytterligare av dess starka bevarande över arter.
Till exempel delar mänsklig XIAP 87,7% proteinidentitet med Bos taurus (nötkreatur) och 89,5% med Mus musculus (mus) [11] . Denna höga likhet gör att forskning från däggdjursmodellsystem kan tillämpas på ett tillförlitligt sätt på cellinjer som används i odlad köttproduktion.
XIAP kan regleras med verktyg som shRNA, antisense-oligonukleotider eller CRISPR/Cas9 [11]. Under extrem stress kan dess RING-domän inducera självubikvitinering [12], medan endogena inhibitorer som SMAC/DIABLO och HTRA2 kan förflytta XIAP från kaspaser [11][13]. Dessa fynd gör XIAP till ett attraktivt mål för genredigeringsmetoder som syftar till att optimera cellinjer för utveckling av odlat kött.
Jämförelse av anti-apoptotiska gener i korthet
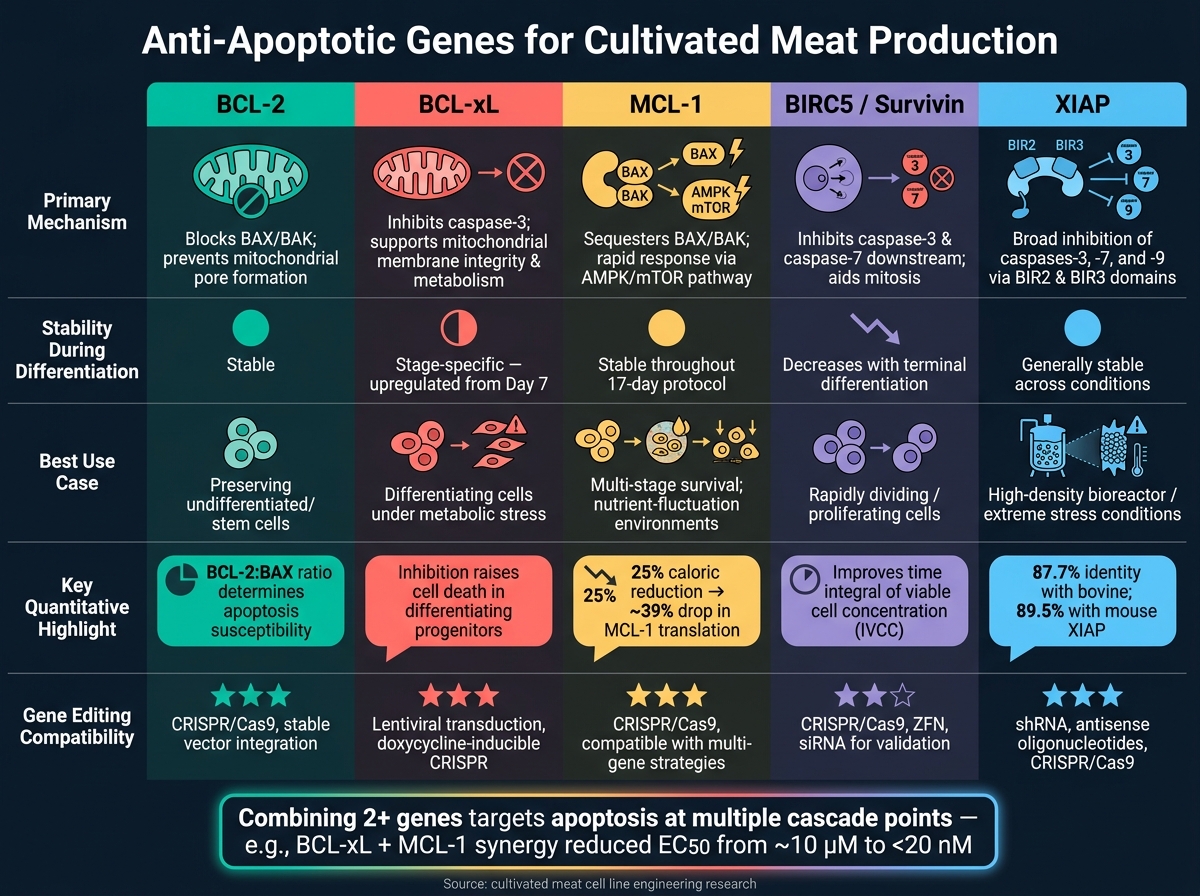
Anti-apoptotiska gener för odlat kött: Jämförelse sida vid sida
När man arbetar med produktion av odlat kött kan förståelsen av hur olika anti-apoptotiska gener fungerar hjälpa till att finjustera cellinjeingenjörskonsten. Varje gen har sin unika mekanism, beteende under differentiering och potentiella tillämpningar. Tabellen nedan sammanfattar dessa skillnader, vilket gör det enklare att avgöra vilken gen - eller kombination av gener - som kan fungera bäst för dina behov.
