

Ribosomitekniikka muokkaa viljellyn lihan tuotantoa parantamalla proteiinisynteesiä solutasolla. Ribosomit, solun proteiinitehtaat, ovat kriittisiä aktiinin, myosiinin ja muiden proteiinien tuottamisessa, jotka määrittävät lihan rakenteen ja ravintoarvon. Vakiintuneet solulinjat eivät kuitenkaan ole optimoituja suurimittakaavaiseen lihantuotantoon tarvittavaa korkeaa tuottavuutta varten.
Keskeisiä edistysaskeleita ovat:
- Optimoidut ribosomaaliset RNA-variantit: 1,7 × 10⁷ varianttia sisältävien kirjastojen seulonta on osoittanut potentiaalia lisääntyneelle translaatioaktiivisuudelle.
- Ortogonaaliset ribosomit: Nämä suunnitellut ribosomit erikoistuvat tuottamaan tiettyjä proteiineja, kuten myosiinia, häiritsemättä solun normaaleja toimintoja.
- Kodonin optimointi: mRNA-sekvenssien räätälöinti ribosomien mieltymyksiin on tuottanut jopa 72-kertaisen proteiini-ilmentymän.<|vq_15392|>
- Myokiini signalointi: Proteiinit kuten IL-15 ja myonektiini parantavat ribosomien biogeneesiä ja proteiinisynteesiä lihasten erilaistumisen aikana.
Haasteita on edelleen energian tarpeiden tasapainottamisessa, solujen vakauden ylläpitämisessä ja tuotannon skaalaamisessa teolliselle tasolle. Esimerkiksi ribosomien yliaktiivisuus voi johtaa väärin laskostuneisiin proteiineihin tai aineenvaihdunnan rasitukseen, kun taas ravinteiden diffuusion rajat bioreaktoreissa rajoittavat kudoksen kasvua yli 200 μm. Näiden ongelmien ratkaiseminen vaatii ribosomien suunnittelun integroimista kehittyneisiin bioprosessointistrategioihin.
Tämä artikkeli tutkii, kuinka nämä menetelmät muokkaavat viljellyn lihan tulevaisuutta ja mitä esteitä on voitettava kaupallisen elinkelpoisuuden saavuttamiseksi.
Ribosomit ja proteiinibiosynteesi: Johdanto
Ribosomien rakenne ja toiminta nisäkässoluissa
Ribosomit ovat proteiinisynteesin ytimessä, kääntäen mRNA-sekvenssejä toiminnallisiksi proteiineiksi.Imettäväisten soluissa ribosomit luokitellaan 80S-partikkeleiksi, jotka koostuvat kahdesta alayksiköstä: 40S pienestä alayksiköstä, joka dekoodaa mRNA:ta, ja 60S suuresta alayksiköstä, joka vastaa peptidisidoksen muodostamisen katalysoinnista. Käännösprosessi sisältää kolme päävaihetta: aloitus, jossa aloituskodoni tunnistetaan; pidentyminen, jossa aminohapot lisätään peräkkäin kasvavaan polypeptidiketjuun; ja lopetus, joka tapahtuu, kun lopetuskodoni saavutetaan.
Kaksi tiettyä aluetta suuressa alayksikössä ovat erityisen tärkeitä insinöörisovelluksille: peptidylitransferaasikeskus (PTC), joka helpottaa peptidisidoksen muodostumista, ja poistumistunneli, jonka kautta vastasyntetisoitu polypeptidi poistuu [3].
Näiden ydintoimintojen ymmärtäminen on olennaista tutkiessa, kuinka ribosomin suorituskykyä voidaan optimoida parantamaan viljellyn lihan tuotantoa.
Miksi proteiinisynteesi on tärkeää viljellylle lihalle
Proteiinisynteesin tehokkuus on kriittinen tekijä viljellyn lihan kehityksessä, erityisesti in vitro myogeneesin aikana. Tämä prosessi muuntaa lihaksen satelliittisolut (MSC:t) monitumaisiksi myofibrilleiksi, jotka ovat rikkaita supistuvissa proteiineissa, kuten aktiinissa ja myosiinissa. Ribosomit ovat keskeisessä roolissa tässä muutoksessa [4].
