

히스톤 변형은 DNA를 변경하지 않고 유전자 활동에 영향을 미치는 단백질의 화학적 변화입니다. 이러한 변형은 배양육 생산에 사용되는 세포주 개발에 필수적이며, 세포가 성장하고 정체성을 유지하며 근육 조직으로 분화하는 데 도움을 줍니다. 이 기사는 H3K4me3 (유전자 활성화), H3K27ac (증강기 활동), H3K27me3 (유전자 억제)와 같은 특정 히스톤 마크가 세포 행동을 어떻게 조절하는지 탐구합니다.
다루는 주요 사항:
- H3K4me3는 활성 유전자와 빠른 분화를 지원합니다.
- H3K27ac는 성장 단계 동안 유전자 발현을 위한 증강기를 제어합니다.
- H3K27me3는 원치 않는 유전자 프로그램이 비활성 상태로 유지되도록 보장합니다.
- 이러한 마크에 의해 형성된 크로마틴 상태는 종과 세포 유형에 따라 다르며, 생산 품질에 영향을 미칩니다.
이 기사는 또한 돼지 세포에서 위치적 유전자 발현이 육질에 미치는 영향과 표적 에피제네틱 편집이 세포주 성능을 어떻게 향상시킬 수 있는지를 포함한 최근 연구를 강조합니다. 미래 방향에는 에피제네틱 도구의 정교화와 염색질 상태 연구를 통해 생산 효율성과 규모를 최적화하는 것이 포함됩니다..
히스톤 수정 설명 | 아세틸화, 메틸화 & 유전자 조절
sbb-itb-ffee270
히스톤 수정의 유형과 그 기능
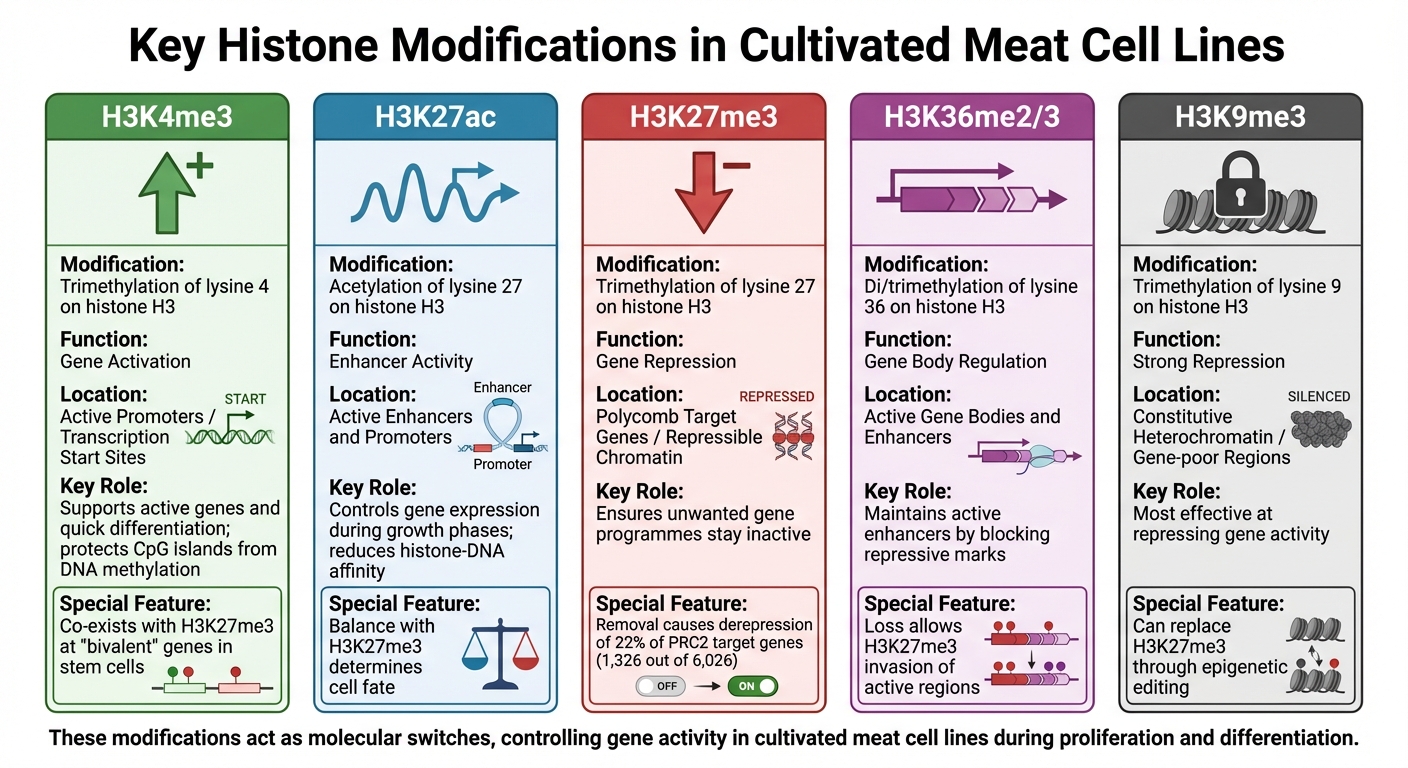
배양육 세포주에서의 주요 히스톤 수정: 기능과 유전체적 맥락
히스톤 수정은 배양육 세포주에서 유전자 활동을 조절하는 데 중요한 역할을 하며, 유전자가 켜지거나 꺼지는지를 제어하는 분자 스위치처럼 작용합니다.이 화학적 태그 - 주로 메틸화와 아세틸화 - 는 히스톤의 특정 잔기에 부착되어 독특한 유전체 패턴을 만듭니다. 각 수정은 특정 기능을 가지고 있으며, 이러한 역할을 이해함으로써 연구자들은 생산 중 세포 행동을 더 잘 예측하고 영향을 미칠 수 있습니다. 이 지식은 배양육 생물공정.
의 프로세스를 최적화하는 데 필수적입니다.배양육 세포주에서 유전자 조절에 영향을 미치는 주요 히스톤 수정의 개요는 다음과 같습니다.