| Gen | Primär mekanism | Expressionsstabilitet | Rapporterad livskraftspåverkan | Redigeringskompatibilitet |
|---|---|---|---|---|
| BCL-2 | Blockerar pro-apoptotiska BAX/BAK och säkerställer överlevnad av odifferentierade celler [2] | Förblir relativt stabil under differentiering [2] | Väsentlig för att bevara den initiala stamcellspoolen [2] | Hög kompatibilitet med redigeringsverktyg |
| BCL-xL | Hämmar klyvd kaspas-3; upprätthåller mitokondriell membranintegritet och metabolism [2] | Uppreglerad från och med dag 7 av differentiering [2] | Kritiskt för att stödja differentierande progenitorer; dess hämning ökar celldöd [2] | Hög kompatibilitet med redigeringsverktyg |
| MCL-1 | Modulerar pro-apoptotiska signaler som en del av BCL-2-familjen [2] | Uttrycket förblir stabilt under linjespecifikation [2] | Erbjuder breda överlevnadsfördelar men saknar stadiespecifika effekter som BCL-xL [2] | Hög kompatibilitet med redigeringsverktyg |
| BIRC5 (Survivin) | Blockerar kaspas-3 och kaspas-7; hjälper till med kromosomal segregation under mitos | Hög i prolifererande celler; minskar med terminal differentiering | Stödjer överlevnad och proliferation i snabbt delande celler | Kompatibel med både shRNA knockdown och CRISPR-redigering |
| XIAP | Hämmar flera kaspaser, vilket ger ett brett apoptotiskt skydd | Generellt stabil under olika förhållanden | Särskilt effektiv under stress, såsom högdensitetsbioreaktorvillkor | Hög kompatibilitet med redigeringsverktyg |
BCL-xL utmärker sig för sin dubbla roll i att främja cellöverlevnad och stödja metabolisk aktivitet, särskilt under den kritiska differentieringsfasen när pro-apoptotiska proteiner som BAK naturligt minskar. BCL-2, å andra sidan, är idealisk för att bevara odifferentierade celler, medan XIAP ger ett brett skydd, särskilt i stressiga miljöer som högdensitetskulturer.
Inget enskilt gen fungerar bäst i alla scenarier. Till exempel är BIRC5 särskilt användbar i situationer som kräver snabb celldelning. I praktiken erbjuder kombinationen av två eller fler gener ofta det mest effektiva skyddet, genom att samtidigt hantera en mängd olika apoptotiska utlösare.
Dessa fynd ger en grund för att integrera dessa gener i cellinjeingenjörsstrategier för odlat köttproduktion. Detta inkluderar att välja rätt odlade köttinsatser för att säkerställa skalbarhet.
Använda Dessa Gener i Cellinjeingenjör för Odlat Kött
För att förbättra cellviabiliteten i odlat köttproduktion är det avgörande att integrera nyckelgener strategiskt.Det räcker inte att identifiera anti-apoptotiska gener - deras effektiva inkorporering i cellinjer är det som gör skillnaden. Två huvudstrategier används ofta: överuttrycka anti-apoptotiska gener som BCL-2, BCL-xL, och MCL-1 för att förbättra cellöverlevnad, eller slå ut pro-apoptotiska gener såsom BAX, BAK, och BOK för att eliminera drivkrafterna för celldöd. Att kombinera dessa metoder resulterar ofta i cellinjer som är bättre lämpade för storskalig produktion [1].
Moderna genredigeringsverktyg som CRISPR/Cas9 möjliggör samtidiga redigeringar, såsom att slå ut Bak1, Bax, och Bok i ett steg. Alternativ som ZFNs eller RNA-interferens kan användas för att tillfälligt minska aktiviteten hos kaspaser ( e.g. caspaser-3, -7, -8 och -9). För överuttrycksstrategier säkerställer syntetiska promotorer konsekventa och höga uttrycksnivåer av gener som BCL-2 under uppskalning, vilket är kritiskt för att upprätthålla cellprestanda i fed-batch eller kontinuerliga odlingssystem . Dessa kombinerade metoder stärker cellinjeutvecklingen för odlat köttapplikationer.
Sådana genetiska modifieringar påverkar direkt förbättrad integral livskraftig cellkoncentration (IVCC), en nyckelmetrik i produktionen av odlat kött. Celldöd är mest uttalad under de första fem dagarna av differentiering, vilket gör tidiga interventioner med gener som BCL-2 eller BCL-xL väsentliga. Forskning publicerad i Cell Death & Disease belyser att BCL-xL uttryck ökar när celler differentierar, vilket indikerar att mer mogna progenitorer är starkt beroende av dess skyddande roll [2] . Genom att övervaka uttrycksnivåerna av BCL-2 familjegener under tillväxtfaserna kan interventioner tidsbestämmas exakt för maximal effekt.