"noin kahdeksan biljoonaa lihassolua tarvitaan tuottamaan 1 kg proteiinia perinteisestä bioreaktorista, jonka kapasiteetti on 5 000 L" [5]
Tämä hämmästyttävä vaatimus korostaa, kuinka jopa pienet parannukset ribosomien tehokkuudessa voivat merkittävästi lisätä tuotantotuottoja, mikä vaikuttaa suoraan viljellyn lihan kaupalliseen toteutettavuuteen.
Kun solut kypsyvät, niiden ribosomaalinen aktiivisuus muuttuu. Proliferaatiovaiheessa MSC:t priorisoivat nopeaa jakautumista. Kuitenkin, kolmen tai viiden päivän kuluttua erilaistumisesta, painopiste siirtyy aikuisten isomuotojen synteesiin supistuvista proteiineista ja solujen fuusioitumiseen myotubeiksi [4]. Tätä siirtymää säätelevät tietyt signaalimolekyylit, tai myokiinit.
Esimerkiksi, Interleukiini‑15 (IL‑15) edistää Myosiini Raskasketjun (MyHC) proteiinin kertymistä samalla vähentäen proteiinien hajoamista, toimien keskeisenä anabolisena tekijänä lihasten kehityksessä [4]. Samoin, Myonektiini tukee lihasten kasvua lisäämällä proteiinisynteesiä PI3K/Akt/mTOR signaalireitin kautta [4]. Ymmärtäminen, miten nämä signaalireitit vaikuttavat ribosomien aktiivisuuteen, on olennaista suunniteltaessa skaalautuvia solulinjoja, jotka täyttävät tuotantovaatimukset.Nämä oivallukset luovat pohjan seuraavissa osioissa käsitellyille insinööristrategioille.
Nykyinen tutkimus ribosomien suunnittelusta
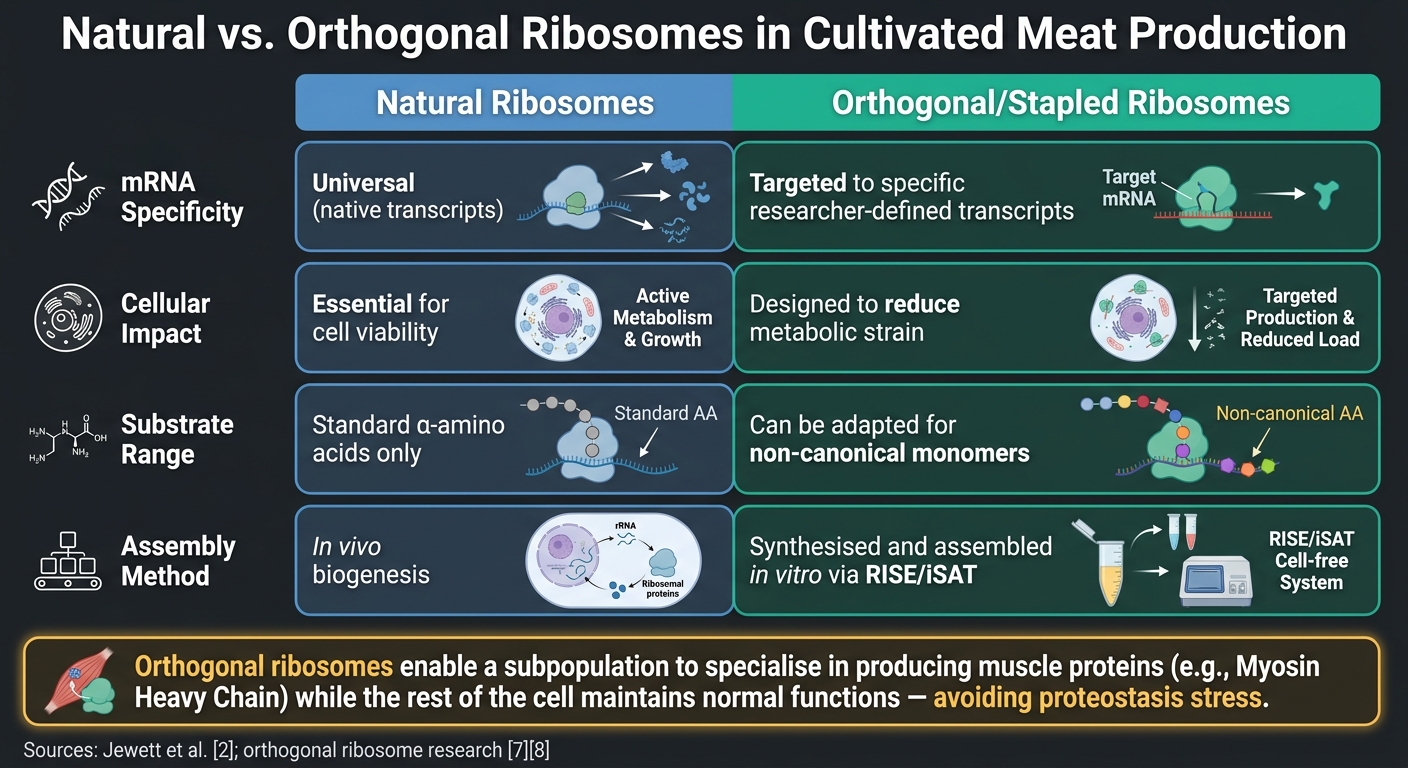
Luonnolliset vs. ortogonaaliset ribosomit viljellyn lihan tuotannossa
Ribosomien biogeneesi ja käännöksen hallinta
Ribosomien biogeneesi, prosessi, jossa solut rakentavat uusia ribosomeja, on erittäin säädelty ja energiaintensiivinen toiminta. Nisäkässoluissa se edustaa suurta osaa solun aineenvaihdunnan tuotannosta. Pelkkä käännös voi kuluttaa jopa 75% solun koko energiabudjetista [8], tehden siitä yhden eniten resursseja vaativista soluprosesseista.
