

For forskere inden for produktion af dyrket kød er det essentielt at minimere apoptose for at forbedre cellelevedygtighed og produktivitet i bioreaktorer. Stressfaktorer som næringsstofmangel, osmotiske ubalancer og ophobning af affald udløser ofte celledød, hvilket reducerer udbyttet. Anti-apoptotiske gener kan afhjælpe disse udfordringer ved at forlænge cellernes levetid under kultur. Her er en hurtig oversigt over de vigtigste gener og deres roller:
- BCL-2: Forhindrer dannelse af mitokondrieporer, hvilket blokerer apoptose ved dens start. Effektiv for udifferentierede celler, men kræver omhyggelig balance med pro-apoptotiske proteiner.
- BCL-xL: Beskytter celler under differentiering og understøtter energimetabolisme. Ideel til høj-stress faser i bioreaktorer.
- MCL-1: Tilbyder hurtig respons på næringsændringer og forbliver stabil under differentiering. Fungerer godt i kombination med andre gener.
- BIRC5 (Survivin) : Hæmmer caspaser for at blokere apoptose nedstrøms. Understøtter proliferation i hurtigt delende celler.
- XIAP: En potent caspasehæmmer effektiv under ekstreme stressforhold, såsom høj-densitetskulturer. Overvågning af disse forhold kræver valg af sensorer til dyrket kød bioreaktorer for at spore næringsstofniveauer og affaldsakkumulering i realtid.
Hurtig Sammenligning
| Gen | Nøgle Rolle | Stabilitet Under Differentiering | Bedste Anvendelsestilfælde |
|---|---|---|---|
| BCL-2 | Blokerer tidlig apoptose (BAX/BAK) | Stabil | Bevare udifferentierede celler |
| BCL-xL | Forhindrer caspase aktivering, understøtter metabolisme | Trin-specifik | Differentiere celler under stress |
| MCL-1 | Hurtig respons på næringsændringer | Stabil | Multi-trins overlevelse |
| BIRC5 | Hæmmer caspaser nedstrøms | Aftager med differentiering | Hurtigt delende celler |
| XIAP | Bred caspase hæmning | Stabil | Høj-stress bioreaktorforhold |
1.BCL-2
BCL-2 er et velundersøgt anti-apoptotisk gen, der spiller en nøglerolle i den intrinsiske (mitokondrielle) apoptosevej. Denne vej er en vigtig mekanisme for celledød, ofte udløst i dyrkede kødceller under bioreaktorstress som næringsstofmangel eller lavt iltniveau.
BCL-2 fungerer ved at binde til og neutralisere pro-apoptotiske proteiner som BAX og BAK. Denne handling forhindrer dannelsen af mitokondrielle porer, stopper frigivelsen af cytochrom c og standser den nedstrøms kaskade af apoptose. Denne mekanisme er afgørende for at forlænge cellernes levedygtige levetid i produktionen af dyrket kød. Som Rønning SB et al. forklarer:
"Forholdet mellem Bcl-2 og Bax bestemmer cellernes modtagelighed for at gennemgå apoptose."[5]
Udover sin mitokondrielle rolle, findes BCL-2 også i det endoplasmatiske retikulum (ER). Her reducerer det calcium niveauer og hæmmer IP3 receptor-medieret calciumfrigivelse, hvilket mindsker calcium-induceret apoptose – et hyppigt problem i høj-densitets bioreaktor kulturer[4]. Håndtering af disse skalering udfordringer er et primært fokus for industrien. Denne dobbelte lokalisering gør det muligt for BCL-2 at beskytte celler mod flere apoptose udløsere.
