

Jos poistan seerumin mutta pidän saman villityypin solulinjan, en voi odottaa, että pelkkä väliaineen säätö estäisi vanhenemista, ajautumista tai kiinnittymisen menetystä. Tässä artikkelissa osoitan, että seerumiton menestys viljellyssä lihassa riippuu yleensä molemmista järjestelmän puolista: määritellystä väliaineesta solun ulkopuolella ja soluissa tehdyistä muokkauksista, jotka auttavat sitä jakautumaan, pysymään kiinnittyneenä ja säilyttämään myogeenisen toiminnan.
Bioprosessien insinööreille ja soluviljelytiimeille ydinkohdat ovat yksinkertaisia:
- Seerumiton väliaine muuttaa solujen käyttäytymistä, ei vain ainesosaluetteloita. Glukoosin, glutamiinin, glysiinin ja kystiinin otto voi muuttua seerumittomissa olosuhteissa.
- Primaariset satelliittisolut saavuttavat solulinjan rajat aikaisin. Villityypin porsassolut menettävät usein myogeeniset piirteensä noin passage 10.
- CDKN2A knockout on yksi selkeimmistä esimerkeistä artikkelissa: muokatut sian satelliittisolulinjat laajennettu 15+ passagen, ajan säilyttivät >90% elinkelpoisuuden , ja joissakin klooneissa osoittivat ~194-kertaisesti korkeampaa PAX7 tasoa passagessa 20 kuin villityypin kontrollit.
- On olemassa kompromissi. Parempi laajentuminen ei takaa myöhäisen passagen erilaistumista; jotkut muokatut linjat osoittivat silti heikompaa erilaistumista passagessa 30.
- Validointi on suoritettava tuotantoasetelmassa: sama seerumiton väliaine, sama viljelytapa ja samat mittaukset kasvulle, jätteen kertymiselle, linjamerkinnöille ja fuusiolle.
Lyhyt versio: jos haluat seerumittoman prosessin, joka voidaan siirtää tuotantoon, käsittelisin mediadesignin, geenimuokkaukset, klooniseulonnan ja bioreaktorin sovituksen yhtenä yhdistettynä työnkuluna, en neljänä erillisenä tehtävänä.
| Kohdealue | Mitä tarkistaisin ensin | Miksi se on tärkeää |
|---|---|---|
| Solusyklin hallinta | CDKN2A, passage life, PAX7 | Auttaa osoittamaan, voiko linja laajentua ilman ennenaikaista vanhenemista |
| Kasvusignaalit | IGF1R, EGFR, FGFR vaste | Seerumittomat järjestelmät tarjoavat vähemmän ulkoista signaalitukea |
| Stressin sietokyky | Elinkelpoisuus, apoptoosin merkkiaineet, leikkausvaste | Seerumin poisto ja passaging voivat johtaa solujen menetykseen |
| Ravinteiden käsittely | Glukoosin käyttö, laktaatti, ammoniakki, aminohappojen otto | Nopeampi otto voi myös tarkoittaa nopeampaa jätteen kertymistä |
| Identiteetin säilyttäminen | PAX7, MYOD, MYOG, fuusioindeksi | Nopeasti kasvava linja ei ole hyödyllinen, jos se ei enää muodosta kohdekudosta |
Käyn sitten läpi, missä seerumiton viljely epäonnistuu, mitkä muokkaukset liittyvät näihin epäonnistumiskohtiin ja miten validoisin muokatun linjan ennen prosessin siirtoa.
CRISPR-Cas Genominmuokkaus Nisäkässolulinjoissa | Protokollan Esikatselu
Pääasialliset Biologiset Esteet Seerumittomassa Viljelyssä
Seerumin poistaminen paljastaa kolme pullonkaulaa: signaalointi, kiinnittyminen ja solun identiteetti. Ongelmat alkavat solun sisällä, eivät vain väliaineen koostumuksessa. Tämä on tärkeää, koska se vaikuttaa siihen, mihin tiimit käyttävät aikaa: väliaineen säätöön, geenimuokkaukseen tai molempien yhdistelmään.