Kun ribosomien jakaminen on tehotonta - esimerkiksi silloin, kun ribosomit pysähtyvät varhaisilla koodausalueilla - se luo pullonkauloja, jotka vähentävät vapaiden ribosomien saatavuutta ja lopulta rajoittavat proteiinituotantoa.Laskennalliset mallit ovat osoittaneet, että näiden pullonkaulojen ratkaiseminen muokkaamalla vain 100 geeniä voisi parantaa ribosomien allokointia 35% hiivassa ( Saccharomyces cerevisiae) ja 57% Escherichia coli [8]. Näillä löydöksillä on suoria vaikutuksia ribosomidynamiikan optimointiin nisäkässoluissa, erityisesti viljellyn lihan teollisuudessa, jossa energiatehokkuus ja proteiinituotos ovat kriittisiä.
Ribosomien muokkaus viljellyn lihan konteksteissa
Edistysaskeleet ribosomien muokkauksessa sovelletaan nyt viljellyn lihan tuotantoon, hyödyntäen ribosomien biogeneesin perustietoa. Jopa tutkimukset, joita ei ole suoraan tehty lihassoluissa, tuottavat oivalluksia, jotka ovat merkityksellisiä viljellyn lihan solulinjoille.
Joulukuussa 2020 Hadas Zur ja Tamir Tuller Tel Avivin yliopistosta osoittivat Ribosome Traffic Engineering (RTE) -menetelmän potentiaalin parantaa kasvunopeuksia ja proteiinituotantoa. Käyttäen CRISPR-Cas9 , he esittelivät synonyymimutaatioita ramp-alueella (kodonit 11–50) RPO21 ja CYS4 S. cerevisiae . Kaksoismutantti osoitti parantunutta log-vaiheen kasvua ja solutiheyttä. Tutkijat kuitenkin varoittivat, että käännösoptimoinnin ja kasvunopeuden välinen suhde heikkenee dioksisessa siirtymässä ja stationaarivaiheissa, joissa muut tekijät kuin käännös tulevat nopeutta rajoittaviksi [8]. Tämä oivallus on erityisen merkityksellinen suunniteltaessa erilaistumisprotokollia viljellyn lihan tuotannossa.
Helmikuussa 2020 Michael Jewettin tiimi Northwestern University:ssa vahvisti RISE (Ribosome In vitro Synthesis and Evolution) -menetelmän. Tämä tekniikka sisältää noin 1.7 × 10⁷ ribosomaalisen RNA:n variantin kirjaston seulonnan. [2]. Toimimalla täysin elävien solujen ulkopuolella, RISE ohittaa tappavien ribosomimutaatioiden asettamat rajoitukset, joita ei voida tutkia in vivo.