Den molekylære struktur af BCL-2, bestående af en otte-alfa-helix bundt og fire veldefinerede BH domæner, gør det til en e
Der er dog en kritisk forbehold: balancen mellem BCL-2 og pro-apoptotiske proteiner som BAX skal håndteres omhyggeligt. Selv høje niveauer af BCL-2-ekspression kan ikke forhindre apoptose, hvis pro-apoptotiske signaler bliver for stærke[2]. Overvågning af denne balance er afgørende for at opnå optimal cellelevedygtighed.
sbb-itb-ffee270
2. BCL-xL
BCL-xL, kodet af BCL2L1 genet, spiller en central rolle i BCL-2 familien ved at lokaliseres til den ydre mitokondriemembran og forhindre apoptose. Det opnår dette ved at modvirke pro-apoptotiske proteiner som BAX og BAK.Derudover hæmmer det kløvet caspase-3 (CASP3), som er essentiel for at standse celledød. Denne mekanisme er særligt værdifuld i høj-densitets bioreaktor kulturer , hvor metabolisk stress kan true cellelevedygtighed.
Interessant nok er BCL-xL's aktivitet i overensstemmelse med specifikke stadier af differentiering. Under visse faser øges dets udtryk, mens andre anti-apoptotiske proteiner, såsom BCL-2 og MCL-1, forbliver uændrede. Dette understreger dets betydning i at opretholde celleoverlevelse under differentiering. Som nævnt i Cell Death & Disease:
"BCL-xL/BCL2L1 er et kritisk anti-apoptotisk protein, der fremmer overlevelsen af differentierende... celler." [2]
Udover sin rolle i apoptose understøtter BCL-xL cellulær energimetabolisme. Det forbedrer både glykolyse og oxidativ fosforylering, hvilket sikrer høj metabolisk aktivitet.Hæmning af BCL-xL har vist sig at reducere udtrykket af metaboliske gener og sænke både basal og maksimal mitokondriel respiration. Denne funktion er særligt vigtig for dyrkede kød celler, som er afhængige af vedvarende metabolisk output.
BCL-xL er meget kompatibel med genredigeringsstrategier, der almindeligvis anvendes i forskning i dyrket kød. Teknikker som lentiviral transduktion muliggør stabil integration af BCL2L1 genet, mens doxycyclin-inducerbare CRISPR/Cas9-systemer giver præcis tidsmæssig kontrol over dets udtryk [2] [6]. Dette niveau af præcision styres ofte gennem avanceret bioproceskontrolsoftware. Disse egenskaber gør BCL-xL til en stærk kandidat til at forbedre cellelinjens levedygtighed i produktionen af dyrket kød.
For differentieringsstadier med høje metaboliske krav kan BCL-xL være mere effektiv end BCL-2.Forskere kan bruge hæmmeren WEHI-539 til at teste en cellelinjes afhængighed af BCL-xL, før de fortsætter med permanente genetiske modifikationer [2]. Derudover kan co-ekspression af BCL-xL med MCL-1 yderligere forbedre celleoverlevelse, da disse proteiner er blevet observeret at arbejde synergistisk i nogle resistente celletyper [6].
3. MCL-1
MCL-1 (Myeloid Cell Leukaemia-1) spiller en nøglerolle i reguleringen af den intrinsiske apoptotiske vej. Fundet på den ydre mitokondriemembran, forhindrer det apoptose ved at binde og sekvestrere de pro-apoptotiske proteiner BAX og BAK, hvilket stopper deres oligomerisering og efterfølgende membranpermeabilisering. Denne handling blokerer frigivelsen af cytochrom c, hvilket stopper den apoptotiske kaskade, før den når eksekveringsfasen [8] . Derudover binder MCL-1 BH3-kun proteiner - såsom Bim, PUMA og NOXA - med høj affinitet [8]. Ligesom BCL-2 og BCL-xL er MCL-1 afgørende for at modvirke apoptotiske signaler, især under bioreaktor stress.
