

Histonmodifikationer er kemiske ændringer af proteiner, der påvirker genaktivitet uden at ændre DNA. Disse modifikationer er afgørende for udviklingen af cellelinjer, der anvendes i produktionen af dyrket kød, idet de hjælper celler med at vokse, bevare deres identitet og differentiere sig til muskelvæv. Artiklen udforsker, hvordan specifikke histonmærker som H3K4me3 (genaktivering), H3K27ac (enhanceraktivitet) og H3K27me3 (genundertrykkelse) regulerer celleadfærd.
Vigtige punkter dækket:
- H3K4me3 understøtter aktive gener og hurtig differentiering.
- H3K27ac kontrollerer enhancere for genekspression under vækstfaser.
- H3K27me3 sikrer, at uønskede genprogrammer forbliver inaktive.
- Kromatintilstande, formet af disse mærker, varierer på tværs af arter og celletyper, hvilket påvirker produktionskvaliteten.
Artiklen fremhæver også nylig forskning, herunder hvordan positionsbestemt genekspression i svineceller påvirker kødets kvalitet, og hvordan målrettet epigenetisk redigering kan forbedre cellelinjens ydeevne. Fremtidige retninger inkluderer at forfine epigenetiske værktøjer og studere kromatintilstande for at optimere produktions effektivitet og skala.
Histonmodifikationer forklaret | Acetylering, Methylering & Genregulering
sbb-itb-ffee270
Typer af histonmodifikationer og deres funktioner
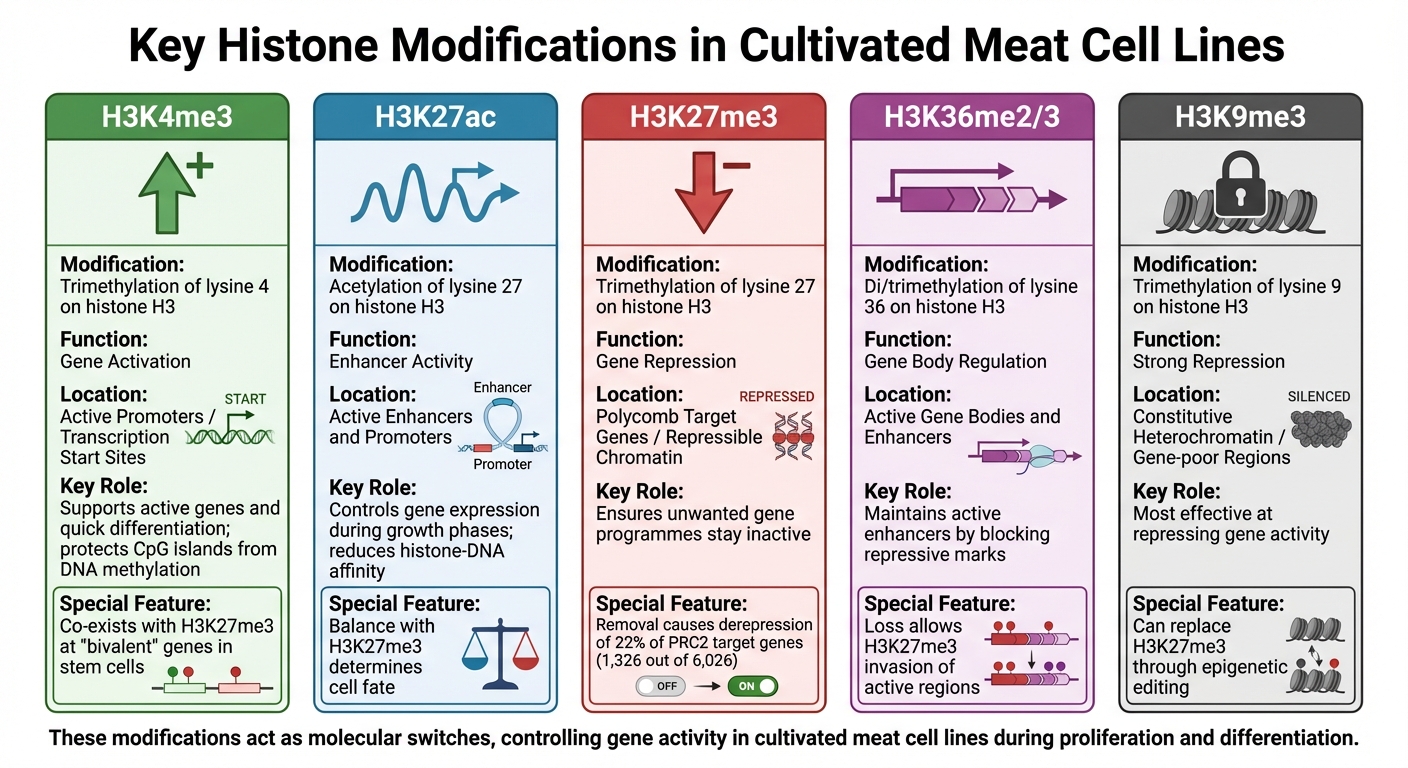
Vigtige histonmodifikationer i dyrkede kød cellelinjer: Funktioner og genomiske kontekster
Histonmodifikationer spiller en afgørende rolle i reguleringen af genaktivitet, idet de fungerer som molekylære kontakter til at kontrollere, om gener er tændt eller slukket i dyrkede kød cellelinjer.Disse kemiske mærker - hovedsageligt methylering og acetylering - binder sig til specifikke rester på histoner og skaber distinkte genomiske mønstre. Hver modifikation har en specifik funktion, og ved at forstå disse roller kan forskere bedre forudsige og påvirke celleadfærd under produktion. Denne viden er essentiel for at optimere processer i dyrket kød bioprocessering.
