

Hvis du kan udvide celler, men ikke kan skifte dem til den rette skæbne på det rette tidspunkt, vil din proces gå i stå ved differentiering. Det er hovedpunktet her: syntetiske genkredsløb giver dig kontrol inde i cellen over forpligtelse, timing, hukommelse og stamlinjeblanding, hvor medieændringer alene ofte efterlader heterogene, delvist forpligtede populationer.
Hvis jeg skulle bygge en differentieringsarbejdsgang for dyrket kød, ville jeg straks tage fire punkter fra denne artikel:
- Start med det oprindelige netværk, ikke konstruktionen. Brug snRNA-seq, trajektorieanalyse, GRN-inferens og miRNA-profilering for at finde ud af, hvor celler går i stå, driver eller forgrener sig til den forkerte skæbne.
-
Match kredsløbstype til procesproblemet.
En tænd/sluk-kontakt passer til lock-in, et feedforward- eller båndpasdesign passer til tidsstyring, en logikport passer til multi-signal gating, og miSFITs passer til gradueret output. - Design for lav lækage, lav støj og sikkerhed fra dag ét. Ortogonale dele, negativ autoregulering, iFFLs, cm transgener og et inducerbart dræb- eller vækststopmodul er en del af opbygningen, ikke en eftertanke.
- Validér under skala-relevante forhold tidligt. Et kredsløb, der fungerer i 2D, kan ændre sig i 3D, mikrobærer eller omrørt suspension på grund af inducergradienter, iltbegrænsninger og skær.
Artiklen gør også et praktisk punkt, der er vigtigt for procesteam: enkelt-linje kontrol og forholdskontrol er forskellige opgaver. En Tet-On MyoD kassette kan fremme myogen indtræden, men helskårne produkter kræver kontrol over muskel, fedt og ECM proportioner, hvilket normalt betyder feedback, parakrin signalering og mere omfattende klonscreening.
Nogle få tal understøtter denne meddelelse:
- Standard myogen differentiering kan stoppe med fusionsindekser på cirka 50–60%
- Konstruerede GRN'er i iPSCs øgede mål-linje differentiering fra 52% til 81%