En af MCL-1's unikke egenskaber er dens korte halveringstid, hvilket gør dens udtryk meget responsivt over for næringstilgængelighed og metaboliske signaler, især gennem AMPK/mTOR-vejen. Studier indikerer, at en 25% reduktion i kalorieindtag kan mindske MCL-1 translation med cirka 39% ± 10% [7]. Denne følsomhed er især relevant for produktion af dyrket kød, hvor udsving i vækstmediekomposition eller næringsstofudtømning under storskalasuspensionskulturer (som kræver omhyggelig produktionsskala planlægning) kan betydeligt sænke MCL-1 niveauer.Sådanne reduktioner kompromitterer cellelevedygtigheden, hvilket underminerer forbedringerne i IVCC (integral viable cell concentration) opnået gennem anti-apoptotiske strategier. For at afbøde dette er serumfrie medieformuleringer, der understøtter robust mTORC1-aktivitet, essentielle [7] .
En anden bemærkelsesværdig egenskab ved MCL-1 er dens stabilitet under differentiering. I pankreatiske progenitormodeller forblev MCL-1-ekspression stabil gennem en 17-dages differentieringsprotokol, i modsætning til BCL-xL, som viste trin-afhængig variation [2]. Denne stabilitet gør MCL-1 særligt fordelagtig for dyrkede kødapplikationer, hvor celler skal overleve flere modningsstadier uden at kræve præcist timede interventioner.&
Genredigeringsværktøjer kan bruges til at modificere MCL-1, ligesom andre anti-apoptotiske gener, hvilket gør det til et alsidigt mål for cellelinjeingeniørarbejde.
Når det bruges i kombination med andre anti-apoptotiske gener, tilbyder MCL-1 yderligere fordele. For eksempel har kombinationen af MCL-1 med BCL-xL vist synergistiske effekter - samtidig hæmning af begge proteiner reducerede EC50 for overlevelsesmedicin fra omkring 10 μM til mindre end 20 nM [6]. Denne tilgang kan betydeligt forbedre celleoverlevelse under de høj-stress faser af dyrket kødproduktion.
4. BIRC5 (Survivin)
BIRC5, ofte omtalt som Survivin, er et medlem af Inhibitor of Apoptosis (IAP) proteinfamilien [2]. I modsætning til BCL-2 familieproteiner, som virker ved mitokondriemembranen for at forhindre initiering af apoptose, opererer BIRC5 længere nede i processen. Det blokerer de caspaser, der er ansvarlige for at udføre apoptose, og fungerer effektivt som en sidste forsvarslinje mod programmeret celledød [10].
I suspensionskulturer kan stressfaktorer som næringsstofmangel, ophobning af metabolisk affald og mekanisk skærestress udløse apoptose. Ved at hæmme caspase-aktivitet på dette senere stadium hjælper overekspression af BIRC5 med at forlænge cellelevedygtighed og produktivitet. Dette resulterer i en forbedring af det tidsintegrerede levedygtige cellekoncentration - en nøglemetrik til optimering af cellekulturens ydeevne [9] . Eric Baek, en forsker ved KAIST, forklarer:
"Forbedring af det tidsintegrerede levedygtige cellekoncentration ved at overvinde celledød, nemlig apoptose, er en af de mest udbredte strategier til effektiv produktion af terapeutiske proteiner [og celler]." [9]
Denne downstream intervention har vist sig at forbedre bioreaktorudbyttet i dyrkede kødcellinjer, herunder porcine satellitceller og bovine myoblaster.
Den mest effektive strategi involverer kombinatorisk ingeniørarbejde, parring af BIRC5 med mitokondrie-beskyttere som BCL-2 eller BCL-xL. Professor Michael Betenbaugh fra Johns Hopkins University fremhæver denne tilgang:
"Strategier, der blokerer celledød på flere punkter langs kaskaden, kan begrænse forstærkningen af disse apoptosesignaler." [10]
Ved at kombinere BIRC5's caspase-hæmning med opstrøms mitokondrie-beskyttelse kan forskere etablere et flerlags forsvar mod apoptose.
BIRC5 integreres også problemfrit i genredigeringsarbejdsgange.CRISPR/Cas9 er den førende metode til at skabe stabile cellelinjer med overekspression [9], selvom zink-finger nukleaser tilbyder et præcist alternativ. siRNA kan bruges til validering af pathways, før man forpligter sig til genomisk integration [9].
