

Leikkausjännitys voi joko edistää tai estää viljellyn lihan tuotantoa. Miksi? Koska käytetyiltä soluilta puuttuvat suojaavat seinämät, mikä tekee niistä alttiita vaurioille bioreaktoreiden nestevoimien vaikutuksesta. Tämä artikkeli syventyy siihen, miten leikkausjännitys vaikuttaa näihin soluihin, millaisia raja-arvoja ne kestävät ja miten suunnitella järjestelmiä, jotka suojaavat niitä.
Keskeiset havainnot:
- Leikkausjännitys syntyy nesteen liikkeestä ja voi vahingoittaa hauraita eläinsoluja aiheuttaen kalvovaurioita, irtoamista tai kuolemaa.
- Suurin osa nisäkässoluista sietää 0,3–1,7 Pascalia, mutta jopa alhaisemmat tasot voivat aktivoida stressivasteita.
- Suunnitteluratkaisut, kuten juoksupyörän tyyppi, ilmastusmenetelmät ja bioreaktorin geometria vaikuttavat suoraan leikkausvoimiin.
- Strategiat vaurioiden minimoimiseksi sisältävät lempeämpien bioreaktorisuunnitelmien käytön (e.g. , ilmankuljetus- tai keinutusjärjestelmät), sekoitusnopeuksien optimoinnin ja suojaavien aineiden, kuten Pluronic F68. lisäämisen.
Viljellyn lihan osalta tämän tasapainon hallinta on kriittistä, jotta solut kasvavat ja erilaistuvat vahingoittumatta, erityisesti tuotannon laajentuessa. Tutkitaan näiden kynnysten taustalla olevaa tiedettä ja käytännön ratkaisuja bioreaktorin suunnitteluun.
110: Pyöriminen kuin Maa: Matalan leikkausvoiman bioreaktorien suunnittelu parempaan soluviljelyyn Olivier De...
Mikä vaikuttaa leikkausjännitykseen bioreaktoreissa
Bioreaktoreiden leikkausjännitykseen vaikuttavien tekijöiden ymmärtäminen on ratkaisevan tärkeää olosuhteiden optimoimiseksi, erityisesti kun kyseessä ovat herkät solut. Sukelletaanpa tärkeimpiin tekijöihin, jotka muokkaavat sen voimakkuutta ja jakautumista.
Bioreaktorin suunnittelu ja käyttöolosuhteet
Bioreaktorin suunnittelu vaikuttaa merkittävästi siihen, missä ja miten leikkausjännitys syntyy. Yksi keskeinen tekijä on käytetty juoksupyörän tyyppi.Esimerkiksi Rushton-turbiinit voivat luoda energian hajontanopeuksia, jotka ovat jopa 280 kertaa suurempia kuin astian keskiarvo, kun taas korkean hyötysuhteen aksiaalivirtausjuoksupyörät, kuten HE3, tuottavat nopeuksia, jotka ovat lähempänä 180 kertaa keskimääräistä hajontaa [4]. Muut suunnitteluelementit, kuten juoksupyörän halkaisija, nopeus ja sijainti, vaikuttavat myös energian jakautumiseen.
Mielenkiintoista on, että ilmastus aiheuttaa paljon kovempia voimia kuin sekoitus. Kun pienet kuplat (1–2 mm) puhkeavat, ne vapauttavat energiamääriä välillä 10⁷–10⁹ W/m³, mikä voi tappaa yli 1 000 solua yhdellä kertaa [4]. Tämä tekee kuplien käyttäytymisestä kriittisen huomion kohteen, erityisesti viljellyn lihan tuotannossa.
Virtausohjaimet ovat toinen keskeinen suunnitteluelementti. Ne estävät pyörteen muodostumisen viljelmässä, mikä muuten vetäisi kuplia nesteeseen ja lisäisi puhkeamistapahtumia pinnalla [4]. Lisäksi juoksupyörän ja astian halkaisijasuhde sekä juoksupyörän korkeus pohjasta vaikuttavat siihen, miten energia leviää bioreaktorissa.