"In vitro -lähestymistapa voittaa solujen elinkelpoisuusrajoitukset, mahdollistaen tappavien ribosomimutaatioiden tutkimisen." - Michael Jewett et al. [2]
Toinen lupaava innovaatio viljellylle lihalle on ortogonaalisten ribosomien. käyttö. Nämä suunnitellut ribosomi–mRNA-parit toimivat itsenäisesti solun alkuperäisestä käännöskoneistosta.Tämä mahdollistaa tutkijoiden kohdistaa ribosomaalista aktiivisuutta tiettyihin kohteisiin, kuten myosiinin raskasketjun (MyHC) isomuotoihin, jotka ovat kriittisiä lihasten rakenteelle, häiritsemättä olennaisia soluprosesseja [6]. Vertailututkimukset korostavat ortogonaalisten ribosomien etuja luonnollisiin verrattuna:
| Ominaisuus | Luonnolliset ribosomit | Ortogonaaliset/Nidotut ribosomit |
|---|---|---|
| mRNA-spesifisyys | Universaali (luonnolliset transkriptit) | Kohdistettu tiettyihin tutkijan määrittelemiin transkripteihin [6] |
| Soluvaikutus | Välttämätön elinkelpoisuudelle | Suunniteltu vähentämään aineenvaihdunnan rasitusta [7] |
| Substraattivalikoima | Standardit α-aminohapot | Voidaan mukauttaa ei-kanonisiin monomeereihin [7] |
| Kokoonpano | In vivo biogeneesi | Synteettisesti ja koottu in vitro RISE/iSAT [2] |
Tärkein huomioitava seikka tässä on, että ortogonaaliset ribosomit mahdollistavat ribosomien alipopulaation erikoistumisen lihasproteiinien, kuten MyHC:n, tuottamiseen samalla kun solun muut toiminnot säilyvät normaaleina.Tämä välttää proteostaasin stressin riskin, joka voi syntyä, kun koko käännösjärjestelmä pakotetaan ylituottamaan tiettyjä proteiineja.
Strategiat ribosomien suorituskyvyn parantamiseksi
Ribosomien biogeneesin lisääminen
Ribosomien määrän lisääminen on suora tapa tehostaa proteiinituotantoa, ja kaksi päämenetelmää on saanut huomiota. Ensimmäinen sisältää ribosomaalisen RNA:n (rRNA) geenien epigeneettisen tilan muokkaamisen niiden käännöskapasiteetin lisäämiseksi.
"Ribosomaalisen RNA:n geenien epigeneettinen muokkaus parantaa proteiinituotantoa." - Santoro R., Lienemann P., Fussenegger M. [1]
Toinen lähestymistapa hyödyntää PI3K/Akt/mTOR-signalointireittiä. Myokiinit kuten IL-15, myonektiini ja irisiini aktivoivat tämän reitin, mikä edistää ribosomien biogeneesiä myotubusten kypsymisen aikana, kuten aiemmin käsiteltiin.
Kuitenkin, tämä ribosomien tuotannon kasvu on tasapainotettava huolellisesti solun aineenvaihduntakyvyn kanssa, sillä ribosomien synteesi on yksi eniten energiaa vaativista prosesseista elävissä soluissa [1].
Kun ribosomien määrä on kasvanut, keskittyminen siirtyy varmistamaan, että ne ovat täysin mukana translaatiossa.
Translaation aloituksen ja pidentämisen parantaminen
Kaikkien ribosomien toiminnan maksimointi on olennaista, sillä jopa kasvun optimoiduissa soluissa 15–20% ribosomeista pysyy passiivisena [9] . Tämä edustaa merkittävää käyttämätöntä kapasiteettivaraa viljellyissä lihasolulinjoissa.
Translaation pidentämisnopeus riippuu kahdesta tekijästä: ribosomin luontaisesta nopeudesta ja aktiivisesti translaatiossa mukana olevien ribosomien osuudesta [9]. Aminohappotasojen ylläpitäminen viljelyalustassa on kriittistä niiden optimoimiseksi. Lisäksi solulinjojen muokkaaminen ribosomiproteiinien stabiloimiseksi auttaa suojaamaan rRNA:ta väärin laskostumiselta ja hajoamiselta, mikä vähentää tyypillistä 10% rRNA:n menetystä huippukasvuolosuhteissa [9] .