H3K4me3와 유전자 활성화
H3K4me3 (히스톤 H3의 라이신 4의 트리메틸화)는 활성 유전자 프로모터와 관련이 있으며, 특히 세포 성장과 대사에 관여하는 유전자의 시작 지점에서 전사를 촉진합니다. 이 수정은 또한 새로운 DNA 메틸화로부터 CpG 섬 프로모터를 보호하여 필수 유전자가 전사에 접근 가능하도록 보장합니다 [4].
주로 배양육에 사용되는 일차 또는 불멸화된 세포주에서, H3K4me3는 종종 H3K27me3과 같은 억제 마크와 함께 "이중" 유전자에 존재합니다. 이러한 유전자는 활성화를 위해 준비된 상태로 남아 있어 필요할 때 근육 조직으로의 빠른 분화를 가능하게 합니다 [4] .
흥미롭게도, H3K4me3는 다른 변형과 상호작용합니다. 예를 들어, H3K36me3의 침착은 H3K4 메틸트랜스퍼라제를 억제하여 프로모터에서 H3K4me3 수준을 감소시키고 유전자 발현 패턴을 변경할 수 있습니다 [4].
H3K27ac와 인핸서 활동
H3K27ac (히스톤 H3의 라이신 27의 아세틸화)는 활성 인핸서와 프로모터의 마커입니다. 히스톤과 DNA 사이의 친화력을 감소시킴으로써, H3K27ac는 전사를 촉진하는 환경을 조성합니다 [5] . 배양육 세포주에서, H3K27ac 수준의 변화는 세포가 증식에서 분화로 이동할 때 어떤 유전자가 발현되는지를 결정합니다.
H3K27ac와 H3K27me3와 같은 억제적 수식 간의 균형은 세포 운명을 결정하는 데 중요합니다. 예를 들어, 증강기능을 지원하는 H3K36me2의 손실은 H3K27me3가 이전에 활성화된 영역을 침범하게 하여 H3K27ac 수준을 감소시키고 표적 유전자를 침묵시킬 수 있습니다 [5].
H3K27me3와 유전자 억제
H3K27me3 (히스톤 H3의 라이신 27의 트리메틸화)는 유전자를 효과적으로 침묵시키는 닫힌 염색질 구조를 촉진하는 억제적 표식입니다. 이 수식은 Polycomb Repressive Complex 2 (PRC2)에 의해 촉매되며, 수천 개의 발달 유전자의 억제를 유지하는 데 중요합니다 [4] .