Leikkausjännityksen epätasainen jakautuminen
Leikkausjännitys ei jakaudu tasaisesti bioreaktorissa. Tutkimukset osoittavat, että energian hajaantuminen keskittyy tiettyihin alueisiin, kuten juoksupyörän purkausalueelle, jälkivirtauksiin ja nesteen pinnalle, jossa kuplat rikkoutuvat. Nämä kuumat pisteet voivat aiheuttaa haasteita mittakaavan suurentamisessa.
Weiwei Hu Biogen Idec korostaa tätä mittakaavaongelmaa:
Käsitys 'leikkausherkkyydestä' on historiallisesti asettanut mielivaltaisen ylärajan sekoitukselle ja ilmastukselle bioreaktorin toiminnassa; kuitenkin, kun solutiheydet ja tuottavuudet jatkavat kasvuaan, massansiirtovaatimukset voivat ylittää nämä mielivaltaiset alhaiset rajat [4].
Esimerkiksi vuonna 2021 Junxuan Zhangin ja Xueliang Lin tekemä tutkimus Jiangnanin yliopistosta vertaili 250 ml:n pyörivää pulloa ja 20 m³:n sekoitettua säiliöreaktoria käyttäen laskennallista virtausdynamiikkaa. He havaitsivat, että jopa alhaisimmilla sekoitusnopeuksilla suuremman reaktorin leikkausvoimat olivat riittävän voimakkaita irrottamaan soluja mikrokantajista, ja ilmastus aiheutti vielä enemmän stressiä kuin sekoitus [3].
Kulttuurimuoto ja leikkausherkkyys
Kulttuurimuoto määrittää myös, miten solut kokevat leikkausjännityksen. Mikrokantajilla kasvatetut solut ovat erityisen haavoittuvia. Jos voimakas sekoitus tai kantajien väliset törmäykset aiheuttavat solujen irtoamisen, nämä solut menetetään käytännössä [4]. Toisaalta, hybridomasolujen suspensioviljelmät ovat osoittaneet kestävyyttä, säilyttäen elinkelpoisuuden jopa 1 500 RPM:n sekoitusnopeuksilla baffled-bioreaktoreissa ilman ilma-neste rajapintaa [4].