| Ominaisuus | Seerumilla täydennetty viljely | Seerumiton viljely |
|---|---|---|
| Proliferaatio | Vahva; tukee monipuoliset kasvutekijät | Vaihtelee; altis replikaatiovanhentumiselle ja G1/S-pysähdykselle |
| Kiinnittyminen | Tukee seerumiperäiset ECM-proteiinit (fibronetiini, vitronektiini) | Vaatii ulkoisia pinnoitteita tai lisäaineita; irtoamisriski kasvaa |
| Ravinteiden kuljetus | Helpottuu kantajaproteiinien, kuten albumiinin ja transferriinin, avulla | Riippuu vähemmän puskuroituneesta otosta; vaatii optimoidut ITS-X- ja lipidipitoisuudet |
| Apoptoosiriski | Matala; PI3K-AKT- ja MAPK-ERK-reitit ovat voimakkaasti aktivoituneet | Korkeampi; herkkyys oksidatiiviselle stressille ja aineenvaihdunnan jätteille kasvaa |
| Identiteetin vakaus | Yleisesti vakaa varhaisista keskivaiheisiin | Korkea fenotyyppisen muutoksen riski; kantasolumerkit usein vähenevät nopeasti |
Kasvu- ja selviytymissignaalien menetys
Kun seerumi poistetaan, kasvutekijöiden tasot laskevat jyrkästi.Solut menettävät sitten suuren osan ulkoisesta tuesta, joka auttaa pitämään PI3K-AKT- ja MAPK-ERK-aktiivisuuden korkeana. Käytännössä tämä tarkoittaa enemmän apoptoosia ja heikompaa proliferaatiota, mikä on suora ongelma laajentamisessa.
Adheesio, Ravinteiden Otto ja Stressipullonkaulat
Seerumi tekee muutakin kuin ruokkii soluja. Se toimittaa myös ECM-proteiineja, jotka tukevat kiinnittymistä ja leviämistä. Ilman fibronetiiniä, vitronetiiniä ja vastaavia tekijöitä primaariset satelliittisolut irtoavat todennäköisemmin ja siirtyvät apoptoosiin, erityisesti bioreaktorin olosuhteissa. ROCK-inhibitio Y-27632 voi auttaa jossain määrin, mutta se ei poista kiinnittymisongelmaa.
Ravinteiden käsittely vaikeutuu myös. Ilman seerumin kantajaproteiineja glukoosin, glutamiinin, glysiinin ja kystiinin otto muuttuu vähemmän puskuroituneeksi [1]. Samaan aikaan aineenvaihdunnan jätteet, kuten ammoniakki ja laktaatti, voivat kerääntyä ja estää kasvua [3]. Joten vaikka perusväliaine näyttäisi paperilla hyvältä, kuljetus ja jätteiden tasapaino voivat silti muodostua rajoittavaksi tekijäksi.
Fenotyyppinen ajautuminen seerumittoman sopeutumisen aikana
Seerumiton sopeutuminen voi valikoida alaryhmiä, jotka sietävät uusia olosuhteita, mutta eivät enää vastaa tuotteen spesifikaatiota. Se on ansa: solut voivat laajentua hyvin, mutta menettää kyvyn muodostaa aiottua kudosta.
Sarjapassagoinnin aikana merkkiaineet, kuten PAX7, MYOD, ja MYOG voivat heikentyä [2]. Seuraa linjamerkkiaineita sopeutumisen aikana, jotta ajautuminen havaitaan aikaisin eikä vasta pitkän väliaineen optimointijakson jälkeen. Nämä ovat reittejä, jotka geenieditoinnin on vakautettava.