Her er en oversigt over de vigtigste histonmodifikationer, der påvirker genregulering i dyrkede kød cellelinjer.
H3K4me3 og Genaktivering
H3K4me3 (trimethylering af lysin 4 på histon H3) er forbundet med aktive genpromotorer og letter transkription ved genstartsteder, især for gener involveret i cellevækst og metabolisme. Denne modifikation beskytter også CpG-ø-promotorer mod ny DNA-methylering, hvilket sikrer, at essentielle gener forbliver tilgængelige for transkription [4].
I primære eller immortaliserede cellelinjer brugt til dyrket kød, sameksisterer H3K4me3 ofte med undertrykkende mærker som H3K27me3 ved "bivalente" gener. Disse gener forbliver klar til aktivering, hvilket muliggør hurtig differentiering til muskelvæv, når det er nødvendigt [4].
Interessant nok interagerer H3K4me3 med andre modifikationer. For eksempel kan deponeringen af H3K36me3 hæmme H3K4 methyltransferaser, reducere H3K4me3 niveauer ved promotorer og ændre genekspressionsmønstre [4].
H3K27ac og Enhancer Aktivitet
H3K27ac (acetylering af lysin 27 på histon H3) er en markør for aktive enhancere og promotorer. Ved at reducere affiniteten mellem histoner og DNA skaber H3K27ac et miljø, der fremmer transkription [5]. I dyrkede kødcellinjer bestemmer ændringer i H3K27ac-niveauer under forskellige vækstfaser, hvilke gener der udtrykkes, når cellerne bevæger sig fra proliferation til differentiering.
Balancen mellem H3K27ac og repressive modifikationer som H3K27me3 er afgørende for at bestemme celleskæbne. For eksempel kan tabet af H3K36me2, som understøtter enhancer-aktivitet, tillade H3K27me3 at invadere tidligere aktive regioner, reducere H3K27ac-niveauer og tavse mål-gener [5].
H3K27me3 og Gen Repression
H3K27me3 (trimethylering af lysin 27 på histon H3) er et repressivt mærke, der fremmer lukkede kromatinstrukturer og effektivt tavser gener. Denne modifikation, katalyseret af Polycomb Repressive Complex 2 (PRC2), er kritisk for at opretholde repressionen af tusindvis af udviklingsgener [4].