5. XIAP
XIAP (X-linked inhibitor of apoptosis) er anerkendt som den mest potente caspasehæmmer inden for IAP (inhibitor of apoptosis protein) familien. Sammen med gener som BCL-2 og MCL-1 spiller XIAP en kritisk rolle i at målrette apoptose i dens eksekveringsfase. Som fremhævet i Genes & Development :
"XIAP betragtes som den mest potente caspasehæmmer in vitro." [12]
XIAP anvender to forskellige mekanismer til at hæmme apoptose. Først blokerer dens BIR2-domæne og linkerregion effektorcaspaser-3 og -7.Andet, dets BIR3-domæne hæmmer caspase-9, hvilket effektivt stopper den indre mitokondrielle apoptotiske vej. Derudover faciliterer dets C-terminale RING-domæne ubiquitinering og efterfølgende proteasomal nedbrydning af mål-caspaser [11]. Ved at gribe ind i både indre og ydre apoptotiske veje, viser XIAP sig yderst effektiv til at håndtere apoptoseudløsere som næringsstofmangel, metaboliske biprodukter og mekanisk stress - faktorer, der ofte opstår i dyrkede kødproduktionssystemer. Dens funktionalitet forbedres yderligere af dens stærke bevarelse på tværs af arter.
For eksempel deler menneskelig XIAP 87,7% proteinidentitet med Bos taurus (bovin) og 89,5% med Mus musculus (mus) [11] . Denne høje lighed gør det muligt at anvende forskning fra pattedyrsmodeller pålideligt på cellelinjer, der bruges i dyrket kødproduktion.
XIAP kan reguleres ved hjælp af værktøjer som shRNA, antisense oligonukleotider eller CRISPR/Cas9 [11]. Under ekstrem stress kan dets RING-domæne inducere selv-ubiquitinering [12], mens endogene inhibitorer som SMAC/DIABLO og HTRA2 kan fortrænge XIAP fra caspaser [11][13]. Disse fund gør XIAP til et attraktivt mål for genredigeringsmetoder, der sigter mod at optimere cellelinjer til udvikling af dyrket kød.
Sammenligning af anti-apoptotiske gener på et øjeblik
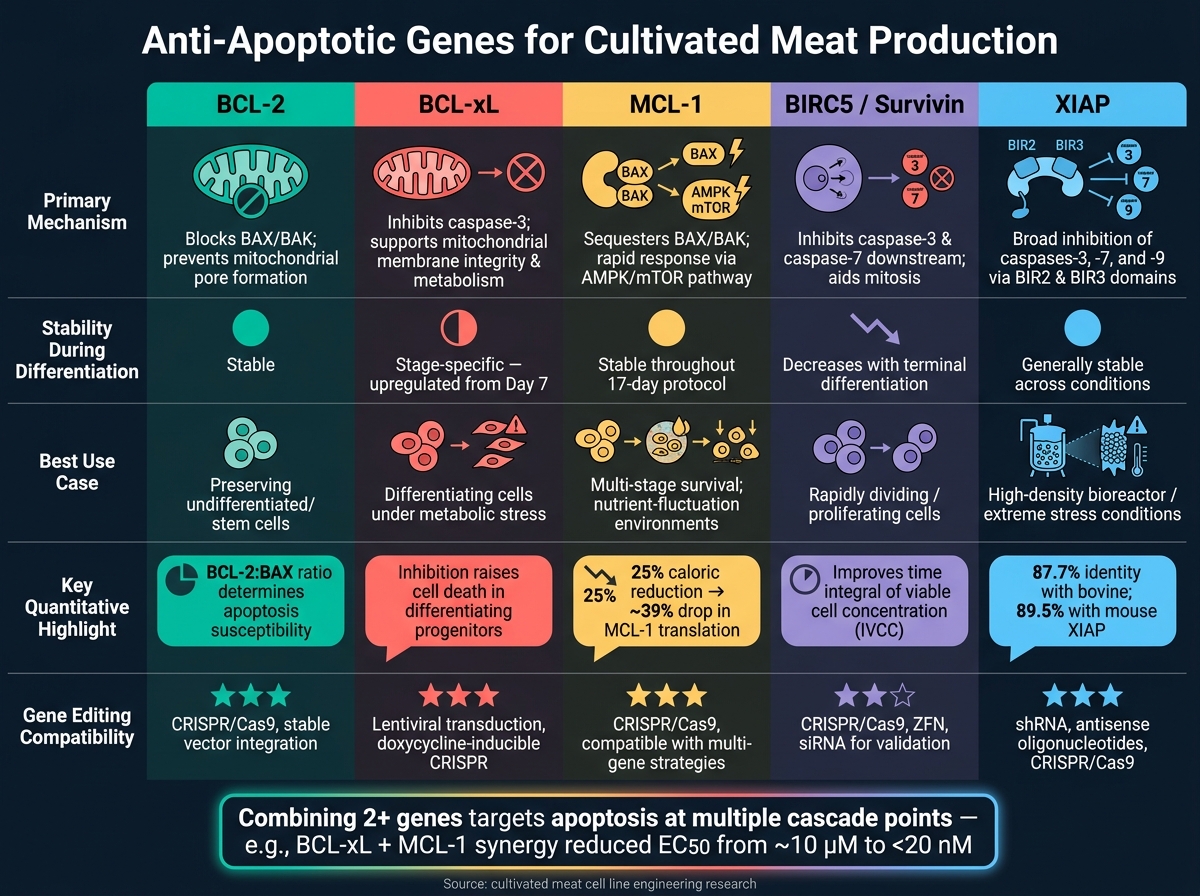
Anti-apoptotiske gener til dyrket kød: Sammenligning side om side