Geenieditointimenetelmät, jotka parantavat seerumittoman suorituskyvyn
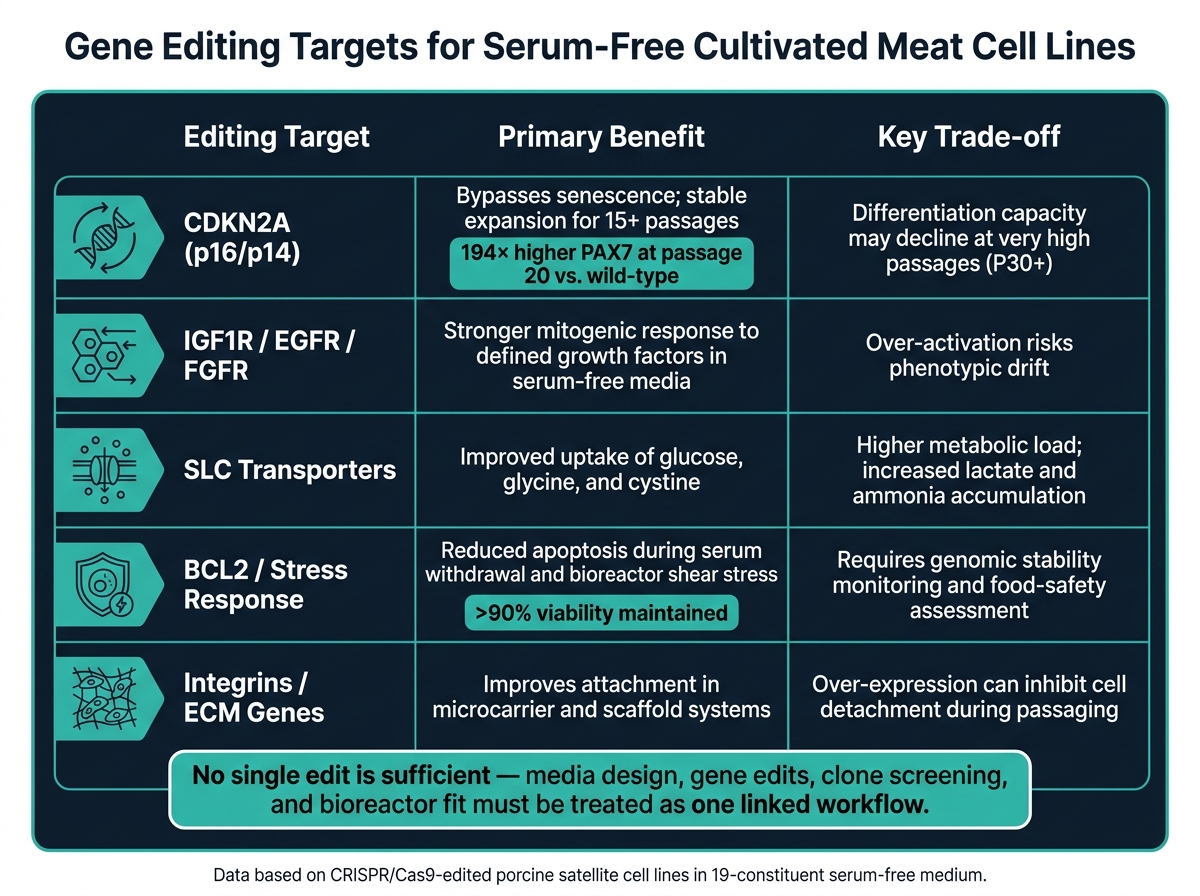
Geenieditointikohteet seerumittomassa soluviljelyssä: Hyödyt vs. kompromissit
Nämä esteet jakautuvat kolmeen editointiluokkaan: signaalinvälitys, selviytyminen ja erilaistuminen.
Kasvutekijäsignaalinvälityksen ja ravinteiden hyödyntämisen editointi
Yksi suora reitti on IGF1R, EGFR, ja FGFR ylösreguloida tai herkistää, jotta solut reagoivat voimakkaammin IGF-1:een, EGF:ään ja bFGF:ään [2]. Tämä on tärkeää seerumittomassa väliaineessa, jossa kasvutekijätasot ovat yleensä alhaiset suunnittelun vuoksi. Jos signaalinvälitys paranee, solusyklin hallinnasta tulee usein seuraava pullonkaula.
Solusyklin säätelijä CDKN2A erottuu tässä.CRISPR/Cas9-knockout exonista 2 tuotti CDKN2A −/− sian satelliittisolulinjoja, jotka laajenivat voimakkaasti yli 15 passagen ajan 19-komponenttisessa seerumittomassa väliaineessa. Tietyissä klooneissa PAX7 -ilmentyminen lisääntyi noin 194-kertaiseksi passage 20:ssa verrattuna villityypin kontrolliin [2].