- Syntetiske kredsløb i modificerede MSC'er drev hjertedifferentiering til 76%
- Nogle svine Tet-On-PAX7 linjer bevarede høj myogen potentiale ud over 40 passager
- Omkring 20% af humane pluripotente stamceller kan bære kræft-relaterede mutationer, hvilket er grunden til, at inducerbare sikkerhedsmoduler er vigtige
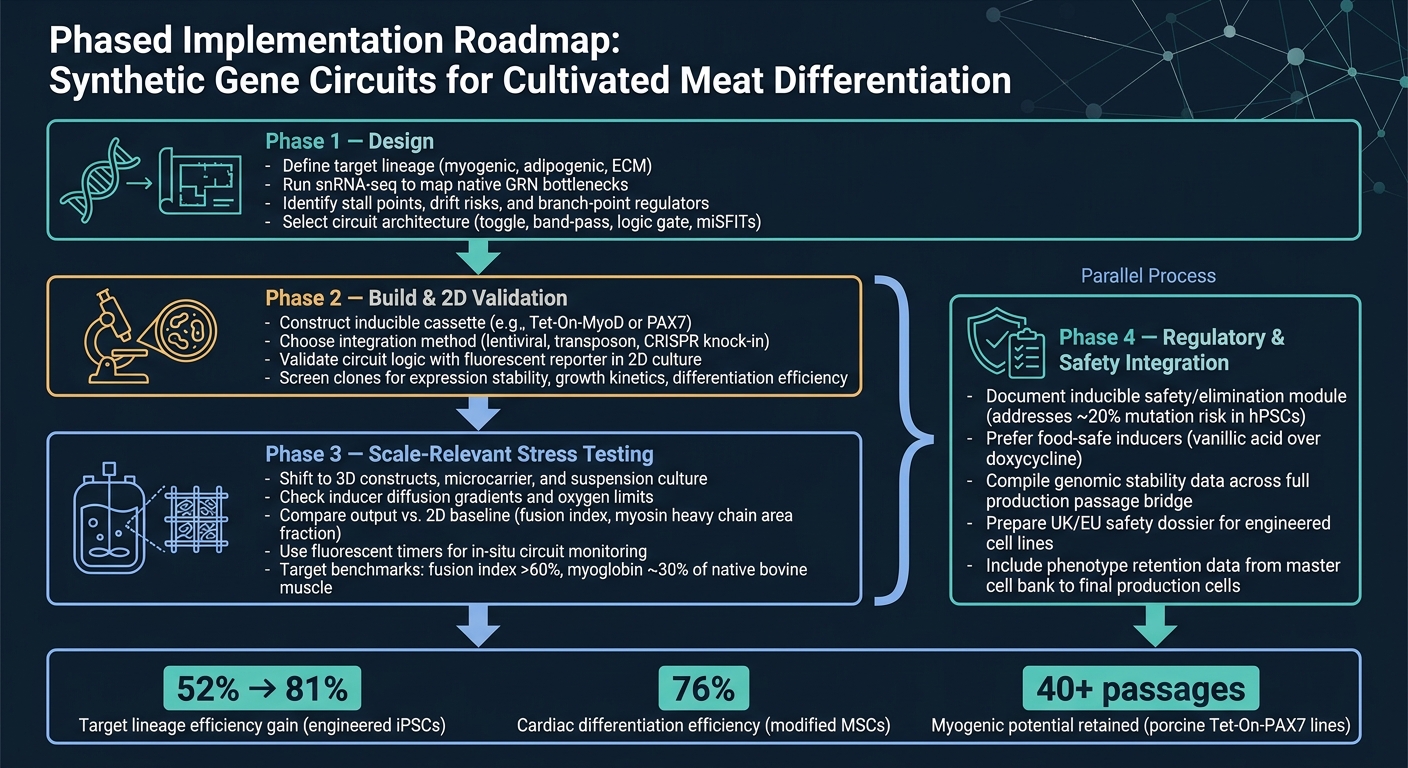
Syntetisk Genkredsløb Arbejdsgang for Dyrket Kød Differentiering
Forskerhistorier: Michael Elowitz, Genetiske Kredsløb i Levende Celler
Hurtig sammenligning
| Emne | Hvad artiklen siger i enkle termer |
|---|---|
| Hovedflaskehals | Differentiation, ikke ekspansion |
| Hvorfor kredsløb hjælper | De tilføjer tærskelkontrol og celle-tilstand hukommelse inde i cellen |
| Bedste første skridt | Kortlæg naturlig skæbnekontrol med snRNA-seq og relaterede værktøjer |
| Hovedkredsløbs muligheder | Toggle, feedforward, band-pass, logiske porte, miSFITs, CRISPRa/i |
| Myogenese eksempel | Tet-On-MyoD for at adskille vækst fra terminal differentiering |
| Adipogenese / ECM kontrol | miSFITs og bow-tie designs for gradueret PPARG/BMP4-type output |
| Hele-skæring udfordring | Forholdskontrol på tværs af muskler, fedt og bindevæv |
| Opskaleringsrisiko | 2D-adfærd holder muligvis ikke i 3D eller bioreaktorer |
| Integrationsvalg | Lentivirus, transposoner, CRISPR knock-in, episomale vektorer |
| Regulatorisk punkt | Konstruerede linjer har brug for en bredere sikkerhedspakke; fødevaresikre induktorer som vanillinsyre foretrækkes frem for DOX, hvor det er muligt |
Så, kort sagt: dette er ikke bare en artikel om kredsløbsdesign. Jeg ville læse det som en guide til at forbinde kredsløbsarkitektur, afstamningsbiologi, klonvalg, bioreaktorpræstation og UK/EU sikkerhedsdokumentation til en differentieringsstrategi.
Læs videre, hvis du vil have den fulde vej fra kortlægning af det oprindelige netværk til valg af konstruktion, opskaleringstjek og regulatorisk tilpasning.
