

Valinta naudan ja sian solulinjojen välillä on kriittinen päätös viljellyn lihan tuotannossa. Jokaisella solutyypillä on omat etunsa ja haasteensa, jotka vaikuttavat skaalautuvuuteen, kasvatusalustavaatimuksiin ja kykyyn luoda rakenteellisia lihatuotteita. Tässä on nopea yleiskatsaus:
- Naudan solulinjat soveltuvat hyvin lihaskudoksen tuotantoon, erityisesti pihvituotteisiin. Ne ovat erinomaisia marmoroitumisessa, mutta kohtaavat haasteita pitkäaikaisessa erilaistumisessa ja vaativat geneettisiä muutoksia skaalautuvuuden saavuttamiseksi.
- Sian solulinjat ovat ihanteellisia rasvantuotantoon, ja niillä on spontaani kuolemattomuus ja vakaa kasvu satojen jakautumisten ajan. Ne ovat kustannustehokkaita suurimittakaavaisessa tuotannossa, mutta saattavat vaatia tarkkaa ajoitusta yhteiserilaistumisessa lihassolujen kanssa.
Nopea vertailu
| Attribuutti | Naudan solulinjat | Sian solulinjat |
|---|---|---|
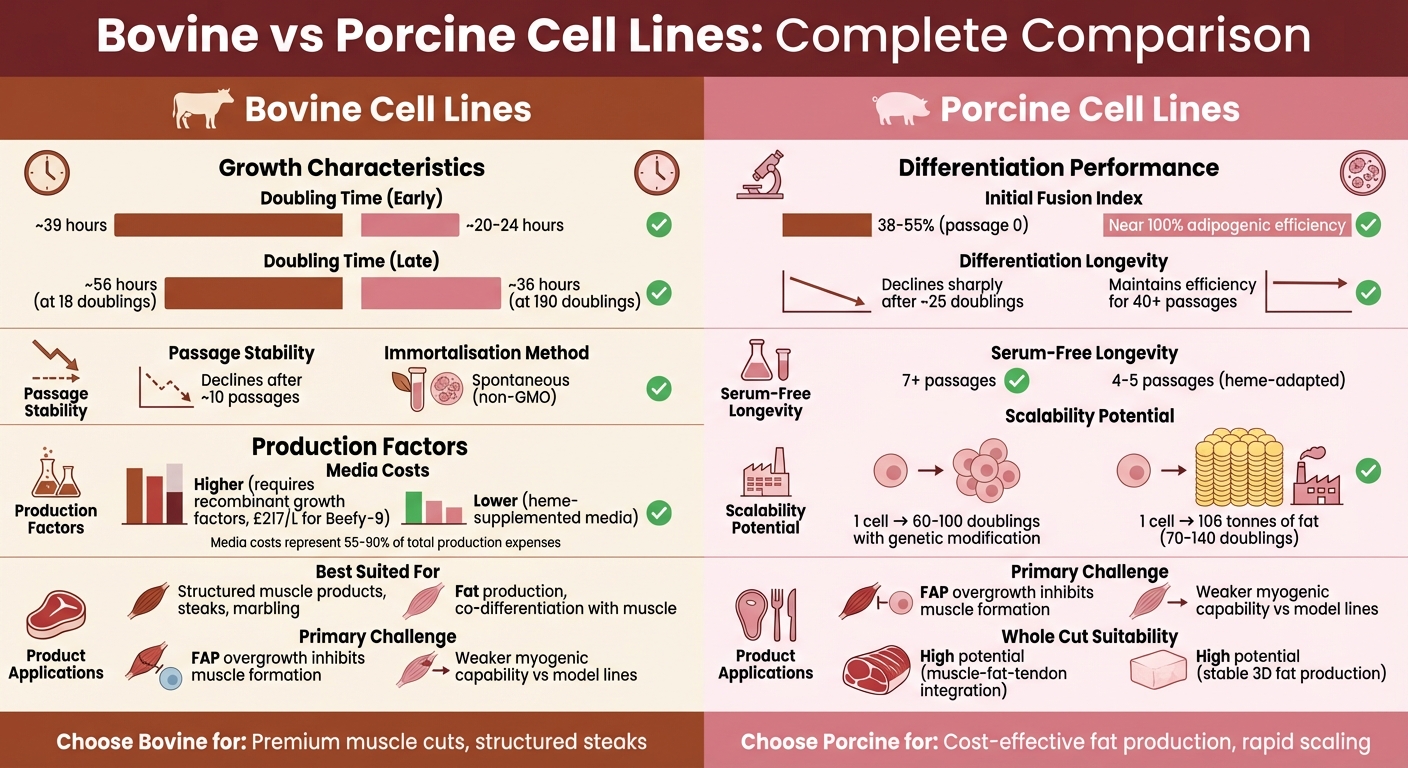
| Kaksinkertaistumisaika | ~39 tuntia (varhaiset passaasit) | 20–24 tuntia (varhaiset passaasit) |
| Kuolemattomuus | Vaatii geneettistä muokkausta | Spontaani |
| Differentoituminen | Vahva alussa, heikkenee ~25 kaksinkertaistumisen jälkeen | Vakaa adipogeeninen tehokkuus (>200 kaksinkertaistumista) |
| Median kustannukset | Korkeammat rekombinanttien kasvutekijöiden vuoksi | Alhaisemmat heemi-lisätyllä medialla |
| Rakenteellisen lihan soveltuvuus | Sopii marmoroitumiseen ja lihas-rasvaerotteluun | Tehokas rasva-lihas yhteisdifferentaatioon |
Molemmilla solulinjoilla on omat ainutlaatuiset vahvuutensa ja rajoituksensa, joten valinta riippuu tuotteen tavoitteista ja tuotantostrategioista.
Nauta- vs. sika-solulinjojen vertailu viljellyn lihan tuotannossa
Nauta-solulinjat
Sovellukset viljellyssä lihassa