Eri viljelyjärjestelmät käsittelevät leikkausvoimia eri tavoin. Kiinteäpetibioreaktorit minimoivat leikkausvoimat pitämällä solut paikallaan kiinteillä pinnoilla, kun taas fluidisoidut petibioreaktorit aiheuttavat kohtalaista tai korkeaa leikkausta mikrokantajien liikkeen ja ylöspäin suuntautuvan nestevirtauksen kautta [2]. Jotkut mikrokantajat, erityisesti huokoiset, tarjoavat sisäpintoja, jotka voivat suojata soluja äärimmäisiltä voimilta, tarjoten paremman suojan verrattuna kiinteisiin mikrokantajiin [2]. Nämä erot korostavat tarvetta tasapainottaa ravinteiden toimitus ja soluvaurioiden riski bioreaktoreita suunniteltaessa.
Leikkausjännityksen Kynnykset Eri Solutyypeille
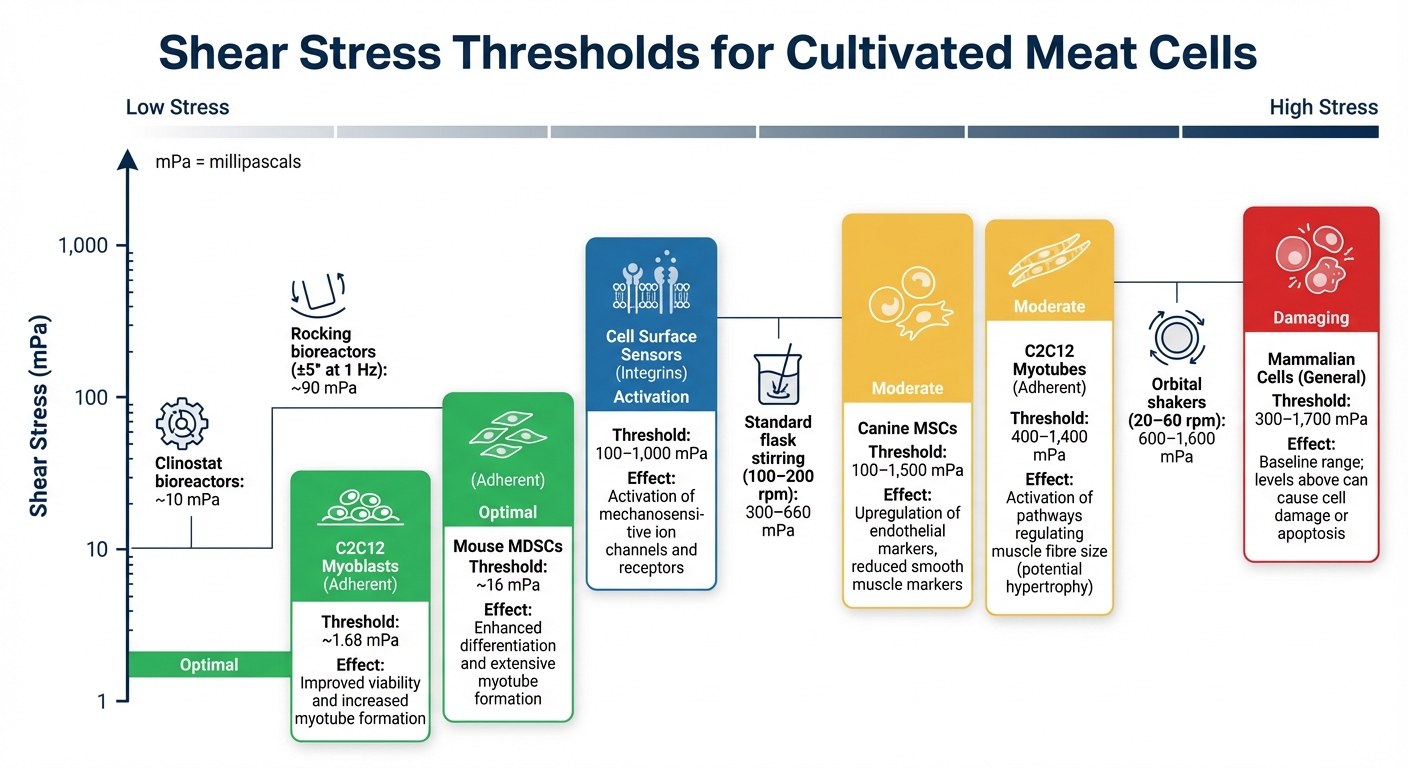
Leikkausjännityksen Sietokyvyn Kynnykset Viljellyille Lihasolutyypeille
Leikkausjännityksen hallinta on kriittistä viljellyn lihan tuotannossa, sillä epätasainen jännitys voi vahingoittaa soluja, joilta puuttuu vahvat soluseinät. Ymmärtämällä kunkin solutyypin sietämät erityiset jännitystasot autetaan ylläpitämään solujen terveyttä, laukaisemaan mekaanisesti herkkiä vasteita tai edistämään erilaistumista.
Kynnysarvot Yleisille Solutyypeille
Leikkausjännityksen sietokyky vaihtelee merkittävästi solutyyppien välillä, ja näiden kynnysten tunteminen on avainasemassa bioreaktorin asetusten hienosäätämisessä.
Esimerkiksi viljellyt lihasmyoblastit, kuten C2C12-linja, menestyvät matalassa leikkausjännityksessä. Syklinen jännitys, noin 1.68 mPa , parantaa myotubien muodostumista ja fuusiota [8] . Hiirilihaksista peräisin olevat kantasolut (MDSC:t) osoittavat parempaa myogeenista erilaistumista ja laajempaa myotubusten muodostumista altistuessaan 16 mPa [8] . Kun myoblastit kypsyvät myotubuksiksi, ne voivat käsitellä korkeampia stressitasoja; pulssitettu stressi välillä 400 mPa ja 1,400 mPa aktivoi reittejä, jotka säätelevät lihaskuitujen kokoa, mikä voi johtaa hypertrofiaan [8] .
Mesenkymaaliset kantasolut (MSC:t) reagoivat myös ainutlaatuisesti. Esimerkiksi, kun koiran MSC:t altistuvat leikkausjännitykselle välillä 100 mPa ja 1,500 mPa, ne ylireguloivat endoteelimerkkejä kuten PECAM-1 ja VE-cadherin samalla kun alireguloivat sileän lihaksen merkkejä [10] .