2. Designprincipper for differentieringskontrolkredsløb
2.1 Kortlæg det oprindelige celle-skæbne netværk, før du designer kredsløbet
Før du designer et kredsløb, har du brug for et klart billede af, hvad cellen allerede gør.
Single-nucleus RNA-sekventering (snRNA-seq) er et godt sted at starte. Det kan vise hvilende subpopulationer, inklusive reserveceller markeret af NOTCH2 og HEYL , og pege på vejmål, der kan forbedre differentiering [3].
Derfra hjælper baneanalyse og genregulatorisk netværk (GRN) inferens med at kortlægge rækkefølgen af regulatoraktivering og fremhæve, hvor celler sandsynligvis vil gå i stå. I myogenese løber hovedkaskaden gennem MYOD1 og MYOG . I adipogenese er hovednoderne PPARG og CEBPA , med fibro-adipogene progenitor (FAP) skæbne som den vigtigste forgreningspunkt risiko. Tabellen nedenfor opsummerer de vigtigste regulatorer og flaskehalse.
| Afstamning | Vigtige master regulatorer | Kritiske signalveje | Identificerede flaskehalse |
|---|---|---|---|
| Myogenisk | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Reserve celle dannelse (quiescence) |
| Adipogenisk | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Fibro-adipogene progenitor (FAP) skæbne |
| Pluripotent | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Spontan differentiering / heterogenitet |
Et andet nyttigt lag er miRNA udtryksprofilering. Endogene miRNA'er såsom miR-302a, forbundet med pluripotens, og miR-375, forbundet med differentiering, kan fungere som interne klassifikatorer i sense-and-respond designs. Det lader kredsløbet læse cellens faktiske tilstand i stedet for kun at afhænge af eksterne induktorer [5].
Disse flaskehalse bør drive kredsløbsvalget. Hvis hovedproblemet er drift, kan du have brug for en lock-in. Hvis timing er problemet, kan en puls passe bedre. Hvis skæbnekontrol afhænger af mere end et signal, giver multi-input logik normalt mere mening.
2.2 Vælg den rigtige kredsløbsarkitektur
Det er her, kompromiserne viser sig tydeligt. Den rigtige arkitektur afhænger af tre praktiske punkter: hvor permanent forpligtelsen skal være, hvor stramt timingen skal kontrolleres, og hvor meget genetisk belastning cellen kan bære uden problemer.
Bistabile toggle switches passer til tilfælde, hvor linjeforpligtelse skal forblive låst. Hovedproblemet er spontan tilstandsændring forårsaget af transkriptionel støj.
Båndpasfiltre passer til tilfælde, hvor en transkriptionsfaktor kun er nødvendig inden for et defineret udviklingsvindue. Ulempen er, at inducer-niveauerne skal være under stram kontrol, ellers glider timingen.
Logiske porte tilføjer specificitet ved at kræve mere end én input på samme tid. For eksempel kan differentiering kun tillades, når en eksogen inducer er til stede og cellen viser den rigtige endogene miRNA-profil. Det hjælper med at reducere risikoen for off-target forpligtelse.
Tabellen nedenfor skitserer de vigtigste arkitekturer og de kompromiser, der følger med dem.
| Arkitektur | Reversibilitet | Tidsmæssig præcision | Integrationskompleksitet | Primær anvendelse | Nøgle risiko |
|---|---|---|---|---|---|
| Bistabil kontakt | Lav (når låst) | Moderat | Moderat | Permanent afstamningsforpligtelse | Spontan flipping på grund af støj |
| Båndpasfilter | Høj (koncentrationsafhængig) | Høj | Høj | Forbigående udviklingsstadier | Kræver stram inducerkontrol |
| Logisk port (AND/OR/NOT) | Variabel | Moderat | Moderat–høj | Celletype-specifik aktivering | Lækage i OFF-tilstande |
| Butterfly / multi-input | Høj | Moderat | Moderat | Multi-signal integration | Afhænger af endogen miRNA stabilitet |
| miSFITs | Høj | Moderat | Lav–moderat | Graderet output tuning | Smal dynamisk rækkevidde hvis dårligt tunet |
"Ved at minimere antallet af beregningslag samtidig med at funktionaliteten opretholdes, adresserer denne strategi skalerbarhedsbarrierer i genkredsløbsingeniørkunst." - Nature Communications [9]
Hvert tilføjet reguleringslag øger belastningen og trækker på cellulære ressourcer. I praksis, hvis to design udfører det samme job, er det enklere normalt det bedre valg, når skalaen betyder noget.