Kun ribosomiaktiivisuus on maksimoitu, mRNA-sekvenssien hienosäätö on seuraava askel proteiinisynteesin nopeuttamiseksi edelleen.
mRNA-optimointi ja kodonien käyttö
Ribosomien suorituskyky riippuu suuresti niiden käsittelemän mRNA:n laadusta. Kodonien optimointi räätälöi kohdeproteiinien koodaavat sekvenssit vastaamaan isäntälajin - kuten naudan, sian tai kalan - tRNA-poolia. Tämä kohdistaminen estää ribosomien pysähtymisen pidentymisen aikana ja lisää läpimenoa kriittisille myogeenisille proteiineille, kuten MyoD ja Myf5.
Kodonin optimoinnin lisäksi transkriptionaalinen viritys varmistaa oikean tasapainon rRNA:n ja mRNA:n välillä solussa. Näiden komponenttien välinen epäsuhta voi luoda pullonkauloja, mikä vähentää kokonaistehokkuutta [1].
Käytännön sovelluksissa Integrated Synthesis, Assembly, and Translation (iSAT) -järjestelmät tarjoavat arvokkaan työkalun. Nämä järjestelmät käyttävät soluvapaita uutteita ja fluoresenssipohjaisia testejä prototyyppien optimoitujen mRNA:iden in vitro -testaukseen ennen niiden integrointia stabiileihin solulinjoihin. Tämä iteratiivinen lähestymistapa mahdollistaa tutkijoiden vertailla nopeasti kodonin optimoituja variantteja, parantaen olennaisten myogeenisten proteiinien tuottoa ja vahvistaen viljellyn lihan tuotannon skaalautuvuutta [1].
Kompromissit: Kasvu, Erottelu ja Tuotelaatu
Ribosomien suorituskyvyn optimointi vaatii hienovaraista tasapainoa proteiinisynteesin tehostamisen ja solujen kasvun ja erottelun vaikutusten hallinnan välillä, kuten aiemmin on esitetty.
Metabolinen Kuormitus ja Proteostaasin Stressi
Ribosomien muokkaaminen proteiinituotannon tehostamiseksi lisää energiantarvetta, koska se ohjaa ATP:tä ja aminohappoja pois muista elintärkeistä solutoiminnoista. Ribosomisynteesi on jo yksi solun energiaintensiivisimmistä prosesseista, ja sen lisävahvistaminen voi pahentaa näitä energiavaatimuksia.
Tämä tehostunut toiminta voi myös vaikuttaa proteiinien laatuun. Yliaktiiviset ribosomit voivat ylikuormittaa solun kaperoneja, mikä johtaa väärin laskostuneisiin proteiineihin ja avaa laskostumattomien proteiinien vasteen (UPR). Tällainen stressi voi estää kasvua tai jopa johtaa solukuolemaan.Ensisijaisille aikuisille kantasoluille karjaeläinlajeista, kuten naudoista tai lampaista, joilla on luonnostaan rajallinen lisääntymiskyky, nämä lisästressit voivat merkittävästi vähentää elinkelpoisten solujakautumisten määrää ennen kuin solujen vanheneminen alkaa [5].
Viljellyn lihan tuotannossa kudoksen paksuus ylittää harvoin 200 μm ravinteiden diffuusion rajoitusten vuoksi, mikä voi johtaa solukuolemaan suurempien kudosaggregaatioiden ytimessä [5]. Strategiat, jotka lisäävät energiankulutusta, voivat kiihdyttää ravinteiden ehtymistä näillä kriittisillä alueilla, joissa johdonmukainen proteiinisynteesi on välttämätöntä. Lisäksi lisääntynyt aineenvaihdunnan rasitus voi häiritä lihasten erilaistumiseen tarvittavia hienosäädettyjä signaalireittejä.Vaikutukset lihasten erilaistumiseen ja proteiinikoostumukseen
Ribosomitekniikan aiheuttamat stressit voivat ulottua aineenvaihdunnan ulkopuolelle, mahdollisesti häiritsemällä lihasten kehitystä. Myogeneesi, lihasten muodostumisprosessi, perustuu tarkasti säädeltyyn transkriptiotekijöiden sarjaan: Pax7 varmistaa, että kantasolut pysyvät lepotilassa, Myf5 edistää myoblastien lisääntymistä, ja MyoD käynnistää erilaistumisen [5] . Proteiinisynteesin muuttaminen voisi häiritä tätä sarjaa, pysäyttää erilaistumisen tai tuottaa epätavallisia lihaskuitukoostumuksia. Tämä saattaa johtaa vähempiin lihaksensisäisiin rasvakerrostumiin, jotka ovat avainasemassa halutun tekstuurin ja maun saavuttamisessa viljellyssä lihassa [5].