Nauta-solulinjat soveltuvat erityisesti rakenteellisten lihatuotteiden, kuten pihvien ja muiden premium-leikkausten, tuottamiseen. Yksi niiden erottuvista ominaisuuksista on kyky kehittää aitoa marmoroitumista - lihaksensisäistä rasvaa, joka vastaa naudanlihan erottuvasta mausta ja koostumuksesta. Tämä marmoroituminen saavutetaan naudan satelliittisolujen (BSC:t), roolin kautta, jotka muodostavat lihaskomponentin, ja fibro-adipogeenisten progenitorien (FAP:t), kautta, jotka tuottavat rasvaa, jonka rasvahappoprofiili on lähes identtinen luonnollisen naudan ihonalaisen rasvan kanssa [2].
Oikean marmoroitumisen luominen vaatii huolellista koordinointia erilaistumisen aikana.Toisin kuin sian järjestelmät, jotka voivat samanaikaisesti erottaa lihaksen ja rasvan, naudan järjestelmät käsittelevät tyypillisesti myogeenisten (lihasmuodostavien) ja adipogeenisten (rasvamuodostavien) solujen erottelua erikseen. Nämä komponentit yhdistetään sitten tarkan rasva-lihas-suhteen hallitsemiseksi. Vaikka tämä menetelmä mahdollistaa suuremman räätälöinnin, se tuo myös lisämonimutkaisuutta tuotantoprosessiin [2].
Kasvutunnusmerkit
Vaikka naudan solut ovat tehokkaita sekä lihaksen että rasvan tuottamisessa, niiden kasvudynamiikka aiheuttaa haasteita laajamittaiselle tuotannolle. Keskeinen ongelma syntyy naudan satelliittisoluista, jotka menettävät kykynsä erilaistua niiden jatkaessa lisääntymistä. Esimerkiksi primaariset naudan myoblastit voivat käydä läpi 60–100 populaatiokaksinkertaistumista säilyttäen normaalin karyotyypin.Kuitenkin niiden kyky sulautua myotubeiksi - olennainen vaihe lihaskudoksen muodostumisessa - laskee merkittävästi noin 25 kaksinkertaistumisen jälkeen. Tämä rajoitus luo pullonkaulan tuotannon laajentamiselle, mikä vaatii noin 2,9×10¹¹ solua kilogrammaa märkämassaa kohden [7].