"Genom att etablera stabila cellinjer som överuttrycker antiapoptotiska gener eller nedreglerar proapoptotiska gener kan slutproduktens avkastning förbättras eftersom cellerna blir mer motståndskraftiga mot miljömässiga påfrestningar." - Gyun Min Lee et al. [1]
För bioreaktorbaserad produktion måste celler också konstrueras för att tåla hyperosmotisk stress och näringsbrist. Innan uppskalning är det viktigt att validera genetiska redigeringar med verktyg som Western blot eller FACS. För forskare som söker specialiserade cellinjer eller genetiska material anpassade för högdensitetsbioreaktormiljöer, erbjuder plattformar som
Slutsats
Att välja anti-apoptotiska gener för odlade köttcellinjer kräver en skräddarsydd strategi. Gener som BCL-2, BCL-xL, och MCL-1 spelar var och en unika roller i att skydda celler, men deras framgång beror på faktorer som celltyp, utvecklingsstadium och de specifika påfrestningar som uppstår under produktionen. Som framhävs i forskning:
"balansen mellan de anti-apoptotiska och pro-apoptotiska medlemmarna avgör slutligen om en cell lever eller dör" [2]
Utöver överlevnad bevarar anti-apoptotisk ingenjörskonst också metaboliska funktioner. Till exempel är proteiner som BCL-xL nära kopplade till att upprätthålla glykolys och oxidativ fosforylering. Dåligt genomförda interventioner kan dock störa dessa kritiska processer [2]. Att säkerställa att konstruerade cellinjer bibehåller sin avsedda identitet och metaboliska aktivitet under hela produktionen är ett avgörande, men ibland förbisett, steg. Dessa insikter formar framtiden för cellinjeingenjörskonst.
Nya multi-genmetoder dyker upp, som kombinerar överuttryck av skyddande gener med CRISPR-knockouts av pro-apoptotiska gener som BAX, BAK1, och BOK för att skapa mer robusta cellinjer för industriellt bruk [1]. Verktyg för metabolisk profilering, såsom bioenergetiska tester, blir nödvändiga för att bekräfta att dessa genetiska modifieringar förbättrar cellens övergripande prestanda. För forskare som söker porcina cellinjer , genetiska material eller bioreaktorutrustning,
Vanliga frågor
Vilken anti-apoptotisk gen bör jag börja med för min cellinje?
BCL-2 föreslås ofta som en startpunkt när man arbetar med cellinjer. Denna välundersökta anti-apoptotiska gen är känd för sin förmåga att förbättra cellöverlevnad, vilket gör den till ett populärt alternativ inom odlat köttforskning. Dess funktion i att stödja cellviabilitet gör den till ett praktiskt val för tidiga experiment.
Är det bättre att överuttrycka anti-apoptotiska gener eller slå ut pro-apoptotiska gener?
Inom produktion av odlat kött ger ökad uttryck av anti-apoptotiska gener, såsom medlemmar av BCL-2-familjen som BCL-xL, ofta bättre resultat än att inaktivera pro-apoptotiska gener. Denna strategi stödjer både cellöverlevnad och proliferation - nyckelfaktorer för att skala upp produktionen - samtidigt som cellens naturliga regleringssystem bevaras.
Genom att öka aktiviteten hos anti-apoptotiska gener får celler större motståndskraft mot apoptos, särskilt under stressiga förhållanden. Detta gör det till en mer pålitlig och säkrare metod för att upprätthålla cellviabilitet under odlingsprocessen.
Hur kan jag bekräfta att en anti-apoptotisk redigering förbättrar IVCC i min bioreaktor?
För att avgöra om en anti-apoptotisk genredigering förbättrar in vitro cellviabilitet och proliferation (IVCC), behöver du en systematisk metod:
- Utvärdera viabilitet och proliferationshastigheter: Använd metoder som cellräkning eller flödescytometri för att mäta dessa hastigheter både före och efter genredigeringen.
- Verifiera genuttryck: Tekniker som qPCR eller Western blotting kan bekräfta det framgångsrika uttrycket av den riktade genen.
- Övervaka apoptosmarkörer: Kontrollera markörer som kaspasaktivitet för att säkerställa att redigeringen effektivt minskar apoptos.
För en komplett utvärdering är det kritiskt att testa den långsiktiga stabiliteten och proliferationen av de redigerade cellerna i en bioreaktor. Detta säkerställer att förbättringarna kvarstår över flera odlingscykler.










