

Jos voit laajentaa soluja, mutta et voi vaihtaa niitä oikeaan kohtaloon oikeaan aikaan, prosessisi pysähtyy erilaistumisessa. Tämä on ydinkohta: synteettiset geenipiirit antavat sinulle solunsisäisen hallinnan sitoutumisesta, ajoituksesta, muistista ja sukulinjan sekoituksesta, kun pelkät mediavaihtelut usein jättävät heterogeeniset, osittain sitoutuneet populaatiot.
Jos rakentaisin viljellyn lihan erilaistumisprosessin työnkulun, ottaisin tästä artikkelista heti neljä kohtaa:
- Aloita alkuperäisestä verkostosta, ei rakenteesta. Käytä snRNA-seq, reittianalyysiä, GRN-päätelmää ja miRNA-profilointia löytääksesi, missä solut pysähtyvät, ajautuvat tai haarautuvat väärään kohtaloon.
-
Sovita piirin tyyppi prosessiongelmaan.
Kytkin sopii lukitukseen, feedforward- tai kaistanpäästösuunnittelu sopii ajoituksen hallintaan, logiikkaportti sopii monisignaalin porttaukseen, ja miSFIT:t sopivat porrastettuun ulostuloon. - Suunnittele alusta alkaen vähävuotoiseksi, vähämeluiseksi ja turvalliseksi. Ortogonaaliset osat, negatiivinen autoregulaatio, iFFL:t, cm-transgeenit ja indusoitava tappo- tai kasvunestomoduuli ovat osa rakennetta, eivät jälkiajatus.
- Vahvista mittakaavaan liittyvissä olosuhteissa aikaisin. Piiri, joka toimii 2D:ssä, saattaa muuttua 3D:ssä, mikrokantajassa tai sekoitetussa suspensiossa indusoijagradienttien, happirajojen ja leikkausvoimien vuoksi.
Artikkeli tekee myös käytännöllisen huomion, joka on tärkeä prosessitiimeille: yksilinjainen hallinta ja suhdehallinta ovat eri tehtäviä. A Tet-On MyoD-kasetti voi edistää myogeenistä sisäänpääsyä, mutta kokonaisleikatut tuotteet tarvitsevat lihaksen, rasvan ja ECM:n osuuksien, hallintaa, mikä yleensä tarkoittaa palautetta, parakriinista signalointia ja raskaampaa kloonien seulontaa.
Muutama luku ankkuroi tämän viestin:
- Standardi myogeeninen erilaistuminen voi pysähtyä fuusioindeksien ollessa noin 50–60%
- Muokatut GRN:t iPS-soluissa lisäsivät kohdelinjan erilaistumista 52% tasolta 81%
- Synteettiset piirit muokatuissa MSC-soluissa ohjasivat sydämen erilaistumista 76%
- Jotkut porsaan Tet-On-PAX7 linjat säilyttivät korkean myogeenisen potentiaalin yli 40 passagen
- Noin 20% ihmisen pluripotenttisista kantasoluista saattaa kantaa syöpään liittyviä mutaatioita, minkä vuoksi indusoitavat turvamoduulit ovat tärkeitä
Synteettinen geenipiirityönkulku viljellyn lihan erilaistumiseen
Tutkijatarinat: Michael Elowitz, Geneettiset piirit elävissä soluissa
Nopea vertailu
| Aihe | Mitä artikkeli sanoo yksinkertaisesti |
|---|---|
| Pääpullonkaula | Differointi, ei laajentuminen |
| Miksi piirit auttavat | Ne lisäävät kynnysarvon hallintaa ja solutilan muistia solun sisällä |
| Paras ensimmäinen askel | Kartoitus alkuperäisen kohtalon hallinnasta snRNA-seq ja vastaavilla työkaluilla |
| Pääpiirivaihtoehdot | Kytkin, eteenpäin syöttö, kaistanpäästö, logiikkaportit, miSFITs, CRISPRa/i |
| Myogeneesin esimerkki | Tet-On-MyoD kasvun erottamiseksi lopullisesta erilaistumisesta |
| Adipogeneesi / ECM-hallinta | miSFITs ja bow-tie-suunnitelmat asteittaiselle PPARG/BMP4-tyyppiselle tuotokselle |
| Kokonaisleikkaushaaste | Suhdehallinta lihaksen, rasvan ja sidekudoksen välillä |
| Laajennusriski | 2D-käyttäytyminen ei välttämättä päde 3D:ssä tai bioreaktoreissa |
| Integraatiovalinnat | Lentivirus, transposonit, CRISPR knock-in, episomaaliset vektorit |
| Sääntelypiste | Suunnitellut linjat tarvitsevat laajemman turvallisuuspaketin; elintarviketurvalliset indusoijat, kuten vanilliinihappo, ovat suositeltavia DOX:n sijaan, jos mahdollista |
Yksinkertaisesti sanottuna: tämä ei ole vain piirikaavion suunnitteluartikkeli. Lukisin sen oppaana, joka yhdistää piiriarkkitehtuurin, sukulinjabiologian, kloonivalinnan, bioreaktorin suorituskyvyn ja UK/EU-turvallisuusdokumentaation yhdeksi erilaistumisstrategiaksi.