Toukokuussa 2023 Tufts University Centre for Cellular Agriculture -tutkijat ratkaisivat tämän ongelman kehittämällä geneettisesti kuolemattomia naudan satelliittisoluja (iBSC). Lisäämällä naudan telomeraasin käänteiskopioijaentsyymiä (TERT) ja sykliiniriippuvaista kinaasia 4 (CDK4), nämä solut pystyivät ylittämään 120 kaksinkertaistumista samalla kun ne muodostivat moniytimisiä myotubeja. Andrew J.Stout Tufts Universitystä korosti tämän läpimurron merkitystä:
"Jotta viljelty liha menestyisi laajassa mittakaavassa, elintarvikkeisiin liittyvien lajien lihassoluja on laajennettava in vitro nopeasti ja luotettavasti, jotta voidaan tuottaa vuosittain miljoonia metritonneja biomassaa." [5]
Kasvusuorituskykyyn vaikuttavat myös voimakkaasti tekijät, kuten kylvötiheys ja väliaineen koostumus. Esimerkiksi naudan rasvakudoksesta peräisin olevat kantasolut (bASCs) osoittivat optimaalista kasvua kylvötiheydellä 1 500 solua/cm², saavuttaen 28-kertaisen laajenemisen pyörivissä pulloissa käyttäen 80% väliaineen vaihtostrategiaa [1] . Lisäksi kemiallisesti määritellyt seerumittomat väliaineet ovat osoittaneet tukevansa naudan myoblastien eksponentiaalista kasvua nopeuksilla, jotka saavuttavat 97% perinteisiä seerumia sisältäviä väliaineita [6] . Tämä ei ainoastaan vähennä kustannuksia, vaan myös vastaa eettisiä näkökohtia, mikä tekee siitä lupaavan lähestymistavan tulevaisuuden tuotannolle.
Nämä naudalle ominaiset kasvupiirteet tarjoavat vankan perustan niiden vertaamiselle sian solulinjoihin viljellyn lihan tuotannon yhteydessä.
sbb-itb-ffee270
Sian Solulinjat
Sovellukset Viljellyssä Lihassa
Sian solulinjat ovat keskeisiä kypsien unilokulaaristen adiposyyttien tuottamisessa, jotka muistuttavat läheisesti luonnollista sianrasvaa[9].
Yksi merkittävä esimerkki on FaTTy-solulinja, joka on luotu spontaanin kuolemattomuuden kautta. Tämä solulinja osoittaa vaikuttavan ~100% adipogeenisen tehokkuuden yli 200 kaksinkertaistumisen ajan, tuottaen rasvahappoprofiileja, jotka vastaavat läheisesti alkuperäistä sianrasvaa. Tästä linjasta johdetut viljellyt adiposyytit voivat saavuttaa lipiditilavuuksia jopa 96,670 μm³.Kun FaTTy-tutkimusryhmä selittää:
"FaTTy on ainutlaatuinen karjasolulinja, jolla on erottuva adipogeeninen fenotyyppi, jolle on ominaista kyky erilaistua luotettavasti korkealla tehokkuudella erilaisissa viljelyolosuhteissa ja tuottaa kypsiä adiposyyttejä, joiden rasvahappoprofiilit ovat verrattavissa alkuperäiseen rasvaan." [9]
Toinen merkittävä solulinja, PK15H, menestyy korkeissa hemipitoisuuksissa, jopa 40 mM. Tämä ominaisuus auttaa jäljittelemään perinteisen sianlihan rikasta väriä ja rautapitoista makua[3]. Lisäksi viljeltyä sianrasvaa voidaan hienosäätää terveellisempien lipidikoostumusten saavuttamiseksi, jolloin monityydyttymättömien ja tyydyttyneiden rasvahappojen suhde on 3,2 verrattuna alkuperäiskudoksessa yleisesti esiintyvään 1,4 suhteeseen[9].