Tämän seurauksena on olennaista ylläpitää tiukkaa laadunvalvontaa seuraamalla myogeenisten merkkiaineiden ilmentymistä koko suunnitteluprosessin ajan, jotta varmistetaan asianmukainen lihasten kehitys ja tuotteen laatu.
sbb-itb-ffee270
Tutkimusaukot ja tulevaisuuden suuntaviivat
Ribosomien suunnittelun edistysaskeleet näyttävät lupaavilta, mutta niiden soveltaminen kaupalliseen viljellyn lihan tuotantoon kohtaa edelleen merkittäviä esteitä. Näiden aukkojen kuromiseksi tutkijoiden on keskityttävä edistyneisiin molekyyliprofilointitekniikoihin ja skaalautuviin bioprosessistrategioihin, jotka kestävät pitkäaikaisen tuotannon vaatimukset.
Moni-Omics ja pitkäaikaiset vakaustutkimukset
Suurena haasteena on pitkän aikavälin vakaustietojen puute suunnitelluille solulinjoille. Ajan myötä nämä solut voivat kerätä spontaanisti mutaatioita, mikä voi mahdollisesti muuttaa niiden fenotyyppiä.Ivana Pajčin Novi Sadin yliopistosta korostaa tätä huolta: kuolemattomat solut "eivät aina edusta primaarikulttuuria mahdollisten spontaanien mutaatioiden vuoksi pitkäaikaisen viljelyn aikana" [13]. Ribosomi-insinöörilinjoille panokset ovat vielä korkeammat - ribosomikomponenttien mutaatiot voivat heikentää käännöstehokkuutta ilman välitöntä havaitsemista.
Moni-omiset lähestymistavat tarjoavat keinon puuttua näihin ongelmiin. Integroimalla transkriptomiikkaa, proteomiikkaa ja metabolomiikkaa, tutkijat voivat seurata kriittisiä myogeenisiä markkereita kuten Pax7, MyoD, ja Myogenin, sekä muutoksia MyHC-isomuodoissa. Genomitason metaboliset mallit voivat sitten kääntää nämä oivallukset toiminnallisiksi muutoksiksi kasvatusalustan koostumuksessa vastaamaan insinööriribosomien ainutlaatuisia vaatimuksia [5][11]. Viljellyn lihan osalta johdonmukaisen proteiinituotannon varmistaminen pitkien jaksojen aikana on olennaista. Ilman tällaista pitkittäismonitorointia on vaikea erottaa kestäviä parannuksia lyhytkestoisista vaikutuksista.
Geneettisen ja metabolisen vakauden lisäksi näiden innovaatioiden skaalaaminen teolliselle tasolle tuo mukanaan omat haasteensa.
Bioprosessin integrointi ja skaalaus
Ribosomimuokattujen solujen skaalaaminen pienistä pulloista teollisiin bioreaktoreihin ei ole mikään pieni saavutus. Vain 1 kg proteiinin tuottaminen 5 000 L sekoitetussa säiliöbioreaktorissa vaatii noin kahdeksan biljoonaa lihassolua [5]. Näissä tiheyksissä ravinnegradientit muodostuvat kriittiseksi ongelmaksi. 200 μm diffuusioraja hapelle ja muille ravinteille tarkoittaa, että solut 3D-kudosrakenteiden ytimessä voivat kohdata nälkiintymistä, erityisesti silloin, kun niiden resurssien tarve on huipussaan korkean proteiinisynteesin vuoksi.