Lue eteenpäin, jos haluat täydellisen polun alkuperäisestä verkon kartoituksesta rakenteen valintaan, skaalaustarkastuksiin ja sääntelyyn sopivuuteen.
2. Suunnitteluperiaatteet erilaistumisen ohjauspiireille
2.1 Kartoitus alkuperäisestä solun kohtalon verkosta ennen piirin suunnittelua
Ennen piirin suunnittelua tarvitset selkeän kuvan siitä, mitä solu jo tekee.
Yksittäisen ytimen RNA-sekvensointi (snRNA-seq) on hyvä paikka aloittaa. Se voi näyttää hiljaiset alipopulaatiot, mukaan lukien varasolut, jotka on merkitty NOTCH2 ja HEYL, ja osoittaa reittikohteisiin, jotka voivat parantaa erilaistumista [3].
Siitä eteenpäin, trajektorianalyysi ja geenien säätelyverkoston (GRN) päättely auttavat kartoittamaan säätelijöiden aktivoinnin järjestystä ja korostavat, missä solut todennäköisimmin pysähtyvät. Myogeneesissä pääkaskadi kulkee MYOD1 ja MYOG kautta. Adipogeneesissä pääsolmut ovat PPARG ja CEBPA, fibro-adipogeenisten progenitorien (FAP) kohtalon toimiessa päähaarakohdan riskinä. Alla oleva taulukko tiivistää pääsäätelijät ja pullonkaulat.
| Sukulinja | Avaimet säätelytekijät | Kriittiset signaalireitit | Tunnistetut pullonkaulat |
|---|---|---|---|
| Myogeeninen | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Reservisolujen muodostuminen (lepotila) |
| Adipogeeninen | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Fibro-adipogeeninen progenitorin (FAP) kohtalo |
| Pluripotentti | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Spontaani erilaistuminen / heterogeenisyys |
Toinen hyödyllinen kerros on miRNA-ilmentymäprofiili. Endogeeniset miRNA:t, kuten miR-302a, liitettynä pluripotenssiin, ja miR-375, liitettynä erilaistumiseen, voivat toimia sisäisinä luokittelijoina aisti-ja-vastaa-suunnitelmissa. Sen avulla piiri voi lukea solun todellisen tilan sen sijaan, että se riippuisi vain ulkoisista indusoijista [5].
Nämä pullonkaulat pitäisi ohjata piirin valintaa. Jos pääongelma on ajautuminen, saatat tarvita lukituspiirin. Jos ajoitus on ongelma, pulssi voi sopia paremmin. Jos kohtalon hallinta riippuu useammasta kuin yhdestä vihjeestä, monisyöttölogiikka on yleensä järkevämpää.
2.2 Valitse oikea piiriarkkitehtuuri
Tässä kohtaa kompromissit tulevat selkeästi esiin. Oikea arkkitehtuuri riippuu kolmesta käytännön seikasta: kuinka pysyvä sitoutumisen täytyy olla, kuinka tarkasti ajoitusta täytyy hallita ja kuinka paljon geneettistä kuormaa solu voi kantaa ilman ongelmia.
Kaksivakaa kytkin sopii tapauksiin, joissa sukulinjan sitoutumisen täytyy pysyä lukittuna. Pääongelma on transkription aiheuttama spontaani tilan vaihtelu.
Kaistanpäästösuodattimet sopivat tapauksiin, joissa transkriptiotekijää tarvitaan vain määritellyn kehitysvaiheen aikana. Haasteena on, että indusoijan tasoja on hallittava tarkasti, muuten ajoitus voi lipsua.
Logiikkaportit lisäävät spesifisyyttä vaatimalla useampaa kuin yhtä syötettä samanaikaisesti. Esimerkiksi erilaistuminen voidaan sallia vain, kun eksogeeninen indusoija on läsnä ja solu osoittaa oikean endogeenisen miRNA-profiilin. Tämä auttaa vähentämään riskiä kohteen ulkopuolisesta sitoutumisesta.