Muokkaukset liuotinkantaja (SLC) kuljettajiin voivat auttaa välttämään otto-rajoituksia glukoosin, glutamiinin, glysiinin ja kystiinin skaalausvaiheessa [1]. Mutta siinä on koukku. Korkeampi otto johtaa myös nopeampaan laktaatin ja ammoniakin kertymiseen, joten kuljettajamuutokset on suunniteltava yhdessä väliaineen vaihdon ja jätteenhallinnan kanssa alusta alkaen. Pelkät ottomuutokset eivät riitä.
Solujen selviytymisen ja vastustuskyvyn parantaminen seerumittomalle stressille
Seerumin poisto, rutiinipassaus ja bioreaktorin leikkaus altistavat solut korkeammille apoptoositasoille.BCL2-reitin muokkaaminen - joko lisäämällä eloonjäämistä edistäviä jäseniä tai tukahduttamalla pro-apoptoottisia - voi vähentää solujen menetystä näiden siirtymien aikana. Tämä on vieläkin merkityksellisempää mikrokantajärjestelmissä, joissa solut kohtaavat sekä kiinnittymis- että mekaanista stressiä.
Kaikki muokkaukset, jotka parantavat eloonjäämistä tai pidentävät lisääntymistä, tarvitsevat genomisen vakauden tarkistuksia koko valmistusprosessin ajan.CDKN2A −/− porsaan satelliittisolut säilyttivät elinkelpoiset solumäärät yli 90% jatkuvan seerumittoman lisääntymisen aikana [2]. Siitä huolimatta tiimien tulisi tarkistaa kromosomaalinen eheys tietyin väliajoin sen sijaan, että olettaisivat vakauden jatkuvan.
Adhesion, Proliferation, and Differentiation Capacityn tasapainottaminen
Vaikein osa on hallita laajentumisen ja erilaistumisen välistä vetovoimaa.CDKN2A knockout säilyttää myogeenisen potentiaalin 10. passagen ajan, kun taas villityypin solut seerumittomissa olosuhteissa menettävät lähes täysin myogeeniset ominaisuutensa. Fuusioindeksit 16.3% - 56.3% raportoitiin editoiduissa linjoissa [2] . 30. passagen kohdalla, kuitenkin, jopa editoidut solut voivat osoittaa heikentyvää erilaistumiskykyä [2].
| Muokkauskohde | Pääasiallinen hyöty seerumittomassa viljelyssä | Tärkeä kompromissi |
|---|---|---|
| CDKN2A (p16/p14) | Ohittaa solujen vanhenemisen; vakaa laajeneminen yli 15 passagen ajan [2] | Differentiatiokyky voi heikentyä erittäin korkeilla passageilla (P30+) [2] |
| IGF1R / EGFR / FGFR | Vahvempi mitogeeninen vaste määritellyille kasvutekijöille [2] | Yliaktivoituminen voi aiheuttaa fenotyyppistä muutosta |
| SLC-kuljettajat | Parantunut glukoosin, glysiinin ja kystiinin otto [1] | Korkeampi metabolinen kuormitus; lisääntynyt laktaatin ja ammoniakin kertyminen [1] |
| BCL2 / stressivaste | Vähentynyt apoptoosi vieroituksen ja leikkausrasituksen aikana [2] | Vaatii genomin vakauden seurantaa ja elintarviketurvallisuuden arviointia [2] |
| Integraatiot / ECM-geenit | Parantaa kiinnittymistä mikrokantaja- ja tukijärjestelmissä [2] | Yli-ilmentyminen voi estää solujen irtoamisen passagoinnin aikana [2] |
Kiinnittymismuutokset ovat hyödyllisimpiä mikrokantaja- tai tukijärjestelmissä.Heitä on parempi käsitellä formaattikohtaisina työkaluina, ei ratkaisuna jokaiseen seerumittomaan prosessiin.
Indusoitavat CRISPR-järjestelmät antavat tiimeille käytännöllisen tavan käsitellä laajentumis- ja erilaistumiskauppaa. Idea on yksinkertainen: käytä indusoitavia muokkauksia erottamaan laajentumisvaihe erilaistumisesta.