Bioreaktorin sekoituksen aiheuttama leikkausjännitys lisää monimutkaisuutta. Vaikka muokkaamattomat solut saattavat sietää tätä turbulenssia, muokatulla käännöskoneistolla varustetut solut voivat olla alttiimpia. Stressi voi paitsi häiritä solujen reittejä, myös fyysisesti vahingoittaa soluja, jotka ovat jo aineenvaihdunnallisessa rasituksessa [13]. Näiden ongelmien ratkaiseminen edellyttää reaaliaikaisten tietojen integroimista digitaalisiin biovalmistusmalleihin, mukaan lukien laskennalliset virtausdynamiikan simulaatiot, jotta voidaan paremmin ymmärtää ja ennustaa suurten astioiden monimuotoisia mikroympäristöjä [10]. Myös jälkikäsittelyprosessit, kuten sadonkorjuu, tarvitsevat huomiota - trypsiiniä sisältävät entsymaattiset menetelmät voivat muuttaa muokattujen solujen pintaproteomia [14], mahdollisesti kumoten ribosomien muokkauksen edut.
| Skaalauskerroin | Keskeinen pullonkaula | Merkitys ribosomien suunnittelulle |
|---|---|---|
| Ravinteiden diffuusio | 200 μm tunkeutumisraja [5] | Voi nälkiinnyttää soluja, joilla on korkeat proteiinisynteesivaatimukset 3D-kudoksissa |
| Geneettinen stabiilisuus | Spontaanit mutaatiot [13] | Voi ajan myötä heikentää suunniteltua käännöstehokkuutta |
| Leikkausjännitys | Sekoitetun säiliön turbulenssi [13] | Voi häiritä suunniteltuja soluprosesseja |
| Sadonkorjuumenetelmä | Proteolyyttinen vaurio trypsiinistä [14] | Voi muuttaa proteomiikkaa ja peittää parannukset proteiinien laadussa |
Näiden laajennushaasteiden ratkaiseminen on olennaista ribosomien suunnittelun siirtämiseksi laboratoriosta kaupalliseen tuotantoon.Jokainen strategia on testattava perusteellisesti, jotta varmistetaan luotettavat proteiinisaannot, vakaus ja turvallisuus teollisissa olosuhteissa.
Päätelmä: Ribosomi-insinöörityön merkitys viljellyssä lihassa
1 kg proteiinin tuottaminen 5 000 L bioreaktorissa vaatii hämmästyttävät 8 biljoonaa lihassolua [5]. Tämä korostaa viljellyn lihan tuotannon skaalaamisen valtavaa haastetta. Ribosomi-insinöörityö tarjoaa ratkaisun parantamalla yksittäisten solujen proteiinituottoa sen sijaan, että yksinkertaisesti lisättäisiin solujen määrää.
Ajoitus on kriittinen ribosomi-insinöörityötä sovellettaessa. Käännöksen tehostaminen väärässä vaiheessa voi häiritä myogeneesiä, mikä voi vaikuttaa keskeisten supistuvien proteiinien, kuten MyHC:n, tuotantoon [5]. Oikean tasapainon saavuttaminen käännöksen ja myogeneesin välillä on yhtä tärkeää kuin itse insinöörityö.
"Jotta saavutettaisiin korkealaatuinen CBM ja sen tuotanto korkealla saannolla, molekyylinäkökulma tarvitsee perusteellisen tarkastelun hyvien laboratoriokäytäntöjen saavuttamiseksi kaupallista tuotantoa varten." - Asim Azhar et al., Frontiers in Food Science and Technology [5]
Useat tekniikat ovat jo osoittaneet lupausta rekombinanttiproteiinin tuotannon lisäämisessä, kuten translaation aloitustekijöiden (eIF3i ja eIF3c) yliekspressio, kodonien optimointi ja mRNA-muutosten kohdentaminen [15] . Kuitenkin näitä menetelmiä on sovellettava huolellisesti, jotta vältetään ongelmat kuten metabolinen kuormitus, proteostaasin stressi ja pitkäaikainen geneettinen epävakaus. Vaikka molekyylin optimointi on olennaista, se ei voi täysin ratkaista haasteita kuten ravinteiden diffuusion rajoitukset, leikkausjännityksen herkkyys ja proteomin häiriöt sadonkorjuun aikana. Nämä esteet vaativat samanaikaisia edistysaskeleita bioprosessisuunnittelussa.