Når man arbejder med produktion af dyrket kød, kan forståelsen af, hvordan forskellige anti-apoptotiske gener fungerer, hjælpe med at finjustere cellelinjeingeniørarbejdet. Hvert gen har sin egen unikke mekanisme, adfærd under differentiering og potentielle anvendelser. Tabellen nedenfor opsummerer disse forskelle, hvilket gør det lettere at beslutte, hvilket gen - eller kombination af gener - der bedst kan opfylde dine behov.
| Gen | Primær Mekanisme | Ekspressionsstabilitet | Rapporteret levedygtighedspåvirkning | Redigeringskompatibilitet |
|---|---|---|---|---|
| BCL-2 | Blokerer pro-apoptotisk BAX/BAK og sikrer overlevelse af udifferentierede celler [2] | Forbliver relativt stabil under differentiering [2] | Essentiel for at bevare den oprindelige stamcellepulje [2] | Høj kompatibilitet med redigeringsværktøjer |
| BCL-xL | Hæmmer kløvet caspase-3; opretholder mitokondriemembranens integritet og metabolisme [2] | Opreguleret fra dag 7 af differentiering [2] | Kritisk for at støtte differentierende progenitorer; dets hæmning øger celledød [2] | Høj kompatibilitet med redigeringsværktøjer |
| MCL-1 | Modulerer pro-apoptotiske signaler som en del af BCL-2 familien [2] | Ekspression forbliver stabil under linjespecifikation [2] | Tilbyder brede overlevelsesfordele men mangler stadiespecifikke effekter som BCL-xL [2] | Høj kompatibilitet med redigeringsværktøjer |
| BIRC5 (Survivin) | Blokerer caspase-3 og caspase-7; hjælper med kromosomal segregation under mitose | Høj i prolifererende celler; falder med terminal differentiering | Understøtter overlevelse og proliferation i hurtigt delende celler | Kompatibel med både shRNA knockdown og CRISPR-redigering |
| XIAP | Hæmmer flere caspaser, hvilket giver bred apoptotisk beskyttelse | Generelt stabil under forskellige betingelser | Særligt effektiv under stress, såsom høj-densitets bioreaktorforhold | Høj kompatibilitet med redigeringsværktøjer |
BCL-xL skiller sig ud for sin dobbelte rolle i at fremme celleoverlevelse og understøtte metabolisk aktivitet, især under den kritiske differentieringsfase, når pro-apoptotiske proteiner som BAK naturligt falder.BCL-2, på den anden side er ideel til at bevare udifferentierede celler, mens XIAP leverer bred beskyttelse, især i stressende miljøer som høj-densitetskulturer.