I dyrkede kød cellelinjer sikrer H3K27me3, at uønskede genprogrammer forbliver inaktive under specifikke vækstfaser, hvilket bevarer cellernes tilsigtede identitet.
"H3K27me3, sammen med H2AK119ub1, er afgørende for at opretholde den transkriptionelle repression af flere tusinde Polycomb-målgener." - Nature Communications [4]
Forskning har vist, at fjernelse af H3K27me3 i musembryonale stamceller resulterer i derepression af omkring 22% (1.326 ud af 6.026) af PRC2-målgenerne [4]. For dyrket kød kan kontrol af denne modifikation hjælpe med at undertrykke alternative cellefater, såsom dannelse af fedt eller bindevæv, mens der fokuseres på muskeludvikling.
| Histonmodifikation | Regulatorisk funktion | Genomisk kontekst |
|---|---|---|
| H3K4me3 | Genaktivering | Aktive promotorer / transkriptionsstartsteder |
| H3K27ac | Enhanceraktivitet | Aktive enhancere og promotorer |
| H3K27me3 | Genundertrykkelse | Polycomb-målgener / undertrykkeligt kromatin |
| H3K36me2/3 | Genkropsregulering | Aktive genkroppe og enhancere |
| H3K9me3 | Stærk undertrykkelse | Konstitutivt heterokromatin / genfattige regioner |
Kromatintilstande i dyrkede kødcellinjer
Histonmodifikationer virker ikke alene - de kombineres for at danne kromatintilstande, som er unikke genomiske miljøer, der kontrollerer geners tilgængelighed.Disse tilstande spiller en afgørende rolle i at forme adfærden af dyrkede kødcellinjer under både ekspansion og differentiering, hvilket gør dem vigtige for at optimere bioprocessering.
Identificering af kromatintilstande gennem histonmærker
Forskere kortlægger kromatintilstande ved at studere kombinationer af histonmærker som H3K4me3, H3K27ac og H3K27me3. For eksempel, i porcine føtale fibroblaster (PFF) og trofektoderm (PTr2) celler, 10 forskellige kromatintilstande er blevet identificeret, inklusive aktive transkriptionsstartsteder, bivalente promotorer og formodede enhancere [6] . Disse tilstande hjælper med at forudsige genaktivitet.
Enhancertilstande, markeret overvejende af H3K27ac i intergeniske og introniske regioner, er ofte co-beriget med kromatin-remodelleringsproteinet BRG1 [6].
En særlig bemærkelsesværdig egenskab er tilstedeværelsen af brede H3K4me3-domæner, som spænder over områder på 4 kb eller mere. Disse domæner repræsenterer kun 1,7% til 1,8% af alle forudsagte transkriptionsstartsteder i svinecellelinjer, men er afgørende for at markere udviklings- og vævsspecifikke gener [6]. Interessant nok er i svinefostrefibroblaster 52% af generne markeret af disse brede domæner vævsspecifikke, sammenlignet med kun 25% i PTr2-celler [6].
"Disse fund forbedrer vores forståelse af det epigenetiske landskab, der er til stede i tidlig svineudvikling, og giver indsigt i, hvordan variationer i kromatintilstand er forbundet med celleidentitet." - BMC Epigenetik & Kromatin [6]
Disse kromatintilstandsprofiler adskiller sig ikke kun inden for en enkelt art, men varierer også på tværs af de forskellige dyrecellelinjer, der anvendes i produktionen af dyrket kød.
Kromatinforskelle på tværs af dyrecellelinjer
Kromatintilstandsmønstre ændrer sig betydeligt afhængigt af arten og celletype, der anvendes i produktionen af dyrket kød. For eksempel, i kyllingecellelinjer, udgør H3K4me3 30% til 55% af dens genomiske tilstedeværelse ved genpromotorer [7]. Men i kyllingens primordiale kimceller (PGCs) falder H3K4me3-niveauerne betydeligt sammenlignet med pluripotente celler. Denne reduktion understøtter overgangen af bivalente tilstande til repressive tilstande under kimlinjespecifikation [7].