Kasvutunnusmerkit
Sikaperäiset solulinjat eivät ole vain taitavia rasvantuotannossa, vaan ne myös menestyvät kasvun ja skaalautuvuuden suhteen. Ne osoittavat vakaata ja nopeaa laajentumista, mikä tekee niistä erityisen sopivia laajamittaiseen tuotantoon. Esimerkiksi FaTTy-linjan populaation kaksinkertaistumisaika on aluksi 20–24 tuntia, mikä hidastuu vain hieman 22–36 tuntiin 140. ja 190. kaksinkertaistumisen välillä. Tämä johdonmukaisuus on merkittävä muutos, sillä yksi FaTTy-solu, joka on laajennettu 70:stä 140:een populaation kaksinkertaistumiseen, voisi teoriassa tuottaa 106 tonnia rasvaa 11 päivän erilaistumisjakson aikana[9] .
Yksi näiden solulinjojen merkittävä etu on niiden spontaani kuolemattomuus, mikä mahdollistaa pitkäaikaisen laajentumisen ilman geneettistä muuntelua. Tämä ei-GMO-status on sääntelyllinen voitto.Korostaen tätä, University of Ulsan College of Medicine totesi:
"Tutkimuksemme raportoi sian soluviljelmän, joka on viljeltävissä korkeassa heme-pitoisessa mediassa ja jota voidaan ylläpitää seerumittomissa olosuhteissa." [3]
Lisäksi sian lihas kantasolut osoittavat huomattavaa skaalautuvuutta, laajentumisnopeuksilla 10⁶ - 10⁷ kertaa, pystyvät tuottamaan 100 g - 1 kg viljeltyä lihaa[10]. Solujen lajittelutekniikoiden edistysaskeleet, käyttäen merkkejä kuten CD31, CD45, JAM1, ITGA5 ja ITGA7, ovat merkittävästi parantaneet korkean puhtauden lihas kantasolujen eristämistä. Nämä menetelmät tarjoavat 20% lisäyksen PAX7-positiivisuusasteissa verrattuna vanhempiin tekniikoihin[11] . Tämä parannus varmistaa, että myogeeninen potentiaali säilyy useiden passagien ajan, ratkaisten yleisen ongelman heikentyneestä erilaistumiskyvystä pitkäaikaisen laajentumisen aikana.
Nämä kasvu- ja erilaistumisedut tekevät sian soluista erinomaisen valinnan naudan soluihin verrattuna viljellyn lihan tuotannossa.
ICAN Webinaari solulinjoista ja viljellyn lihan sovellusten viljelyalustoista
Kasvunopeudet ja lisääntyminen verrattuna
Sukelletaanpa siihen, miten sian ja naudan solulinjat pärjäävät kasvun ja lisääntymisen suhteen. sian solulinjat, kuten spontaanisti kuolemattomaksi muuttunut FaTTy-linja, ovat huomattavasti nopeampia. Niiden alkuperäinen populaation kaksinkertaistumisaika on vain 20–24 tuntia [9]. Sen sijaan naudan satelliittisolut, vaikka niitä kasvatettaisiin optimoidussa seerumittomassa viljelyalustassa kuten Beefy-9, vievät noin 39 tuntia kaksinkertaistuakseen [12].
Eroavaisuudet tulevat vielä selvemmiksi useiden passagointien jälkeen.Ensisijaiset naudan satelliittisolut menettävät yleensä sekä lisääntymis- että erilaistumiskyvyn noin 10 passagen jälkeen [2]. Toisaalta FaTTy-sikalinja on säilyttänyt lähes 100% adipogeenisen tehokkuuden yli 200 populaation kaksinkertaistumisen ajan. Myöhemmissä vaiheissa niiden kaksinkertaistumisaika kasvaa vain hieman 22–36 tuntiin [9]. Tuftsin yliopiston toukokuussa 2022 tekemä tutkimus korosti, että Beefy-9:n naudansolut saavuttivat 18,2 populaation kaksinkertaistumista seitsemän passagen aikana (28 päivää) säilyttäen yli 96% Pax7⁺ kantasolukyvyn [12]. Samaan aikaan Edinburghin yliopiston tammikuussa 2025 julkaistu raportti vahvisti, että FaTTy-linja ylitti 200 kaksinkertaistumista menettämättä erilaistumispotentiaaliaan [9].