배양육 세포주에서 H3K27me3는 특정 성장 단계 동안 원치 않는 유전자 프로그램이 비활성 상태로 유지되도록 하여 세포의 의도된 정체성을 보존합니다.
"H3K27me3는 H2AK119ub1과 함께 수천 개의 Polycomb 표적 유전자의 전사 억제를 유지하는 데 필수적입니다." - Nature Communications [4]
연구에 따르면, 마우스 배아 줄기 세포에서 H3K27me3를 제거하면 PRC2 표적 유전자의 약 22% (6,026개 중 1,326개)가 억제 해제됩니다 [4]. 배양육의 경우, 이 변형을 제어하면 지방이나 결합 조직 형성과 같은 대체 세포 운명을 억제하고 근육 발달에 집중할 수 있습니다.
| 히스톤 수정 | 조절 기능 | 유전체 맥락 |
|---|---|---|
| H3K4me3 | 유전자 활성화 | 활성 프로모터 / 전사 시작 지점 |
| H3K27ac | 증강기 활동 | 활성 증강기 및 프로모터 |
| H3K27me3 | 유전자 억제 | 폴리콤 타겟 유전자 / 억제 가능한 크로마틴 |
| H3K36me2/3 | 유전자 본체 조절 | 활성 유전자 본체 및 증강기 |
| H3K9me3 | 강한 억제 | 구성적 이질염색질 / 유전자 빈약 지역 |
배양육 세포주에서의 크로마틴 상태
히스톤 수정은 단독으로 작용하지 않으며, 크로마틴 상태, 를 형성하기 위해 결합하여 유전자 접근성을 제어하는 독특한 유전체 환경을 만듭니다.이러한 상태는 확장 및 분화 과정에서 배양육 세포주의 행동을 형성하는 데 중요한 역할을 하며, 생물공정 최적화의 핵심 요소입니다.
히스톤 마크를 통한 크로마틴 상태 식별
연구자들은 H3K4me3, H3K27ac, H3K27me3와 같은 히스톤 마크의 조합을 연구하여 크로마틴 상태를 매핑합니다. 예를 들어, 돼지 태아 섬유아세포(PFF)와 영양외배엽(PTr2) 세포에서 10개의 구별된 크로마틴 상태가 식별되었으며, 여기에는 활성 전사 시작 지점, 이중 기능 프로모터, 잠재적 인핸서가 포함됩니다 [6]. 이러한 상태는 유전자 활동을 예측하는 데 도움을 줍니다.
인핸서 상태는 주로 H3K27ac에 의해 비유전자 및 인트론 영역에서 표시되며, 종종 크로마틴 리모델링 단백질 BRG1과 함께 풍부하게 존재합니다 [6].
특히 주목할 만한 특징은 4 kb 이상의 영역을 차지하는 넓은 H3K4me3 도메인, 의 존재입니다. 이러한 도메인은 모든 예측된 전사 시작 지점의 1.7%에서 1.8%에 불과하지만 돼지 세포주에서 발달 및 조직 특이적 유전자를 표시하는 데 중요합니다 [6] . 흥미롭게도, 돼지 태아 섬유아세포에서는 이러한 넓은 도메인으로 표시된 유전자의 52% 가 조직 특이적인 반면, PTr2 세포에서는 25%에 불과합니다 [6].
"이러한 발견은 초기 돼지 발달에서 존재하는 후성유전학적 풍경에 대한 이해를 높이고, 염색질 상태의 변동성이 세포 정체성과 어떻게 연결되는지에 대한 통찰력을 제공합니다." - BMC Epigenetics & Chromatin [6]
이들 크로마틴 상태 프로필은 단일 종 내에서도 다를 뿐만 아니라 배양육 생산에 사용되는 다양한 동물 세포주에서도 차이가 있습니다.
동물 세포주 간의 크로마틴 차이
크로마틴 상태 패턴은 배양육 생산에 사용되는 종과 세포 유형에 따라 크게 달라집니다. 예를 들어, 닭 세포주에서는 H3K4me3가 유전자 프로모터에서 30%에서 55%의 유전체 존재를 차지합니다 [7]. 그러나 닭의 원시 생식 세포(PGC)에서는 H3K4me3 수준이 다능성 세포에 비해 상당히 감소합니다. 이 감소는 생식 계통 사양화 동안 이중 상태가 억제 상태로 전환되는 것을 지원합니다 [7].