Leikkausjännityksen kynnysarvojen vertailutaulukko
Tässä on nopea vertailu leikkausjännityksen kynnysarvoista eri viljeltyjen lihasolutyyppien välillä:
| Solutyyppi | Leikkausjännityksen kynnysarvo (mPa) | Havaitut vaikutukset | Lähde |
|---|---|---|---|
| Nisäkässolut (yleinen) | 300–1,700 | Perustaso; tätä korkeammat tasot voivat johtaa soluvaurioihin tai apoptoosiin | [1] |
| C2C12 Myoblastit (kiinnittyneet) | ~1.68 | Parantunut elinkelpoisuus ja lisääntynyt myotubusten muodostuminen | [8] |
| Hiiren MDSC:t (Adherent) | ~16 | Parantunut erilaistuminen ja laaja myotubusten muodostuminen | [8] |
| C2C12 Myotubes (Adherent) | 400–1,400 | Lihaskuitujen kokoa säätelevien reittien aktivointi (mahdollinen hypertrofia) | [8] |
| Koiran MSC:t | 100–1,500 | Endoteelimerkkien yliregulaatio, vähentyneet sileän lihaksen merkit | [10] |
| Solupinnan sensorit (integriinit) | 100–1,000 | Mekanoherkkien ionikanavien ja reseptorien aktivointi | [1] |
Kontekstina, kulttuurin sekoittaminen 100–200 rpm nopeudella standardipullossa tuottaa leikkausjännitystasoja 300–660 mPa, kun taas kiertävät ravistimet, jotka toimivat 20–60 rpm nopeudella, tuottavat suurempia voimia, jotka vaihtelevat 600 mPa:sta 1,600 mPa:han [1]. Hellävaraisemmat järjestelmät, kuten keinuvat bioreaktorit (±5° taajuudella 1 Hz), aiheuttavat stressiä noin 90 mPa [9], ja klinostaatin bioreaktorit toimivat noin 10 mPa, pysyen selvästi alle mekaanisesti herkkien solupintatunnistimien aktivointikynnyksen [1].
Nämä kynnykset toimivat ohjeena bioreaktorin olosuhteiden säätämisessä, auttaen ylläpitämään optimaalisia ympäristöjä skaalaus- ja solukasvuvaiheiden aikana.
sbb-itb-ffee270
Kuinka vähentää leikkausstressivaurioita
Leikkausstressivaurioiden minimointi viljellyn lihan tuotannossa on tasapainon saavuttamista. Tavoitteena on varmistaa tehokas sekoitus ja hapen toimitus samalla kun suojellaan herkkiä soluja mekaanisilta vaurioilta. Tämä edellyttää älykästä bioreaktorin suunnittelua ja harkittuja operatiivisia strategioita.
Bioreaktorin suunnittelumuutokset
CFD (Computational Fluid Dynamics) -mallinnuksen käyttö on keskeinen askel bioreaktorin suorituskyvyn optimoinnissa. Nykyaikaiset CFD-tekniikat sisältävät nyt monivaihevirtauksen simuloinnin, joka ottaa huomioon solujen ja mikrokantajien väliset vuorovaikutukset. Tämä johtaa tarkempiin arvioihin leikkausjännityksestä ja sen mahdollisista vaurioista [5].
Bioreaktorin tyyppi vaikuttaa merkittävästi leikkausjännityksen tasoihin. Vaikka sekoitussäiliöreaktoreita käytetään edelleen laajalti, vaihtoehtoiset suunnitelmat voivat tarjota lempeämpiä olosuhteita:
- Ilmanvaihtobioreaktorit: Nämä poistavat mekaaniset sekoittimet ja käyttävät sen sijaan kaasun aiheuttamaa kiertoa mekaanisen leikkausjännityksen vähentämiseksi [5].
- Aaltoliike- tai keinubioreaktorit: Luottaen pintaliikkeeseen potkurien sijasta, nämä ovat ihanteellisia matala- tai keskipitoisille viljelmille, jotka vaativat hellävaraista sekoitusta [5].
- Pystypyöräbioreaktorit: Erityisen tehokkaita aggregaattipohjaisille viljelmille, nämä ovat osoittautuneet menestyksekkäiksi solujen elinkelpoisuuden ylläpitämisessä ihmisen iPSC-aggregaattien laajentamisen aikana [11].