On myös selvä ero siinä, miten nämä solut saavuttavat kuolemattomuuden.Nisäkässolut tarvitsevat tyypillisesti geenimuuntelua - yleisesti TERT- ja CDK4-yli-ilmentämisen kautta - ylläpitääkseen pitkäaikaisen laajenemisen yli 120 kaksinkertaistumisen [5]. Vertailun vuoksi, porsassolut, kuten FaTTy-linja, saavuttavat spontaanin kuolemattomuuden ilman geneettistä muuntelua. Tämä tarjoaa selvän sääntelyedun, erityisesti markkinoilla, jotka suhtautuvat varauksella GMO:ihin [9].
Vertailutaulukko
| Ominaisuus | Naudan satelliittisolut | Sian MSC:t (FaTTy-linja) |
|---|---|---|
| Keskimääräinen kaksinkertaistumisaika | ~39 tuntia (optimoitu seerumiton) [12] | 20–24 tuntia (varhaiset passaasit) [9] |
| Myöhäisvaiheen kaksinkertaistumisaika | ~56 tuntia (18 kaksinkertaistumista) [12] | ~36 tuntia (190 kaksinkertaistumista) [9] |
| Passaasien stabiilisuus | Heikkenee ~10 passaasin jälkeen [2] | Vakaa >200 kaksinkertaistumista [9] |
| Kuolemattomuusmenetelmä | Engineered (TERT/CDK4) [2] | Spontaneous [9] |
| Stemness/Differentiation | >96% Pax7⁺ (jopa kulku 6) [12] | Lähellä 100% adipogeenista tehokkuutta [9] |
On syytä huomata, että in vivo satelliittisolut kaksinkertaistuvat noin 17 tunnissa, mikä korostaa vaikeutta saavuttaa luonnollisia kasvunopeuksia in vitro [12].
Median vaatimukset ja erottelutehokkuus
Median riippuvuus verrattuna
Median kustannukset voivat hallita viljellyn lihan tuotantoa, usein muodostaen 55% - 90% kuluista, ja joissakin järjestelmissä jopa ylittäen 99% [3] [12] .
Nautasoluille yleinen vaatimus on 20% sikiönaudan seerumi, joka voi olla merkittävä mediakustannus [12]. Seerumiton vaihtoehto, Beefy-9, käyttää B8-perusväliainetta yhdistettynä rekombinanttiin ihmisen albumiiniin. Kustannukset vaihtelevat hankintalähteen ja tilausmäärän mukaan, joten on parasta tarkistaa toimittajalta tai tuotesivulta ajankohtaiset hintatiedot [12] . Korkeat albumiinitasot seerumittomassa mediassa voivat kuitenkin haitata solujen kiinnittymistä, joten rekombinantti albumiini lisätään tyypillisesti 24 tuntia passauksen jälkeen [12].
Sikojen solulinjat lähestyvät seerumittomaan sopeutumiseen eri tavalla. PK15-solut käyttävät esimerkiksi bakteerien hemeuutteita Corynebacterium [3]. Heme ei ainoastaan vähennä seerumin riippuvuutta, vaan myös parantaa makua ja väriä. Kuitenkin pitoisuudet yli 10 mM voivat olla myrkyllisiä, vaikka sikasolut sietävät jopa 40 mM detoksifikaatiogeenejä ylireguloimalla [3] . Huolimatta tästä sietokyvystä, hemeillä täydennetyssä väliaineessa kasvatetut sikasolut pysyvät yleensä elinkelpoisina vain 4–5 passagen ajan, kun taas Beefy-9:ssä viljellyt naudansolut voivat ylläpitää kasvua seitsemän tai useamman passagen ajan [3][12].
Molemmat solutyypit ovat voimakkaasti riippuvaisia fibroblastien kasvutekijästä-2 (FGF-2).Nisäkässolut, kuten naudan solut, voivat ylläpitää lyhytaikaista kasvua, vaikka FGF-2-tasot vähennetään 40 ng/mL:sta 5 ng/mL:aan [12]. Lisäksi matalan glukoosipitoisuuden omaavan kasvatusalustan (1 g/L) käyttö auttaa säilyttämään kantasolumerkit naudan soluissa [13].