Yksikään näistä muokkauksista ei ole merkityksellinen, jos fenotyyppi ei säily aiotussa seerumittomassa väliaineessa.
sbb-itb-ffee270
Muokatun solulinjan rakentaminen ja validointi seerumittomaan viljelyyn
Oikean muokkauksen löytäminen on vain osa työtä. Vaikeampi osa on muuttaa tuo muokkaus vakaaksi solulinjaksi, joka kestää seerumittoman valmistuksen. Se vaatii tiukan työnkulun, joka yhdistää muokkauksen, kloonivalinnan ja validoinnin yhdeksi putkeksi. Ja tuon putken tulisi suoraan testata jo tunnistettuja signaalointi-, selviytymis- ja kiinnittymisrajoituksia.
Muokkaustyökalujen ja toimitustapojen valinta
Kohteille, kuten CDKN2A, CRISPR/Cas9 knockout on käytännöllinen ensimmäinen askel, kun tavoitteena on poistaa solusyklin estäjä ja tukea pitkäaikaista laajentumista [2]. Ensisijaisissa karjasoluissa yleisiä toimitusreittejä ovat ei-virukselliset transfektiomenetelmät, kuten Lipofectamine, ja virukselliset järjestelmät, kuten lentiCRISPR v2 [2][4] . Ennen siirtymistä kloonaustyöhön, varmista toimitustehokkuus.
Yksi asia on tärkeämpi kuin joskus saa tunnustusta: tarkista jokainen klooni täsmälleen siinä väliaineessa ja viljelytilassa, joka on suunniteltu tuotantoa varten. Jos valmistusprosessi käyttää määriteltyä seerumitonta väliaineetta, staattista adherenttiviljelyä, mikrokantajia tai muuta järjestelyä, se on se olosuhde, johon solujen tulisi kohdata seulonnan aikana.
Muokattujen solujen seulonta tuotannon seerumittomassa formuloinnissa
Yleinen tapa on eristää klooneja rajoittavalla laimennuksella ja sitten vahvistaa muokkaus Sanger-sekvensoinnilla kohdelokuksessa [2]. Kun muokkaus on vahvistettu, seulontaa tulisi jatkaa samassa seerumittomassa formuloinnissa ja viljelytilassa, joka on tarkoitettu valmistukseen [2][1].
Tässä vaiheessa mittaa perusasiat, jotka kertovat, voiko klooni elää prosessin kanssa sen sijaan, että se vain selviäisi muokkauksesta:
- Kasvu
- Elinkelpoisuus
- Glukoosin kulutus
- Maitohapon tuotanto
- Ammoniakin kertyminen
On myös järkevää lisätä PAX7 RT-qPCR aikaisin, koska kantasolujen menetys voi ilmetä ennen kuin linja epäonnistuu ilmeisemmällä tavalla [1][2].
Muokattujen solujen karakterisointi ennen prosessin siirtoa
Ennen prosessin siirtoa validoinnin tulisi kattaa neljä toisiinsa liittyvää aluetta: genomin muokkaus, reittivaste, passagen stabiilisuus ja toiminta. Jokainen vastaa eri ongelmaan. Genomiset tarkastukset käsittelevät fenotyyppisen ajautuman riskiä. Käytetyn kasvatusliuoksen analyysi osoittaa ravinteiden ottoa ja jätteen kertymisen rajoja.Fuusioindeksi kertoo, onko myogeeninen erilaistuminen edelleen olemassa [2][1].