Toinen tärkeä tekijä on solususpensioiden ei-Newtonilainen käyttäytyminen. Esimerkiksi seerumia sisältävät suspensiot osoittavat leikkausohentavia ominaisuuksia, joita perinteiset mallit eivät usein pysty kuvaamaan. Kehittyneitä malleja, kuten Sisko-mallia, käyttämällä saadaan tarkempia ennusteita leikkausjännityksestä, mikä auttaa hienosäätämään mekaanisia voimia ja välttämään kynnysarvoja, jotka voisivat muuttaa geneettistä ilmentymistä [6].
Solujen kylvö- ja sekoitusmenetelmät
Toimintastrategiat ovat myös merkittävässä roolissa leikkausjännityksen aiheuttamien vaurioiden vähentämisessä. Esimerkiksi ajoittainen sekoitus solujen kiinnittymisen alkuvaiheessa voi rajoittaa leikkausaltistusta samalla kun varmistetaan ravinteiden tehokas jakautuminen. Sekoituksen säätäminen vaatii huolellista harkintaa tekijöistä, kuten seerumin pitoisuus, solutiheys ja viljelmän ikä [6].
Sekoitusnopeuksia määritettäessä CFD-mallinnus voi auttaa löytämään ihanteellisen tasapainon - riittävä hapensiirto ilman mekaanista vahinkoa. Osastoidut simulaatiot voivat edelleen hienosäätää leikkausjännityksen jakautumista, mikä tekee prosessista tehokkaamman [5].
Vaikutus bioreaktorin suunnitteluun ja skaalaamiseen
Kun bioreaktoreita skaalataan kasvatetun lihan tuotantoa varten, leikkausjännityksen kynnysarvojen ymmärtäminen ja soveltaminen on kriittistä. Nämä kynnysarvot vaikuttavat päätöksiin sekoittimen nopeudesta, ilmastimen suunnittelusta ja muista parametreista solujen elinkelpoisuuden varmistamiseksi tuotantomäärien kasvaessa.
Bioreaktorin käyttöparametrien asettaminen
Leikkausjännityksen kynnysarvot ovat keskeisessä roolissa operatiivisten rajojen määrittelyssä. Esimerkiksi hematopoieettisilla kantasoluilla (HSC) on noin 0.092 Pa[12]. Tämän tason alapuolella pysyminen - kuten toiminta 50 rpm:llä, joka tuottaa noin 0.068 Pa - tukee terveellistä solujen laajenemista, saavuttaen 27.4-kertaisen kasvun. Kuitenkin, sekoituksen nostaminen 100 rpm:ään lisää leikkausjännitystä noin 0.192 Pa:han, mikä johtaa 72% apoptoosin määrään ja rajoittaa laajenemisen 24.5‐kertainen[12].
"HSC-solujen lisääntymisen ja toiminnan kynnysleikkausjännityksen on raportoitu olevan 0,092 Pa." – Hosseinizand et al. [12]
Leikkausvaurioita syntyy, kun turbulentit pyörteet pienenevät noin kahteen kolmasosaan solun tai aggregaatin halkaisijasta[12][13]. 50 rpm:ssä pyörteet ovat noin 280 µm, mikä on turvallista soluille. Mutta 100 rpm:ssä pyörteet kutistuvat 166 µm:iin, mikä lisää mekaanisten vaurioiden riskiä.
Ilmastus lisää hydrodynaamista rasitusta. Pienet kuplat (1 mm halkaisija) tuottavat paikallisia nesteen nopeuksia noin 6,4 m/s puhkeamisen aikana, kun taas suuremmat 6 mm kuplat tuottavat lempeämpiä huippuja 0,94 m/s[13]. Tämän vastapainoksi käytetään lisäaineita, kuten Pluronic F68, estämään solujen tarttumista kuplien pintoihin.Kuitenkin niiden tehokkuus riippuu oikean pitoisuuden ylläpitämisestä suhteessa kaasun pinta-alaan[13] .
Nämä parametrit ovat olennaisia siirryttäessä suurempiin bioreaktorijärjestelmiin.