돼지 영양막 (PTr2) 세포는 프로모터 영역에서 H3K27ac 수준이 태아 섬유아세포(41.58%)에 비해 높습니다 (57.36%), 반면 H3K27me3 농축은 PTr2 세포(7.77%)에서 PFF 세포(22%)보다 낮습니다 [6]. 이러한 변이는 각 발달 단계의 독특한 후성유전적 요구를 반영하며, 이러한 세포가 배양 조건에 반응하는 방식에 영향을 미칩니다.
소 衛星 세포, 의 분화는 "예비 세포" 운명 (Pax7+/Ki-67-)으로의 분화는 NOTCH 및 MAPK/ERK 신호에 의해 조절되는 휴면 염색질 상태에 의해 주도됩니다. 그러나 이 과정은 단백질 수율을 감소시킵니다 [3]. 이러한 변동성은 염색질 상태가 생산 효율성에 직접적으로 영향을 미친다는 것을 강조합니다. 이러한 차이에 대한 깊은 이해를 얻는 것은 배양육 생산에서 세포주 성능을 미세 조정하는 데 중요합니다.
히스톤 수정을 사용하여 세포주 개선하기
크로마틴 상태에 대한 기존 지식을 바탕으로, 표적 히스톤 수정을 통해 배양육 세포주의 성능을 직접적으로 개선할 수 있는 방법을 탐구해 봅시다.
증식 촉진 및 부유 성장에 대한 적응력 향상
히스톤 마크를 조정하면 세포 증식을 크게 증가시키고 세포가 부착성에서 부유 성장으로 전환하는 데 도움을 줄 수 있습니다. 이 전환은 배양육을 위한 바이오리액터 시스템. 에 매우 중요합니다. 예를 들어, H3K36 메틸화 감소는 섬유아세포가 TGFβ에 덜 반응하게 하여 보다 유연한 세포 상태를 만들어 줍니다 [1].
2022년 12월, Believer Meats 의 연구자들은 닭 섬유아세포(HUN-CF-2 및 HUN-CF-4)와 관련하여 획기적인 성과를 달성했습니다.그들은 무혈청 부유 배양, 에서 자발적인 불멸화를 시연했으며, 100 million cells per ml (10⁸ cells/ml)에 도달하고 36% w/v. 의 수율을 달성했습니다. Yaakov Nahmias가 이끄는 팀은 유전자 변형에 의존하지 않고 지방 형성을 촉진하기 위해 PPARγ 경로를 활성화하는 식품 안전 소분자인 레시틴을 사용했습니다. 그들이 배양한 닭고기 프로토타입은 4.5 out of 5.0 [2] .
"유전자 변형 없는 불멸화와 고수율 제조는 배양육의 시장 실현에 필수적입니다." - Yaakov Nahmias, Believer Meats의 최고 과학 책임자 [2]
이러한 발견은 세포주 개발을 더욱 정밀하게 개선할 수 있는 정밀한 후성유전학 도구의 잠재력을 강조합니다.
에피제네틱 편집의 정밀성
이러한 세포 변화에 보완하기 위해, 정밀한 에피제네틱 편집 방법은 히스톤 마크의 표적 조작을 가능하게 합니다. 2025년 쥐 배아 줄기 세포에 대한 연구는 SUV39H2 또는 SETD2의 촉매 도메인과 융합된 키메라 리크루터(S12N)가 수천 개의 유전자에서 H3K27me3을 H3K9me3 또는 H3K36me3으로 대체할 수 있음을 보여주었습니다. 이 중 H3K9me3이 유전자 활동 억제에 더 효과적임이 입증되었습니다 [8].
그러나 이러한 수정의 성공은 기존의 크로마틴 환경에 크게 의존합니다. 예를 들어, 유전자 프로모터에 남아 있는 H3K4me3은 DNA 메틸화 기계를 차단하여 원하는 유전자 침묵을 달성하기 어렵게 만들 수 있습니다 [8]. 이는 세포 성능을 최적화하려면 단일 수정에 집중하기보다는 여러 히스톤 마크를 동시에 조정해야 한다는 것을 시사합니다.