| Testityyppi | Mitä se mittaa | Miksi se on tärkeää seerumittomille viljellyille lihalinjoille |
|---|---|---|
| T7 Endonukleaasi I / Sanger-sekvensointi | Editointitehokkuus ja tarkka genominen sekvenssi | Vahvistaa onnistuneen geenin poiston tai lisäyksen ennen laajentamista [2] |
| RT-qPCR (PAX7, MYOD, MYOG, BAX, CCND1) | Kantasolujen, erilaistumisen ja apoptoosin merkkiaineiden transkriptiotasot | Seuraa solujen terveyttä ja erilaistumispotentiaalia pitkän aikavälin viljelyssä [2][4] |
| Immunofluoresenssi (MyHC / CK18) | Linjaspesifinen proteiini-ilmentyminen | Varmistaa, että solut säilyttävät lihas- tai epiteeli-identiteetin muokkauksen ja sopeutumisen jälkeen [2][4] |
| Käytetyn kasvatusliuoksen analyysi | Glukoosi-, aminohappo-, laktaatti- ja ammoniakkiprofiilit | Määrittää ravintoaineiden tarpeet ja ohjaa bioreaktorin syöttöstrategiaa [1] |
| Fuusioindeksi | Monitumaisiin myotubeihin sisällytettyjen tumien prosenttiosuus | Vahvistaa, että myogeeninen erilaistumiskyky säilyy ilman seerumia [2] |
| Tekstuuriprofiilianalyysi (TPA) | 3D-rakenteiden kovuus, kimmoisuus ja pureskeltavuus | Vahvistaa, että muokatut solut tuottavat lopputuotteen, jolla on lihamaiset fysikaaliset ominaisuudet [2] |
Genominen validointi perustuu T7 Endonuclease I -testiin sekä yksittäisten kloonien Sanger-sekvensointiin [2]. Reitin vahvistus käyttää RT-qPCR:ää tai Western blot -menetelmää osoittamaan, että suunniteltu transkripti tai proteiinimuutos on todella tapahtunut, mukaan lukien merkkiaineet kuten PAX7, MYOD , MYOG ja MyHC [2][4].
Pitkäaikainen vakaus , tavoitteena on 15-30 passagen vertailuarvo, jossa toistuvat tarkastukset kasvusta, elinkelpoisuudesta ja merkkiaineiden ilmentymisestä. CDKN2A knockout -possun satelliittisolut säilyttivät elinkelpoiset solumäärät yli 90% 15 passagen ajan seerumittomissa olosuhteissa, mutta erilaistumiskyky alkoi heikentyä 30 passagen kohdalla [2].
Toiminnallinen testaus esittää sitten yksinkertaisimman kysymyksen: ovatko nämä yhä ne solut, joita tarvitset? Myogeenisissä linjoissa fuusioindeksi osoittaa, voivatko muokatut solut yhä muodostaa monitumaisia myotubeja ilman seerumia [2] . Tekstuuriprofiilianalyysi (TPA) tarkistaa sitten, osoittavatko 3D-rakenteet lihaa muistuttavaa kovuutta, kimmoisuutta ja pureskeltavuutta [2].
Käytä näitä tietoja kloonin siirto-olosuhteiden asettamiseen seerumittomalle valmistukselle.
Muokatuista solulinjoista seerumittomaan valmistukseen
Muokattujen solujen sovittaminen väliaineeseen ja bioreaktorin suunnitteluun
Kun klooni läpäisee validoinnin, tehtävä muuttuu. Siinä vaiheessa menestys riippuu siitä, kuinka hyvin solulinja sopii prosessiin. Lisää seulontaa ei korjaa huonoa prosessisovitusta.
Käytetyn väliaineen analyysin tulisi ohjata glukoosin lisäyksiä, aminohappojen täydentämistä ja kasvutekijöiden annostelua, mukaan lukien määritellyt syötteet, kuten bFGF ja IGF-1 [2]. Adherenttisissa järjestelmissä tukirakenteen kylvötiheys ja tarttumisikkuna - noin 2 tuntia ennen bioreaktoriin siirtoa - tulisi asettaa muokatun linjan kiinnittymiskäyttäytymisen perusteella, ei seerumia sisältävien protokollien perusteella [2]. Näiden tietojen tulisi sitten suoraan vaikuttaa päätöksiin syöttöajankohdasta, kylvötiheydestä ja siirtoajankohdasta.
Muokatut linjat voivat tukea pidempää laajentumista, korkeampaa solutiheyttä ja vakaampaa merkkiaineiden ilmentymistä. Tämä tarkoittaa, että skaalaus on tehtävä muokatun linjan mitatun käyttäytymisen perusteella, ei villityypin oletusten mukaan.
Käytännössä linjan valinnasta tulee hankinta- ja skaalauspäätös, ei vain biologinen päätös.