Olosuhteiden ylläpitäminen skaalausvaiheessa
Skaalaus 250 ml:n pyörivästä pullosta 20 m³:n sekoitettuun säiliöreaktoriin tuo mukanaan ainutlaatuisia haasteita. Pienimuotoisten järjestelmien hydrodynaamiset olosuhteet eivät suoraan siirry teollisiin tilavuuksiin. Suurten reaktoreiden käyttö jopa minimisekoittamisnopeuksilla voi aiheuttaa leikkausvoimia, jotka ovat riittävän voimakkaita irrottamaan soluja mikrokantajista[3].
"Vaikka sekoitusnopeus olisi lähellä Njs:ää, pelkästään juoksupyörien aiheuttama leikkaus voi aiheuttaa solujen irtoamisen mikrokantajista, ja vielä enemmän hydrodynaamista stressiä aiheutuu ilmastuksen kautta." – Zhang et al.[3]
Jotta leikkausolosuhteet pysyisivät johdonmukaisina skaalausprosessin aikana, yksi lähestymistapa on pitää juoksupyörän kärjen nopeus vakiona. Tämä voi kuitenkin johtaa pidempiin sekoitusaikoihin sekä ravinteiden ja hapen gradienttien muodostumiseen, mikä voi vaikuttaa negatiivisesti solujen kasvuun ja suorituskykyyn[3]. Laskennallinen virtausdynamiikka (CFD) -mallinnus on olennaista stressivyöhykkeiden tunnistamiseksi ja reaktorin suunnittelun optimoimiseksi skaalausprosessin aikana[5].
Solulinjoille, jotka ovat erittäin herkkiä leikkaukselle, vaihtoehtoiset reaktorisuunnitelmat ovat usein sopivampia. Ilmanvaihtoreaktorit, jotka poistavat mekaaniset sekoittimet, on onnistuneesti mallinnettu jopa 300 000 litran tilavuuksille, saavuttaen teoreettisia solutiheyksiä 2 × 10⁸ solua/mL[7]. Samoin, keinuvat bioreaktorit käyttävät lempeitä aaltoliikkeitä vähentääkseen leikkausvoimia, mikä tekee niistä tehokkaita siemenlinjoille jopa 500 L asti[14][15]. Alustat kuten
Yhteenveto ja suositukset
Leikkausjännityksen hallinta tehokkaasti on ratkaisevan tärkeää solujen elinkelpoisuuden ja tuottavuuden ylläpitämiseksi viljellyn lihan tuotannossa. Tutkimukset osoittavat, että kuplien rikkoutuminen ilmastuksen aikana luo vahingollisempia voimia kuin mekaaninen sekoitus. Esimerkiksi pienet kuplat (1 mm) tuottavat nesteen nopeuksia 6,4 m/s rikkoutuessaan, kun taas suuremmat kuplat (6 mm) tuottavat lempeämpiä huippuja 0,94 m/s [13] . Vähentääkseen näitä voimia hankintatiimien tulisi keskittyä bioreaktoreihin, jotka on varustettu sintratuilla mikrospargereilla (15 μm huokoskoko), jotka mahdollistavat pulssitetun ilmastuksen ja vähentävät kaasu-neste rajapintaa. Nämä seikat ovat elintärkeitä bioreaktorijärjestelmien skaalaamisessa.
Toinen tärkeä tekijä on pyörteiden mittakaavan ja solun halkaisijan suhde (η/d_c), joka voi auttaa vähentämään sekoittamisesta aiheutuvaa vahinkoa. Elokuussa 2017 Bioprosessitekniikan ja farmaseuttisen teknologian instituutti teki tästä tutkimuksen. Käyttäen 3 litran Applikon lasibioreaktoria Sf21 hyönteissoluilla, he osoittivat, että kuusilapainen Rushton-juoksupyörä 205 rpm nopeudella, yhdistettynä 199 μm kupliin, tuotti GFP-proteiinisaannon 12,75 μg/mL. Sen sijaan kaltevalapainen juoksupyörä 171 rpm nopeudella, joka tuotti suuremman spesifisen kaasupinta-alan 18,0 m²/m³, tuotti vain 4,0 μg/mL [13]. Tämä osoittaa, että kaasun kokonaispinta-ala on vaikuttavampi kuin sekoitusnopeus.