Nämä erityiset kasvatusalustavaatimukset ovat kriittisiä tuotannon laajentamisessa ja vaikuttavat suoraan erilaistumistehokkuuteen.
Erilaistumistehokkuus
Vaikka kasvatusalustojen kustannukset ovat merkittävä tekijä, erilaistumistehokkuus vaikuttaa myös merkittävästi viljellyn lihan skaalautuvuuteen.
Naudan solut kohtaavat haasteita erilaistumistehokkuudessa niiden laajentuessa. Esimerkiksi Belgian Blue -karjan naudan myoblastit saavuttavat aluksi fuusioindeksin noin 55% 14 populaatiokaksinkertaistumisen jälkeen, mutta tämä laskee jyrkästi alle 10% 25 kaksinkertaistumisen jälkeen [7]. Samoin, sikiöperäiset naudansolut aloittavat korkeammilla fuusioindekseillä (noin 54.6%) verrattuna aikuisperäisiin soluihin (noin 38.0%), mutta molemmat kokevat erilaistumiskyvyn heikkenemistä noin 6.81% per passaus [7].
Sikojen solut puolestaan osoittavat vakaampaa suorituskykyä. Kuolemattomaksi tehdyn ISP-4 sikojen preadiposyyttikannan adipogeeninen erilaistumistehokkuus säilyy korkeana yli 40 passauksen ajan, saavuttaen 100-kertaisen lipidikertymän 8 päivän erilaistumisprotokollan aikana [8]. Tämä tekee sikojen soluista erityisen houkuttelevia rasvantuotantoon, kun taas naudansolut soveltuvat paremmin lihaserilaistumiseen varhaisissa passauksissa, mutta niiden pitkäaikainen ylläpito on haastavaa.
| Ominaisuus | Naudan satelliittisolut | Sikojen solulinjat |
|---|---|---|
| Alkuperäinen fuusioindeksi | 38–55% (passage 0) [7] | Ei määritelty lihakselle |
| Differentiation Longevity | Vähenee jyrkästi noin 25 kaksinkertaistumisen jälkeen [7] | Säilyttää tehokkuuden yli 40 passagen ajan (ISP-4 adipogeeninen) [8] |
| Seerumiton pitkäikäisyys | Ylläpitää kasvua yli 7 passagen ajan [12] | Käyttökelpoinen 4–5 passagea (heme-adaptoitu) [3] |
| Keskeiset lisäravinteet | Rekombinantti albumiini, FGF-2 [12] | Hemiuute, insuliini, deksametasoni [3][8] |
| Lipidituotanto | Minimaalinen (lihaskeskeinen) | 100-kertainen kasvu (ISP-4) [8] |
Soveltuvuus rakenteellisille lihatuotteille
Solulinjojen valinta on keskeisessä asemassa muokkaamassa paitsi kasvua ja mediaympäristöjä, myös viljellyn lihan tuotteiden rakennetta.Kun pyritään jäljittelemään pihvin tai porsaanleikkeen rakennetta ja ulkonäköä, rasva- ja lihassolujen oikea suhde on olennaista.
Rasva-Lihas Yhteisdifferentaatio
Naudan ja sian solulinjat käyttäytyvät eri tavoin yhteisdifferentaatiossa. Naudan soluviljelmät kohtaavat usein haasteita, kuten FAP (fibro-adipogeeninen progenitori) liikakasvua, joka häiritsee lihasten kehitystä alentamalla fuusioindeksiä. Lisäksi näiden viljelmien adiposyytit vapauttavat signaaleja, kuten myostatiinia ja IL-6:ta, jotka estävät myogeniinin ilmentymistä, pysäyttäen tehokkaasti lihassyiden muodostumisen[16].
Tämän ratkaisemiseksi Mosa Meat-tutkijat loivat optimoidun seerumittoman kasvualustan (i-SFGM). Tämä alusta sisältää trijodityroniinia (T3) ja lisää HGF:ää samalla kun se sulkee pois PDGF-BB:n FAP:n liikakasvun hallitsemiseksi.He käyttävät myös modulaarisia adiposfäärejä (200–400 µm) pitääkseen rasva- ja lihassolut fyysisesti erillään varhaisissa kasvuvaiheissa[4][14].