Keskeiset huomiot T&K, tuotanto- ja hankintatiimeille
Seerumiton sopeutuminen ei ole pelkästään median formulointikysymys. Se alkaa solulinjasta, eikä pelkkä median optimointi ratkaise sitä.Kohdennettu geenieditointi, erityisesti solusyklin repressoreiden, kuten CDKN2A, poistaminen käsittelee taustalla olevaa biologiaa, joka aiheuttaa primaaristen satelliittisolujen epäonnistumisen seerumittomissa olosuhteissa. CDKN2A−/− porsaan satelliittisolut ylläpitivät PAX7-ilmentymistä noin 194-kertaisena villityypin kontrollisoluihin verrattuna 20. passaasissa ja saavuttivat fuusioindeksin jopa 56.3% 10. passaasissa - vaiheessa, jossa muokkaamattomat solut olivat suurelta osin menettäneet myogeenisen toimintansa [2].
Kehitys- ja valmistustiimien välillä jako on melko selkeä:
- R&D-tiimien tulisi rakentaa validointiputki, joka testaa muokattuja klooneja todellisissa tuotanto-olosuhteissa alusta alkaen. Tämä sisältää kasvun, ravinteiden kulutuksen, linjan vakauden ja 3D-erilaistumiskyvyn.
- Tuotantotiimien tulisi käyttää muokatun linjan ravintoprofiilia syötteen suunnitteluun ja bioreaktorin parametrien asettamiseen, koska seerumia sisältävistä protokollista kopioidut oletukset eivät todennäköisesti pidä paikkaansa [1].
- Hankintatiimien on laadittava hankintasuunnitelmat, jotka vastaavat muokatun linjan erityisvaatimuksia, mukaan lukien määritellyt kasvutekijät, lipidit, antioksidantit sekä linjan tarttumisprofiiliin sopivat tukirakenteet tai mikrokantajat.
UKK:t
Miksi pelkkä median optimointi ei riitä?
Pelkkä median optimointi ei riitä. Monissa tapauksissa eläinsoluilla ei yksinkertaisesti ole suurimittakaavaiseen tuotantoon tarvittavia ominaisuuksia, kuten leikkausjännityksen kestävyys, aineenvaihdunnan tehokkuus, ja elinkelpoisuus suuritiheyksisessä suspensiossa.
Seerumittomilla medioilla on merkitystä, mutta ne eivät korjaa solun sisäänrakennettuja rajoituksia. Nämä rajoitukset sisältävät rajoitetun lisääntymisiän, herkkyyden bioreaktorin stressille, ja erilaiset ravitsemukselliset tarpeet lajien ja kehitysvaiheiden välillä.
Mitkä geenimuokkaukset ovat tärkeimpiä seerumittomassa viljelyssä?
Viljellyn lihan tuotannossa tärkeimpiä muokkauksia ovat ne, jotka vähentävät riippuvuutta lisätyistä kasvutekijöistä. Yksi esimerkki on CDKN2A-deleetio, joka voi parantaa sian satelliittisolujen lisääntymistä ja erilaistumista seerumittomissa olosuhteissa.
Toinen reitti on suunnitella lihaksen kantasoluja indusoitavalle FGF2-yliekspressiolle ja mutantille RasG12V. Tämä järjestely tukee autokriinista signalointia ja poistaa tarpeen rekombinantille FGF2:lle väliaineessa.
Miten muokattuja solulinjoja tulisi validoida valmistusta varten?
Muokattujen solulinjojen tulisi käydä läpi genomiset, proteomiset ja toiminnalliset testit valmistuskyvyn ja erilaistumispotentiaalin vahvistamiseksi.
Käytännössä tämä tarkoittaa, että tarkistetaan, että muokkaus teki sen, mitä sen oli tarkoitus tehdä ilman että se aiheuttaa ongelmia muualla. Tutkijoiden tulisi varmistaa, että geneettiset muutokset eivät heikennä erilaistumista kohdekudoksiin ja että aiotut ominaisuudet, kuten stressinsietokyky tai seerumista riippumaton kasvu, ilmenevät odotetusti.