결론 및 향후 방향
핵심 요약
히스톤 변형은 배양육 세포주에서 유전자 활동을 제어하는 분자 스위치 , 로 중요한 역할을 합니다. 특히, H3K36me2와 H3K36me3는 H3K27me2/3와 같은 억제 마크가 유전자 본체에 침투하는 것을 막아 활성화된 인핸서를 유지하는 데 도움을 줍니다 [9][10]. H3K36 메틸화가 손실되면, 크로마틴 구조가 붕괴되어 H3K9me3와 같은 억제 마크가 활성 영역에 침투할 수 있게 됩니다 [9].
"H3K36 메틸화는 크로마틴 상태와 유전체 구조의 중추적인 조절자입니다." - Nature Communications [9]
히스톤 마크 간의 상호작용은 세포주 성능 향상에 필수적입니다.연구에 따르면 여러 히스톤 변형을 함께 타겟팅하는 것이 단일 변형에 집중하는 것보다 더 나은 결과를 얻는 경우가 많습니다 [4].
이러한 발견을 염두에 두고, 미래 연구는 정밀한 후성유전학 도구를 활용하여 배양육 세포주 성능의 지속적인 개선을 보장해야 합니다.
미래 연구 기회
세포주 성능을 향상시키기 위해서는 단일 핵 RNA 시퀀싱, 과 같은 혁신적인 접근 방식이 필요합니다. 이는 특히 분화에 저항하는 정지 "예비 세포"를 식별하고 이해하는 데 중요합니다. 이러한 세포는 PAX7 및 NOTCH2와 같은 마커를 발현하며, 근육 융합에 전념하지 않고 배양육 생산에 상당한 도전을 제시합니다 [3].
또 다른 유망한 방법은 정밀하고 비유전적인 제어를 위한 키메라 후성유전 복합체의 개발을 포함합니다. 예를 들어, 2025년에 연구자들은 SUZ12의 N-말단을 SUV39H2 또는 SETD2의 촉매 도메인과 결합하여 여러 유전자에서 H3K27me3을 H3K9me3 또는 H3K36me3으로 효과적으로 대체할 수 있음을 입증했습니다 [4] . 또한, 증강기에서 H3K36me2를 모니터링하는 것은 세포주 안정성을 보장하기 위한 품질 관리 마커로 작용할 수 있습니다 [9].
미래의 노력은 세대에 걸쳐 H3K36 메틸화를 유지하는 데 중점을 두어야 합니다. 이는 후성유전적 드리프트를 방지하여 연구자와
자주 묻는 질문
히스톤 마크는 배양육 세포주에서 근육 분화에 어떻게 영향을 미치나요?
히스톤 마크는 특히 배양육 세포주에서 근육 분화의 핵심 요소입니다. 예를 들어, 분화 과정에서 H3K27me3의 감소는 근육 발달에 필요한 유전자의 활성화를 가능하게 하는 근원성 전사 프로그램을 촉발합니다. H3K27me3와 같은 히스톤 수정을 미세 조정하면 세포주가 증식에서 특정 특성을 가진 근육 조직을 형성하는 것으로 전환하는 데 도움이 됩니다. 이러한 후성유전적 조정은 배양육 생산을 발전시키는 데 필수적입니다.
어떤 히스톤 수정이 바이오리액터에서 안정적이고 높은 수율의 세포 성장을 가장 잘 예측하나요?
H3K36 메틸화는 바이오리액터에서 안정적이고 높은 수율의 세포 성장을 위한 신뢰할 수 있는 마커로 두드러집니다.이 수정은 세포 정체성을 보존하고 계통 프로그램을 관리하는 데 중요한 역할을 합니다. 이는 특히 배양육 생산에서 일관된 세포 증식을 보장하는 데 필수적입니다.
후성유전학적 편집이 DNA 서열을 변경하지 않고 세포주를 개선할 수 있습니까?
후성유전학적 편집은 DNA 서열을 변경하지 않고 세포주를 개선할 수 있는 방법을 제공합니다. 히스톤 표지와 염색질 구조를 조정하여 유전자 발현을 제어합니다. 히스톤 수정에 대한 연구는 이러한 변화가 세포 정체성과 기능에 어떻게 영향을 미칠 수 있는지를 강조합니다. 이 접근법은 배양육 세포주를 정제하는 데 특히 유망합니다.