Sikojen solulinjat puolestaan osoittavat koordinoidumman lähestymistavan yhteisdifferentaatioon. ISP-4 preadiposyyttikanta toimii esimerkiksi hyvin sikojen lihassatelliittisolujen kanssa, tuottaen marmoroitumista, joka muistuttaa perinteistä lihaa. Tämä prosessi sisältää 48 tunnin adipogeenisen induktiovaiheen, jota seuraa 96 tuntia 2% hevosen seerumissa myogeneesin käynnistämiseksi. Tämä johtaa kypsiin lihassäikeisiin, jotka ovat kietoutuneet adiposyyttien kanssa[8]. Kuitenkin sikojen lihassatelliittisolut ovat yleensä heikompia myogeenisissä kyvyissään verrattuna standardimalleihin, kuten C2C12, mikä vaatii tarkkaa ajoitusta, jotta adiposyytit eivät hallitse viljelmää[8].
Nämä erot erilaistumisessa korostavat ainutlaatuisia haasteita ja mahdollisuuksia, joita jokainen solutyyppi esittää tuotannon laajentamiselle.
Skaalautuvuus ja tuotantohaasteet
Rakenteellisen viljellyn lihan tuotannon skaalaaminen vaatii johdonmukaista solusuorituskykyä. Sian solulinjat ovat yleensä skaalautuvampia. Esimerkiksi spontaanisti kuolemattomaksi muuttunut FaTTy-linja säilyttää lähes 100% adipogeenisen tehokkuuden yli 200 populaatiokaksinkertaistumisen ajan[9]. Sian solulinjan laajentaminen 70:stä 140 kaksinkertaistumiseen voisi teoriassa tuottaa jopa 106 tonnia rasvaa[9]. Lisäksi ISP-4-kanta on osoittanut 40-kertaisen solutiheyden kasvun kuuden päivän aikana, kun sitä kasvatetaan mikrokantajilla pyörivässä pullotusjärjestelmässä [8].
"FaTTy on ainutlaatuinen karjasolulinja, jolla on erottuva adipogeeninen fenotyyppi...nämä ominaisuudet yhdessä sen ei-GMO-luonteen kanssa tekevät FaTTysta erittäin lupaavan perustyökalun." – Nature Food, 2025[9]
Naudan solulinjat kohtaavat enemmän esteitä. FAP-kontaminaatio heikentää niiden kykyä erilaistua tehokkaasti lihaskudokseksi[4]. Lisäksi kasvutekijöiden, kuten FGF-2:n ja TGF-β:n, korkea hinta - usein yli 90% media kuluista - tekee naudan solulinjojen skaalaamisesta kalliimpaa[17]. Nämä solut vaativat myös erikoispinnoitteita, kuten Laminin-521, satelliittisolujen kiinnittymisen edistämiseksi ja FAP-häiriöiden minimoimiseksi[4].
Yhden tonnin viljellyn lihan tuottaminen sisältää noin 10¹³ solua, ja rakenteelliset tuotteet, kuten kokonaiset leikkaukset, vaativat kehittyneitä tuotantojärjestelmiä, kuten perfuusio- tai pakattujen sänkyjen reaktoreita, tukemaan 3D-tukirakenteita ja biomateriaaleja, joita tarvitaan niiden kehittämiseen[15].