Når arkitekturen er fastlagt, er den næste opgave at få den til at holde under lav lækage, støjundertrykkelse og fejlsikker kontrol.
2.3 Byg for pålidelighed, lav lækage og sikkerhed
Et kredsløb skal forblive stabilt over længere kultur. En kort periode med god ydeevne er ikke nok til produktionsbrug.
Ortogonale dele er det første forsvar. Promotorer, transkriptionsfaktorer og reguleringselementer, der ikke krydsreagerer med det native maskineri, hjælper med at begrænse off-target effekter og reducerer chancen for, at endogene signaler vil tænde eller slukke kredsløbet.Modificerede høj-stramhedspromotorer såsom PCREm er blevet brugt til at reducere basal lækage i inducerbare pattedyrsystemer [6] .
Negativ autoregulering er også værd at tilføje, hvor det er muligt. Det er et af de bedre kendte motiver til at reducere transkriptionel støj og producere en mere lineær respons på inducerkoncentration [6] . Inkoherente feedforward-sløjfer (iFFLs) kan tilføje et andet lag af kontrol ved at filtrere stokastiske udsving, så celler reagerer på vedvarende signaler frem for korte støjspidser.
Kodon-modificerede ( cm) versioner af syntetiske transkriptionsfaktorer gør karakterisering lettere. De lader dig adskille kredsløbsdrevet udtryk fra endogent genomisk (g) udtryk under validering [1]. Det kan lyde som en lille detalje, men det sparer tid, når du prøver at finde ud af, om en aflæsning kommer fra kredsløbet eller værtsgenomet.
Sikkerhedsmoduler er påkrævet. Omkring 20% af humane pluripotente stamceller bærer kræftassocierede mutationer [7] . Så hvis et kredsløb skal ind i en stamcelle-afledt linje, bør det inkludere et inducerbart vækststop eller elimineringsmodul. Vanillinsyre er en nyttig inducer at prioritere her, fordi det er et godkendt fødevaretilsætningsstof, hvilket hjælper sagen for at bruge det som en kredsløbsudløser i dyrkede kødcellinjer [1] .
"Syntetisk biologi giver ingeniører midlerne til at implementere kredsløb for nemt og præcist at justere udtrykket af flere gener for at... eliminere potentielle negative off-target bivirkninger." - npj Systems Biology and Applications [6]
Disse valg opretter de stammespecifikke kredsløb i afsnit 3.
sbb-itb-ffee270
3. Kredsløbsstrategier for myogen, adipogen og forholdskontrolleret differentiering
3.1 Myogene kredsløb, der adskiller ekspansion fra terminal differentiering
Når kredsløbsarkitekturen er fastlagt, er den næste opgave stammespecifik implementering. For myogenese er hovedproblemet enkelt at formulere, men svært at udføre: celler skal først proliferere og derefter skifte til differentiering, når det kræves, uden at drive for tidligt i nogen retning.
En Tet-On-MyoD kassette er en af de mest direkte måder at gøre dette på. I denne opsætning prolifererer celler under standardbetingelser, når doxycyclin (DOX) er fraværende. Tilføj DOX, og kredsløbet driver myogen forpligtelse.Forskere ved flere kinesiske institutioner brugte denne tilgang i kylling embryonale fibroblaster og rapporterede effektiv myotubedannelse efter induktion [4] .