Viljellyn lihan mahdolliset ympäristöhyödyt ovat valtavat. Se voisi vähentää kasvihuonekaasupäästöjä 78%–96%, vähentää maankäyttöä 99%, ja vähentää vedenkäyttöä 82%–96% verrattuna perinteiseen karjatalouteen [12]. Näiden hyötyjen saavuttaminen laajassa mittakaavassa riippuu nykyisen soluviljelytuottavuuden ja taloudellisen toteutettavuuden välisen kuilun kaventamisesta. Ribosomi-insinööritaito on tehokas työkalu tämän kuilun kaventamiseksi, mutta sen on oltava osa laajempaa, integroitua lähestymistapaa, joka sisältää molekyylibiologian, bioprosessi-innovaatiot ja kattavan multi-omiksien seurannan. Vain yhdistämällä nämä ponnistelut voidaan viljellyn lihan koko lupaus toteuttaa.
Kuinka Cellbase Tukee Viljellyn Lihan Tutkimusta
Siirtyminen molekyylien optimoinnista viljellyn lihan laajamittaiseen tuotantoon vaatii tarkkoja työkaluja ja materiaaleja jokaisessa vaiheessa.
Solulinjojen optimointia tekeville tiimeille
Kun tuotantoa laajennetaan,
UKK:t34972>
Mikä ribosomitekniikan lähestymistapa on lupaavin viljellyille lihasolulinjoille?
Tutkimus ribosomitekniikassa viljellyn lihan osalta pyrkii parantamaan proteiinien biosynteesiä ja vaikuttamaan solujen kohtalopäätöksiin. Yksi lupaava lähestymistapa on ribosomipoolin muokkaus, joka muokkaa ribosomaalisia RNA-operooneja parantaakseen käännöstehokkuutta. Työkalut kuten iSAT ja RISE tarjoavat alustoja in vitro ribosomien evoluutiolle, mahdollistaen ribosomien kehittämisen parannetulla toiminnallisuudella. Lisäksi alustat kuten
Kuinka korkeampia käännösmääriä voidaan lisätä ilman, että aiheutetaan väärin laskostuneita proteiineja tai solustressiä?
Parantaakseen käännösmääriä ilman, että aiheutetaan proteiinien väärin laskostumista tai solustressiä, tutkijat keskittyvät hienosäätämään käännösprosessia sen sijaan, että kiihdyttäisivät sitä kauttaaltaan. Joitakin keskeisiä lähestymistapoja ovat:
- Hitaasti kääntyvien kodonien käyttö: Nämä auttavat sovittamaan käännöksen tahdin proteiinien luonnolliseen laskostumisprosessiin, varmistaen oikean rakenteen muodostumisen.
- Vapaan laskostumisenergian vähentäminen 5' koodausalueella: Tämä säätö voi parantaa proteiinituotannon tehokkuutta samalla kun se ylläpitää solun terveyttä.
Muita tekniikoita ovat matalan induktion järjestelmät, lämpötilan laskeminen, ja kehittyneet synteettiset työkalut kuten SINEUP RNA:t. Nämä strategiat mahdollistavat suuremmat proteiinisaannot ilman, että solua kuormitetaan liikaa.
Niille, jotka työskentelevät erikoismateriaalien parissa, resurssit kuten
Mitkä muutokset ovat tarpeen bioreaktoreissa tukemaan ribosomi-insinöityä lihaskudosta yli 200 µm?
Kasvattaakseen lihaskudosta paksummaksi kuin 200 µm, bioreaktoreiden on voitettava haasteet, jotka liittyvät ravinteiden, hapen ja pH:n diffuusioon - tekijät, jotka ovat ratkaisevia solujen selviytymiselle kolmiulotteisissa rakenteissa. Sekoitettavat säiliöbioreaktorit vaativat tarkkoja säätöjä yhtenäisten olosuhteiden ylläpitämiseksi samalla kun vähennetään leikkausjännitystä, joka voisi vahingoittaa soluja. Monissa tapauksissa perfuusioon perustuvat järjestelmät ovat keskeisessä roolissa vakaan ympäristön luomisessa, erityisesti tiheästi pakatuissa kudoksissa. Niille, jotka työskentelevät erikoistuneiden bioreaktoreiden ja materiaalien parissa,