Intet enkelt gen fungerer bedst i alle scenarier. For eksempel er BIRC5 særligt nyttigt i situationer, der kræver hurtig celledeling. I praksis tilbyder kombinationen af to eller flere gener ofte den mest effektive beskyttelse, idet de adresserer en række apoptotiske udløsere samtidigt.
Disse fund giver et grundlag for at inkorporere disse gener i cellelinje-ingeniørstrategier for dyrket kødproduktion. Dette inkluderer valg af de rigtige dyrkede kødsinput for at sikre skalerbarhed.
Brug af Disse Gener i Dyrket Kød Cellelinje Ingeniørarbejde
For at forbedre cellelevedygtigheden i dyrket kødproduktion er det afgørende at integrere nøglegener strategisk.Det er ikke nok at identificere anti-apoptotiske gener - deres effektive inkorporering i cellelinjer er det, der gør forskellen. To hovedstrategier anvendes almindeligvis: overekspression af anti-apoptotiske gener som BCL-2, BCL-xL, og MCL-1 for at forbedre celleoverlevelse, eller udslåning af pro-apoptotiske gener såsom BAX, BAK, og BOK for at eliminere drivkræfterne bag celledød. Kombination af disse tilgange resulterer ofte i cellelinjer, der er bedre egnet til storskalaproduktion [1].
Moderne genredigeringsværktøjer som CRISPR/Cas9 muliggør samtidige redigeringer, såsom udslåning af Bak1, Bax, og Bok i ét trin. Alternativer som ZFNs eller RNA-interferens kan bruges til midlertidigt at reducere aktiviteten af caspaser ( e.g. caspases-3, -7, -8, og -9). For overexpression strategies, synthetic promoters ensure consistent and high expression levels of genes like BCL-2 during scale-up, which is critical for maintaining cell performance in fed-batch or continuous culture systems . Disse kombinerede metoder styrker cellelinjeudvikling til anvendelser inden for dyrket kød.
Sådanne genetiske modifikationer påvirker direkte forbedret integral viable cell concentration (IVCC), et nøglemål i produktionen af dyrket kød. Celledød er mest udtalt i løbet af de første fem dage af differentiering, hvilket gør tidlige interventioner med gener som BCL-2 eller BCL-xL essentielle. Forskning offentliggjort i Cell Death & Disease fremhæver, at BCL-xL ekspression øges, når celler differentierer, hvilket indikerer, at mere modne progenitorer er stærkt afhængige af dens beskyttende rolle [2] . Ved at overvåge udtryksniveauerne af BCL-2 familie gener gennem vækstfaserne, kan interventioner præcist tidsbestemmes for maksimal effekt.