En bistabil toggle switch tilbyder strammere tilstandskontrol. Systemer bygget fra gensidigt hæmmende repressorer som E-KRAB og Pip-KRAB kan omdanne en kort DOX-puls til et stabilt myogent program [6] . I praksis betyder det, at den differentierede tilstand forbliver låst efter induktion i stedet for at falme, når signalet fjernes. Tilføjelse af negativ autoregulering hjælper også med at reducere stokastisk støj og begrænse læk differentiering under ekspansionsfasen [6].
Dette er vigtigt, fordi standard in vitro myogen differentiering ofte går i stå halvvejs. Fusionsindekser er normalt kun omkring 50–60%, hvilket efterlader en stor ikke-fusionerende reservepopulation [3]. Kredsløbsdrevet MyoD-aktivering kan forbedre engagement, men engagement alene garanterer ikke ensartet fusion. Når MyoD-kredsløb kombineres med MEK, NOTCH og RXR-modulation, bliver næsten fuldstændig fusion i 2D-kultur mulig [3]. Kredsløbet styrer timingen af linjeindtræden; de små molekyler fremmer en renere og mere ensartet cellecyklus-exit.
3.2 Adipogen og fibrogen kontrol for sammensætning og struktur
Muskel alene er ikke nok. Når myogen kontrol er på plads, er det næste problem sammensætning: hvor meget fedt der dannes, hvor meget ECM der deponeres, og hvornår disse programmer aktiveres. Her er binær TIL/FRA-kontrol ofte for grov. Hvad teams normalt har brug for, er gradueret output, især omkring det adipogene-fibrogene forgreningspunkt.
miSFITs giver en praktisk måde at justere udtryk i trin.Ved at placere muterede miRNA-målsteder - for eksempel, steder for miR-17 - i 3′UTR af outputgener som PPARG eller BMP4, kan forskere vælge udtryksniveauer fra et variantbibliotek [5]. Det gør adipocytinduktion mere som en lysdæmper end en lyskontakt. I stedet for at skubbe celler ind i en alt-eller-intet respons, kan teams titrere adipogenese mere omhyggeligt [5].
Fibroblaster er heller ikke bare tilskuere her. De leverer ECM-proteiner, der former tekstur [10]. Det gør fibrogenisk kontrol til en del af produktdesign, ikke bare et sideemne. Kredsløb kan hjælpe med at styre skiftet mellem fibrogeniske og adipogene tilstande, og i dyrket fjerkræ kan direkte PPARG-aktivering i fibroblaster være nødvendig for at generere meningsfuld fedtaflejring [10].
En butterfly-arkitektur passer godt til dette problem, fordi den holder sensing og output adskilt. Sensing-laget læser cellens nuværende tilstand, mens output-laget justerer PPARG, CEBPA eller andre linjeregulatorer. Denne adskillelse hjælper med at forhindre, at adipogene eller bindevævsprogrammer aktiveres, før cellerne har nået det rette udviklingsstadie.
3.3 Multi-linje ratio kontrol og feedback sensing
Ratio-kontrol kredsløb håndterer et andet problem: ikke om differentiering sker, men om den endelige populationssammensætning forbliver, hvor den skal. For hele skårne produkter er det lige så vigtigt at få muskler, fedt og ECM i de rette proportioner som at få en hvilken som helst linje til at differentiere.
Disse systemer bygger feedback kontrol ind i cellerne selv. Tilstands-specifikke promotorer begrænser signalproteinudtryk til celler, der allerede har forpligtet sig til en given linje.Syntetiske parakrine moduler lader derefter forpligtede myogene celler frigive et hæmmende signal, der undertrykker adipogen forpligtelse i nærliggende celler. Logikken ligner lateral inhibering i Delta-Notch udviklingssystemer [1] [6]. Hvor forgrening bliver mere kompliceret, kan multi-input porte kombinere ekstracellulære signaler med interne tilstandssignaler [9].
miSFITs fungerer også på dette stadium. Ved at justere outputstyrken af BMP4 eller andre morfogener kan teams ændre linjebalancen uden at omskrive den opstrøms beslutningslogik. I konstruerede genreguleringsnetværk øgede denne type kontrol målrettet linjedifferentieringseffektivitet fra 52% i kontroller til 81% i konstruerede iPSCs [2]. I modificerede mesenkymale stamceller drev syntetiske kredsløb hjertedifferentieringseffektiviteten til 76% [2].