Suojaavat aineet, kuten Pluronic F68 (0,5–3 g/L), voivat muodostaa 16–40 μm suojaavan kerroksen kuplien ympärille, estäen soluja kiinnittymästä [13]. Kuitenkin, kuten Tobias Weidner ja kollegat havaitsivat:
Jos [kaasun kokonais]pinta-ala ylittää tietyn kynnyksen, Pluronicin pitoisuus ei enää riitä solujen suojaamiseen [13].
Tämä tarkoittaa, että insinöörien on tarkasti seurattava kaasun pinta-alaa suhteessa Pluronic F68 -pitoisuuteen skaalausprosessin aikana varmistaakseen, että solut pysyvät suojattuina.
Herkille solulinjoille vaihtoehtoiset reaktorisuunnitelmat voivat tarjota ratkaisuja. Esimerkiksi ilmankohotusreaktorit poistavat mekaaniset sekoittimet, luoden lempeämmän sekoitusympäristön [7]. Kiinteäpetibioreaktorit ovat toinen vaihtoehto, jotka pystyvät ylläpitämään erittäin matalia seinämän leikkausjännityksiä, jotka vaihtelevat 10⁻³ - 10⁻² Pa [17]. Erikoistuneita matalan leikkausjärjestelmiä tutkiville tiimeille, toimittajat kuten
Lisäksi, naudan myoblastien pitäminen alle 25 populaatiokaksinkertaistumisen on olennaista niiden erilaistumiskyvyn säilyttämiseksi [16]. Tämän kynnyksen ylittäminen voi johtaa fuusioindeksin laskuun noin 6.81% jokaisella siirroksella [16], vähentäen solujen kykyä muodostaa lihassäikeitä. Tämän ratkaisemiseksi prosessi-insinöörien tulisi käyttää laskennallista virtausdynamiikkaa (CFD) mallinnusta tunnistaakseen korkean leikkausjännityksen alueet ennen siirtymistä laboratoriosta teollisiin järjestelmiin. Tämä lähestymistapa varmistaa sujuvammat siirtymät ja paremmat tulokset skaalausvaiheessa.
UKK
Kuinka mittaan leikkausjännitystä bioreaktorissani?
Leikkausjännitystä bioreaktoreissa arvioidaan usein käyttämällä laskennallisia mallinnustekniikoita kuten laskennallista virtausdynamiikkaa (CFD). Nämä menetelmät mahdollistavat virtauskuvioiden analysoinnin ja leikkausalueiden tunnistamisen bioreaktorissa. Lisäksi pienikokoiset leikkaustestaustyökalut ovat hyödyllisiä arvioitaessa, kuinka herkkiä tietyt solulinjat ovat, ja arvioitaessa erilaisia prosessiolosuhteita. Jatkuvaa seurantaa varten leikkausjännitys voidaan määrittää laskemalla nesteen nopeus ja viskositeetti. Tämä lähestymistapa on erityisen tehokas mikrofluidisissa järjestelmissä tai käyttämällä online-leikkausjännityslaskureita.
Mikä ilmastusmenetelmä minimoi kuplien rikkoutumisvauriot?
Kuplien rikkoutumisvaurioiden minimointi perustuu vahvasti pienempien kuplien käyttöön. Nämä kuplat aiheuttavat vähemmän soluvaurioita, kun niitä verrataan tilavuus-tilavuus -perusteella.Vaikka tarkkoja tekniikoita ei ole esitetty, kuplien koon ja käyttäytymisen hallinta - kuten niiden koon säätely - on ratkaisevan tärkeää repeämisen haitallisten vaikutusten vähentämisessä.
Mitä minun pitäisi pitää vakiona, kun skaalaan ylöspäin leikkausvoiman vähentämiseksi?
Kun kasvatetaan viljellyn lihan bioreaktoreiden kokoa, on tärkeää pitää leikkausjännitys alle noin 3 Pa solujen vahingoittumisen estämiseksi. Kiinnitä erityistä huomiota tekijöihin, kuten sekoitus, virtauksen mallit, ja ilmastus varmistaaksesi, että leikkaustasot pysyvät tasaisina koko toiminnan ajan.