Porcine trophectoderm (PTr2) celler viser højere H3K27ac niveauer i promotorregioner (57,36%) sammenlignet med føtale fibroblaster (41,58%), mens H3K27me3 berigelse er lavere i PTr2 celler (7,77%) end i PFF celler (22%) [6]. Disse variationer afspejler de forskellige epigenetiske behov i hver udviklingsfase og påvirker, hvordan disse celler reagerer på kulturforhold.
I bovine satellitceller, differentiation til en "reserve celle" skæbne (Pax7+/Ki-67-) drives af hvilende kromatintilstande reguleret af NOTCH og MAPK/ERK signalering. Denne proces reducerer dog proteinudbyttet [3]. Sådan variabilitet understreger, hvordan kromatintilstande direkte påvirker produktionseffektiviteten. At opnå en dybere forståelse af disse forskelle er afgørende for at finjustere cellelinjens ydeevne i dyrket kødproduktion.
Brug af histonmodifikationer til at forbedre cellelinjer
Med udgangspunkt i vores viden om kromatintilstande, lad os dykke ned i, hvordan målrettede histonmodifikationer direkte kan forbedre ydeevnen af dyrkede kød cellelinjer.
Forøgelse af proliferation og tilpasning til suspension vækst
Justering af histonmærker kan betydeligt øge celleproliferation og hjælpe celler med at overgå fra adherente til suspension vækst. Dette skift er afgørende for bioreaktorsystemer til dyrket kød. For eksempel, reduktion af H3K36 methylering gør fibroblaster mindre responsive over for TGFβ, hvilket resulterer i en mere fleksibel cellestatus [1].
I december 2022 opnåede forskere hos Believer Meats et gennembrud med kyllingefibroblaster (HUN-CF-2 og HUN-CF-4).De demonstrerede spontan immortalisation i serumfri suspensionskulturer, nåede 100 millioner celler pr. ml (10⁸ celler/ml) og opnåede udbytter på 36% w/v. Teamet, ledet af Yaakov Nahmias, brugte lecithin - et fødevaresikkert lille molekyle - til at aktivere PPARγ-vejen og fremme fedtdannelse uden at stole på genetisk modifikation. Deres dyrkede kyllingeprototype fik en sensorisk vurdering på 4,5 ud af 5,0 [2].
"Immortalisering uden genetisk modifikation og højtydende fremstilling er afgørende for markedsrealiseringen af kultiveret kød." - Yaakov Nahmias, Chief Scientific Officer, Believer Meats [2]
Disse fund fremhæver potentialet i præcise epigenetiske værktøjer til yderligere at forfine cellelinjeudvikling.
Præcision i epigenetisk redigering
For at supplere disse cellulære ændringer tillader præcise epigenetiske redigeringsmetoder målrettet manipulation af histonmærker. En undersøgelse fra 2025 på musembryonale stamceller viste, at en kimær rekrutter (S12N) fusioneret med katalytiske domæner fra SUV39H2 eller SETD2 kunne erstatte H3K27me3 med H3K9me3 eller H3K36me3 ved tusindvis af gener. Blandt disse viste H3K9me3 sig at være mere effektiv til at undertrykke genaktivitet [8].
Dog afhænger succesen af disse modifikationer stærkt af det eksisterende kromatinmiljø. For eksempel kan resterende H3K4me3 ved genpromotorer blokere DNA-methyleringsmaskineriet, hvilket gør det sværere at opnå den ønskede genslukkelse [8]. Dette antyder, at optimering af celleydelse ofte kræver justering af flere histonmærker på samme tid snarere end at fokusere på en enkelt modifikation.