Tabellen nedenfor sammenligner enkelt-linje og forholds-kontrol tilgange ved hjælp af de punkter, der er vigtigst i produktionen.
| Funktion | Enkelt-linje kredsløb | Forholdskontrol kredsløb |
|---|---|---|
| Kompleksitet | Lav; typisk en enkelt inducerbar promotor og regulator [4] | Høj; kræver logiske porte og parakrin signalering [6][9] |
| Overvågningsbyrde | Lav; følger normalt en enkelt reporter [4] | Høj; kræver sporing af flere linjemarkører [5] |
| Robusthed | Moderat; tilbøjelig til heterogenitet og reservecelleformation [3] | Høj; bruger feedback og lateral inhibering til at balancere populationer [1] |
| Produktionsværdi | Høj for biomasse; begrænset for kompleks vævsstruktur [10] | Essentiel for hele-skåret produkter, der kræver muskel, fedt og ECM [4] |
Forholdskontrolkredsløb tilføjer en tungere valideringsbelastning. Men deres indbyggede feedback er svær at matche med proceskontrol alene, hvilket lægger mere pres på klonvalg og procestest.
4. Fra konstruktion til proces: validering, opskalering og regulatorisk tilpasning
4.1 Integrationsstrategi og klonvalg for stabil ydeevne
Efter kredsløbsdesign starter den svære del: at få det kredsløb ind i celler på en måde, der forbliver stabil gennem produktion i stor skala.
Lentiviral levering er ofte effektiv og giver stabile integranter hurtigt. Men integrationen er tilfældig. Det betyder mere regulatorisk opmærksomhed, plus en risiko for, at udtrykket falder over tid på grund af nedlukning. Transposonsystemer såsom PiggyBac og Sleeping Beauty ligger i midten. De kan holde ydeevnen på tværs af mange passager, men du skal stadig screene for kopiantallet og indsættelsessteder. Sleeping Beauty, for eksempel, er blevet brugt til stabilt at immortalise bovine satellitceller ved at overudtrykke TERT og CDK4, med linjer der bevarer myogenisk potentiale ud over 40 passager [10] . CRISPR knock-in giver den strammeste kontrol over, hvor konstruktionen lander, og den mest præcise genomiske indstilling, selvom klonudvælgelse er langsommere og gennemløb er lavere.
| Integrationsmetode | Indsættelseskontrol | Stabilitet | Skalerbarhed | Regulatoriske overvejelser |
|---|---|---|---|---|
| Lentiviral levering | Lav (tilfældig integration) | Høj, men tilbøjelig til at blive tavs | Høj | Større regulatorisk kontrol på grund af tilfældig indsættelse og virale rester |
| Transposoner (PiggyBac/SB) | Moderat | Høj over mange passager | Høj | Kræver screening for kopiantal og indsættelsessteder |
| CRISPR Knock-in | Høj (sted-specifik) | Meget høj | Moderat | Gunstig; reducerer risikoen for at forstyrre endogene gener |
| Episomale Vektorer | Ingen (ekstrakromosomale) | Lav; kan gå tabt under deling | Lav | Lavere integrationsbarriere, men uegnet til langsigtet ekspansion |
Kloningsscreening skal gøre mere end at bekræfte, at konstruktionen er til stede.Det bør spore udtryksdrift, indsætningsprofil, vækstkinetik, differentieringseffektivitet og fænotypebevarelse ved højt passagenummer. snRNA-seq er nyttigt her, fordi det kan udelukke kloner beriget for Pax7⁺/Ki-67⁻ reserve celler - celler, der forlader cellecyklussen uden at differentiere - før opskalering [3]. Porcine EPSCs med et Tet-On-PAX7 kredsløb bevarede høj muskeldifferentiering i 3D mikrocarrier og suspension kultur over 40 passager [8].