Vertailutaulukko
| Ominaisuus | Naudan solulinjat | Sian solulinjat |
|---|---|---|
| Pääasiallinen skaalautuvuushaaste | FAP-liikakasvu lihaskulttuureissa[4] | Sovittaminen suspensio/seerumittomaan viljelyyn[9] |
| Differentiation vakaus | Heikkenee noin 10 passagen jälkeen[2] | Kannat kuten FaTTy vakaita >200 kaksinkertaistumiseen asti[9] |
| Yhteisdifferentaatio | Adiposyytit estävät myogeneesiä[16] | Onnistuneet marmoroitumisprototyypit saavutettu[2][8] |
| Rakenteellinen lujuus | Korkea; kykenee lihas-rasva-jänne-integraatioon[14] | Kohtalainen; keskittyy kuitujen kohdistettuun kasvuun[14] |
| Kokonaisleikkauksen soveltuvuus | Korkea potentiaali, rajoitettu FAP-häiriön vuoksi[4] | Korkea potentiaali vakaan 3D-rasvatuotannon ansiosta[9] |
| Tekstuurin haaste | Vähentynyt yhtenäisyys kypsennyksen jälkeen[14] | Taipumus olla pehmeämpää kuin kaupallinen sianliha[14] |
Päätelmä
Päätös naudan ja sian solulinjojen välillä edellyttää niiden erilaisten hyötyjen ja haasteiden tasapainottamista viljellyn lihan tuotannossa.Naudan satelliittisolut ovat suora reitti luustolihaskudoksen luomiseen ja hyötyvät olemassa olevista seerumittomista väliainekoostumuksista, kuten Beefy-9 [2]. Toisaalta sian solulinjoja on jo käytetty viljellyn sianlihan prototyyppien kehittämiseen, ja ne osoittavat lupausta satelliittisolujen kanssa yhteisdifferentioitumisessa marmoroitujen liharakenteiden luomiseksi [2].
Skaalautuvuus on edelleen merkittävä este. Väliainekustannukset ja bioreaktorin skaalautuvuus muodostavat 55%–90% kokonaisvalmistuskustannuksista, ja optimoitujen solulinjojen saatavuus on edelleen rajallista, mikä hidastaa kaupallista edistymistä [3][2] .
"Viljellyn lihan tuotannossa käytetyt solulinjat määrittävät lopulta monia alavirran muuttujia, jotka on otettava huomioon." – GFI [2]
UKK:t39022>
Mikä solulinja on paras kokonaisille tuotteille, kuten pihveille tai kyljyksille?
Lihaksen progenitorisoluista, kuten satelliittisoluista, johdetut solulinjat ovat usein ihanteellisia tuottamaan kokonaisia tuotteita, kuten pihvejä tai kyljyksiä. Nämä solut voivat kehittyä kypsäksi lihaskudokseksi, luoden rakenteellisen koostumuksen ja muodon, joita tarvitaan tämän tyyppisille tuotteille.
Kuinka valitsen geneettisen ja spontaanin kuolemattomuuden välillä?
Solujen kuolemattomuuden valinta viljellyn lihan tuotannossa riippuu prioriteeteistasi, mukaan lukien turvallisuus, skaalautuvuus ja sääntelyyn liittyvät näkökohdat.
Geneettinen kuolemattomuus sisältää tiettyjen geenien, kuten telomeraasin, lisäämisen solujen jakautumiskyvyn tarkkaan hallintaan.Vaikka tämä menetelmä tarjoaa ennustettavuutta ja johdonmukaisuutta, se saattaa herättää huolta geneettisestä muuntelusta ja mahdollisista riskeistä, kuten kasvainten muodostumisesta.
Toisaalta, spontaani immortalisaatio tapahtuu luonnollisesti ajan myötä pitkäaikaisissa soluviljelmissä. Tämä lähestymistapa välttää geenitekniikan, mikä voisi helpottaa sääntelyhyväksyntää ja lisätä hyväksyntää kuluttajien keskuudessa, jotka ovat varovaisia geneettisen muuntelun suhteen.
Molemmilla menetelmillä on omat vahvuutensa ja haasteensa, tarjoten erilaisia polkuja kohti skaalautuvaa viljeltyä lihantuotantoa. Valinta riippuu lopulta kontrollin, sääntelyesteiden ja kuluttajien luottamuksen tasapainottamisesta.
Mikä on suurin kustannustekijä naudan- ja sianlihansolujen viljelyalustassa?
Suurin kustannus naudan- ja sianlihansolujen viljelyalustan tuotannossa johtuu sen komponenttien hinnasta ja monimutkaisuudesta.Medioiden kehittäminen ja hienosäätö on merkittävä haaste, erityisesti koska media muodostaa vähintään 50% muuttuvista käyttökustannuksista. Lisäksi kullekin lajille räätälöidyt säädöt lisäävät monimutkaisuutta. Nämä tekijät vaikuttavat merkittävästi viljellyn lihan kokonaiskustannuksiin.