"Ved at etablere stabile cellelinjer, der overudtrykker antiapoptotiske gener eller nedregulerer proapoptotiske gener, kan det endelige produktudbytte forbedres, da cellerne bliver mere modstandsdygtige over for miljømæssige stressfaktorer." - Gyun Min Lee et al. [1]
For bioreaktorbaseret produktion skal celler også konstrueres til at modstå hyperosmotisk stress og næringsmangel. Før opskalering er det essentielt at validere genetiske redigeringer ved hjælp af værktøjer som Western blot eller FACS. For forskere, der søger specialiserede cellelinjer eller genetiske materialer skræddersyet til høj-densitets bioreaktormiljøer, tilbyder platforme som
Konklusion
Valg af anti-apoptotiske gener til dyrkede kød cellelinjer kræver en skræddersyet tilgang. Gener som BCL-2, BCL-xL, og MCL-1 spiller hver især unikke roller i at beskytte celler, men deres succes afhænger af faktorer som celletype, udviklingsstadie og de specifikke stressfaktorer, der opstår under produktionen. Som fremhævet i forskning:
"balancen mellem de anti-apoptotiske og pro-apoptotiske medlemmer bestemmer i sidste ende, om en celle lever eller dør" [2]
Udover overlevelse bevarer anti-apoptotisk engineering også metaboliske funktioner. For eksempel er proteiner som BCL-xL tæt forbundet med opretholdelse af glykolyse og oxidativ fosforylering. Dog kan dårligt udførte interventioner forstyrre disse kritiske processer [2]. At sikre, at konstruerede cellelinjer bevarer deres tilsigtede identitet og metaboliske aktivitet gennem hele produktionen, er et afgørende, men nogle gange overset, trin. Disse indsigter former fremtiden for cellelinjeingeniørkunst.
Nye multi-gen tilgange dukker op, som kombinerer overekspression af beskyttende gener med CRISPR knockouts af pro-apoptotiske gener som BAX, BAK1, og BOK for at skabe mere robuste cellelinjer til industriel brug [1]. Værktøjer til metabolisk profilering, såsom bioenergetiske assays, bliver essentielle for at bekræfte, at disse genetiske modifikationer forbedrer den samlede celleydelse. For forskere, der søger svinecellelinjer , genetiske materialer eller bioreaktorudstyr,
Ofte stillede spørgsmål
Hvilket anti-apoptotisk gen skal jeg starte med til min cellelinje?
BCL-2 foreslås ofte som et udgangspunkt, når man arbejder med cellelinjer. Dette velundersøgte anti-apoptotiske gen er anerkendt for sin evne til at forbedre celleoverlevelse, hvilket gør det til en populær mulighed inden for forskning i dyrket kød. Dets funktion i at støtte cellelevedygtighed gør det til et praktisk valg til tidlige eksperimenter.
Er det bedre at overudtrykke anti-apoptotiske gener eller slå pro-apoptotiske gener ud?
Inden for produktion af dyrket kød giver det ofte bedre resultater at øge udtrykket af anti-apoptotiske gener, såsom medlemmer af BCL-2 familien som BCL-xL, end at deaktivere pro-apoptotiske gener. Denne strategi understøtter både celleoverlevelse og proliferation - nøglefaktorer for opskalering af produktionen - samtidig med at cellens naturlige reguleringssystemer bevares.
Ved at øge anti-apoptotisk genaktivitet opnår celler større modstandsdygtighed over for apoptose, især under stressende forhold. Dette gør det til en mere pålidelig og sikker tilgang til at opretholde cellelevedygtighed under dyrkningsprocessen.
Hvordan kan jeg bekræfte, at en anti-apoptotisk redigering forbedrer IVCC i min bioreaktor?
For at afgøre, om en anti-apoptotisk genredigering forbedrer in vitro cellelevedygtighed og proliferation (IVCC), skal du bruge en systematisk tilgang:
- Vurder levedygtigheds- og proliferationsrater: Brug metoder som celletælling eller flowcytometri til at måle disse rater både før og efter genredigeringen.
- Bekræft genekspression: Teknikker som qPCR eller Western blotting kan bekræfte den vellykkede ekspression af det målrettede gen.
- Overvåg apoptosemarkører: Kontroller for markører som caspaseaktivitet for at sikre, at redigeringen effektivt reducerer apoptose.
For en komplet evaluering er det kritisk at teste den langsigtede stabilitet og proliferation af de redigerede celler i en bioreaktor. Dette sikrer, at forbedringerne vedvarer gennem flere kulturcyklusser.