4.2 Hvordan kredsløbsadfærd ændrer sig i 3D, mikrocarrier og bioreaktor kultur
Når du har en klon, er den næste test, om den opfører sig på samme måde uden for 2D. I mange tilfælde gør den ikke. Ydeevne i 2D overføres sjældent rent til suspension, mikrocarrier eller scaffold kultur, fordi diffusionsgradienter, iltbegrænsninger og shear alle ændrer kredsløbsoutput.
En af de første kontroller er inducer diffusion. I omrørte tankbioreaktorer skal småmolekyleinducere nå cellerne jævnt. I praksis kan der dannes gradienter, især i tætte mikrobærerkulturer og inde i aggregater eller stilladskerne. Suspensionskultur er normalt bedre egnet til storskalaproduktion af dyrket kød, fordi det understøtter højere celletæthed og giver strammere proceskontrol.
Overvågning af cellestatus bliver også sværere, når systemet skaleres. Fluorescerende reportersignaler, der er nemme at aflæse ved mikroskopi i 2D, kan blive svære at opløse i uigennemsigtige 3D-konstruktioner. Fluorescerende timere - prober, der skifter emission fra blå til rød, når et protein modnes - kan give realtidsdata om kredsløbsaktivering in situ [1]. Den fornuftige rute er iscenesat validering: først i 2D, derefter i 3D-konstruktioner, og til sidst under de endelige bioreaktorbetingelser [3] [8].
4.3 Karakterisering, fødevaresikkerhedsdokumentation og UK/EU-overvejelser
Efter procestestning skal karakterisering vise, at kredsløbsfunktion, fænotype og sikkerhed stadig holder. Den centrale datastak bør inkludere flowcytometri, qPCR med cm-sekvenser, RNA-seq tidsforløb og funktionelle aflæsninger såsom myosin tung kæde arealfraktion og myoglobin ekspression [1]. Optimerede serumfrie differentieringsmedier har vist sig at bringe myoglobin ekspression til cirka 30% af niveauerne fundet i oprindelig bovin muskel [3]. Det giver teams en klar benchmark fremfor et vagt mål.
Du skal også dokumentere protein-, aminosyre- og fedtprofiler samt sensoriske egenskaber [10][3].
Fra et regulatorisk synspunkt trækker Storbritannien og EU en klar linje mellem spontant immortaliserede (ikke-GMO) cellelinjer og genetisk konstruerede linjer. Sidstnævnte kræver bredere sikkerhedsdossierer [10][3]. Stabilitetspakker skal vise fænotypebevaring og genomisk stabilitet på tværs af hele produktionsbroen - fra mastercellebank til endelige produktionsceller - og sporbarhedsoptegnelser skal redegøre for hver passage imellem [10]. Hvis kredsløbet afhænger af en kemisk inducer, er et fødevaresikkert eller licenseret additiv som vanillinsyre at foretrække frem for doxycyclin [1].
Rutinemæssig genomovervågning er et must, og en inducerbar selvmords- eller eliminationskontakt bør dokumenteres som en central risikokontrolforanstaltning [7]. Dens funktion bør også inkluderes i sikkerhedsdossieret, især da UK og EU-regler for dyrket kød fortsætter med at tage form.
5. Praktisk køreplan og konklusion
5.1 En faseopdelt implementeringskøreplan for dyrkede kødhold
Den reneste vej fra koncept til produktion er et trinvis arbejdsforløb.
Fase 1 er design. Start med at definere den målrettede linje, brug derefter snRNA-seq til at bekræfte de vigtigste flaskehalse, før du vælger en kredsløbsarkitektur. Dette trin er vigtigt, fordi et kredsløb kun kan løse de begrænsninger, du faktisk har identificeret.
Fase 2 er konstruktion og 2D-validering. Byg konstruktionen og kontroller, at kredsløbet opfører sig som forventet i 2D ved hjælp af en simpel reporteraflæsning.På dette stadium er målet ligetil: bekræft, at logikken fungerer, før der skiftes til sværere og mere omkostningstunge modeller.