Konklusion og Fremtidige Retninger
Vigtige Punkter
Histonmodifikationer spiller en kritisk rolle som molekylære kontakter, der kontrollerer genaktivitet i dyrkede kødcellinjer. Specifikt hjælper H3K36me2 og H3K36me3 med at opretholde aktive forstærkere ved at blokere undertrykkende mærker som H3K27me2/3 fra at infiltrere genlegemer [9][10]. Når H3K36-methylering går tabt, forstyrres kromatinstrukturen, hvilket tillader undertrykkende mærker som H3K9me3 at invadere aktive regioner [9]."H3K36-methylering [er] en afgørende regulator af kromatintilstand og genomisk struktur." - Nature Communications [9]
Interaktionen mellem histonmærker er essentiel for at forbedre cellinjens ydeevne.Forskning antyder, at målretning af flere histonmodifikationer sammen ofte opnår bedre resultater end at fokusere på kun én [4].
Med disse fund i tankerne skal fremtidige studier udnytte præcise epigenetiske værktøjer for at sikre løbende forbedringer i præstationen af cellelinjer til dyrket kød.
Fremtidige Forskningsmuligheder
For at forbedre cellelinjens præstationer kræves innovative tilgange, såsom single-nucleus RNA-sekventering, for at kortlægge det epigenetiske landskab inden for forskellige cellesubpopulationer. Dette er særligt vigtigt for at identificere og forstå hvilende "reserve celler", der modstår differentiering. Disse celler, som udtrykker markører som PAX7 og NOTCH2 i stedet for at forpligte sig til myogen fusion, udgør en betydelig udfordring i produktionen af dyrket kød [3].
En anden lovende vej involverer udviklingen af kimære epigenetiske komplekser til præcis, ikke-genetisk kontrol. For eksempel demonstrerede forskere i 2025, at kombinationen af N-terminalen af SUZ12 med katalytiske domæner fra SUV39H2 eller SETD2 effektivt kunne erstatte H3K27me3 med H3K9me3 eller H3K36me3 ved adskillige gener [4] . Derudover kunne overvågning af H3K36me2 ved enhancere fungere som en kvalitetskontrolmarkør for at sikre cellelinjestabilitet [9].
Fremtidige bestræbelser bør fokusere på at opretholde H3K36-methylering på tværs af cellegenerationer. Dette ville hjælpe med at forhindre epigenetisk drift, hvilket gør det muligt for forskere og virksomheder som
Ofte stillede spørgsmål
Hvordan påvirker histonmærker muskeldifferentiering i dyrkede kødcellinjer?
Histonmærker er nøgleaktører i muskeldifferentiering, især for dyrkede kødcellinjer. For eksempel udløser reduktionen af H3K27me3 under differentiering myogene transkriptionsprogrammer, hvilket muliggør aktivering af gener, der er nødvendige for muskeludvikling. Finjustering af histonmodifikationer som H3K27me3 understøtter overgangen af cellinjer fra proliferation til dannelse af muskelvæv med specifikke karakteristika. Disse epigenetiske justeringer er essentielle for at fremme produktionen af dyrket kød.
Hvilke histonmodifikationer forudsiger bedst stabil, højtydende cellevækst i bioreaktorer?
H3K36-methylering skiller sig ud som en pålidelig markør for stabil, højtydende cellevækst i bioreaktorer.Denne ændring spiller en nøglerolle i at bevare celleidentitet og styre stamtræsprogrammer - begge er essentielle for at sikre konsekvent celleproliferation, især i produktionen af dyrket kød.
Kan epigenetisk redigering forbedre cellelinjer uden at ændre deres DNA-sekvens?
Epigenetisk redigering tilbyder en måde at forbedre cellelinjer uden at ændre deres DNA-sekvens. Ved at justere histonmærker og kromatinstruktur kontrollerer det genekspression. Forskning i histonmodifikationer fremhæver, hvordan disse ændringer kan påvirke celleidentitet og funktion. Denne tilgang har særligt potentiale til at forfine dyrkede kød cellelinjer.