Fase 3 er skaleringsrelevant stresstestning. Skift til 3D-systemer og bioreaktor-relevante forhold, og sammenlign derefter output med 2D-baseline. Det er her, mange design begynder at vise deres svage punkter, især når masseoverførsel, skære- og matrixeffekter kommer i spil.
Fase 4 er regulerings- og sikkerhedsintegration, og den bør køre parallelt med Fase 3. Sikkerheds- og reguleringsarbejde bør ikke vente til slutningen. Kør det sideløbende med opskalering, inklusive dokumentation for enhver inducerbar sikkerhedsmodul.
5.2 Indkøb af muliggørende værktøjer og materialer gennem Cellbase
Når arbejdsgangen er fastlagt, bliver indkøb ofte den hastighedsbegrænsende faktor.
- cellelinjer
- serumfri og kemisk definerede medier
- stilladser
- bioreaktorkomponenter
- sensorer
- analytisk udstyr
Pålidelig adgang til kompatible materialer på hvert trin har en direkte effekt på, hvor hurtigt kredsløbsadfærd kan karakteriseres under skaleringsrelevante forhold.
5.3 Vigtige punkter
Syntetiske genkredsløb giver dyrket kød-teams programmerbar kontrol over timing, tærskler og linjebalance, som medie-only protokoller ikke kan matche. Arkitekturvalg former reversibilitet, lækage og sikkerhed.Inducerbare systemer foretrækkes normalt, fordi de giver betinget kontrol og en lavere metabolisk belastning [6].
"Det syntetiske biologi-værktøjssæt kan bruges til at etablere cellelinjer med justerbar genekspression, som, når det kombineres med PAT og computermodellering, kan muliggøre lukkede kontrolsystemer til at levere optimal produktudbytte og kvalitet." - npj Systems Biology and Applications [6]
Succesfuld implementering er ikke kun et biologiproblem. Det afhænger af tæt kobling mellem kredsløbsingeniørarbejde, bioprocesdesign, regulatorisk dokumentation og indkøb.
Ofte stillede spørgsmål
Hvordan forbedrer syntetiske genkredsløb differentieringskonsistens?
Syntetiske genkredsløb kan gøre differentiering mere konsistent, fordi de giver dig programmeret kontrol over celleadfærd og linjeforpligtelse.I praksis betyder det at bruge modulære logiske operationer til at justere gen- og transkriptionsfaktoreksponering med præcis timing.
Den timing er vigtig. Det hjælper celler med at bevæge sig gennem definerede tilstandsændringer i den rigtige rækkefølge, i stedet for at drive ind i blandede eller uønskede skæbner. Det reducerer også off-target differentiering og mindsker støj på tværs af kulturen.
Resultatet er ligetil: mere ensartede, stabile og modne cellepopulationer til produktion af dyrket kød.
Hvilket kredsløbsdesign passer til myogen eller adipogen kontrol?
I forskning i dyrket kød kan de samme kyllingefibroblaster skubbes ind i enten linje. Myogenese følger et sæt induktionsprotokoller, mens adipogenese kan aktiveres ved at udsætte cellerne for input som kyllingeserum eller fedtsyrer.
Derfra kan disse celle skæbner kontrolleres trin for trin inde i 3D hydrogel stilladser for at bygge kødstrukturer med definerede fedt- og kollagenforhold.
Hvorfor opfører genkredsløb sig ofte anderledes i 3D-kultur?
I 3D-kultur, opfører genkredsløb sig ofte anderledes, fordi celler håndterer fysiske og strukturelle input, der simpelthen ikke eksisterer i 2D monolag. Disse input inkluderer mekanisk spænding, forskydningsspænding, matrixstivhed og lokal celletæthed.
Disse signaler kan ændre signalveje som Notch. De kan også ændre, hvordan syntetiske kredsløb registrerer kraft og koordinerer nedstrøms reaktioner, herunder celle-celle adhæsion og vævsmorfogenese.