Alla oleva taulukko esittelee tärkeimmät arkkitehtuurit ja niihin liittyvät kompromissit.
| Arkkitehtuuri | Käännettävyys | Aikaprecisio | Integraation monimutkaisuus | Pääasiallinen käyttötapaus | Keskeinen riski |
|---|---|---|---|---|---|
| Kaksivakaa kytkin | Matala (kun lukittu) | Kohtalainen | Kohtalainen | Pysyvä sukulinjan sitoutuminen | Spontaani kääntyminen melun vuoksi |
| Kaistanpäästösuodatin | Korkea (pitoisuudesta riippuva) | Korkea | Korkea | Väliaikaiset kehitysvaiheet | Vaatii tiukkaa indusoijan hallintaa |
| Logiikkaportti (JA/TAI/EI) | Vaihteleva | Kohtalainen | Kohtalainen–korkea | Solutyyppikohtainen aktivointi | Vuotavuus POIS-tiloissa |
| Rusetti / monisyöte | Korkea | Kohtalainen | Kohtalainen | Monisignaalinen integrointi | Perustuu endogeenisen miRNA:n stabiilisuuteen |
| miSFITs | Korkea | Kohtalainen | Matala–kohtalainen | Asteittainen lähtötuloksen säätö | Kapea dynaaminen alue, jos huonosti viritetty |
"Vähentämällä laskennallisten kerrosten määrää samalla säilyttäen toiminnallisuuden, tämä strategia käsittelee skaalautuvuuden esteitä geenipiirien suunnittelussa." - Nature Communications [9]
Jokainen lisätty sääntelykerros lisää kuormitusta ja kuluttaa solun resursseja. Käytännössä, jos kaksi suunnitelmaa tekevät saman työn, yksinkertaisempi on yleensä parempi vaihtoehto, kun mittakaavalla on merkitystä.
Kun arkkitehtuuri on asetettu, seuraava tehtävä on varmistaa sen kestävyys alhaisella vuotavuudella, melunvaimennuksella ja vikasietoisella ohjauksella.
2.3 Rakenna luotettavuuden, alhaisen vuotavuuden ja turvallisuuden varmistamiseksi
Piirin on pysyttävä vakaana pitkän viljelyn ajan. Lyhyt hyvä suorituskyky ei riitä tuotantokäyttöön.
Ortogonaaliset osat ovat ensimmäinen puolustus. Promoottorit, transkriptiotekijät ja sääntelyelementit, jotka eivät ristiinreagoi alkuperäisen koneiston kanssa, auttavat rajoittamaan kohteettomia vaikutuksia ja vähentämään mahdollisuutta, että endogeeniset signaalit kytkevät piirin päälle tai sammuttavat sen.Muokatut korkean tiiviyden promoottorit, kuten PCREm, on käytetty vähentämään perusvuotoa indusoitavissa nisäkässoluissa [6] .
Negatiivinen autoregulaatio on myös syytä lisätä, jos mahdollista. Se on yksi tunnetuimmista motiiveista transkriptionaalisen kohinan vähentämiseksi ja lineaarisemman vasteen tuottamiseksi indusoijan pitoisuuteen [6]. Inkoherentit eteenpäin suuntautuvat silmukat (iFFL:t) voivat lisätä toisen kontrollikerroksen suodattamalla stokastisia vaihteluita, jolloin solut reagoivat jatkuviin signaaleihin lyhyiden kohinapiikkien sijaan.
Kodonimuokatut (cm ) synteettisten transkriptiotekijöiden versiot helpottavat myös karakterisointia. Ne mahdollistavat piirin ohjaaman ilmentymisen erottamisen endogeenisestä genomisesta (g) ilmentymisestä validoinnin aikana [1]. Se saattaa kuulostaa pieneltä yksityiskohdalta, mutta se säästää aikaa, kun yrität selvittää, tuleeko lukema piiristä vai isäntägenomista.
Turvallisuusmoduulit ovat pakollisia. Noin 20% ihmisen pluripotenttisista kantasoluista kantaa syöpään liittyviä mutaatioita [7] . Joten, jos piiri menee kantasoluperäiseen linjaan, sen tulisi sisältää indusoitava kasvunpysäytys- tai eliminointimoduuli. Vanilliinihappo on hyödyllinen indusoija, jota kannattaa priorisoida tässä, koska se on lisensoitu elintarvikelisäaine, mikä tukee sen käyttöä piirin laukaisijana viljellyissä lihassolulinjoissa [1].
"Synteettinen biologia tarjoaa insinööreille keinot ottaa käyttöön piirejä, joilla voidaan helposti ja tarkasti säätää useiden geenien ilmentymistä... poistaakseen mahdolliset haitalliset kohteen ulkopuoliset sivuvaikutukset." - npj Systems Biology and Applications [6]
Nämä valinnat asettavat sukulinjakohtaiset piirit kohdassa 3.
sbb-itb-ffee270
3. Piiristrategiat myogeeniselle, adipogeeniselle ja suhdeohjatulle erilaistumiselle
3.1 Myogeeniset piirit, jotka erottavat laajenemisen terminaalisesta erilaistumisesta
Kun piirin arkkitehtuuri on asetettu, seuraava tehtävä on sukulinjakohtainen käyttöönotto. Myogeneesissä pääongelma on yksinkertainen ilmaista, mutta vaikea toteuttaa: solujen on ensin lisäännyttävä, sitten siirryttävä erilaistumiseen pyydettäessä, ilman että ne ajautuvat liian aikaisin kumpaankaan suuntaan.
A Tet-On-MyoD kasetti on yksi suorimmista tavoista tehdä tämä. Tässä asetelmassa solut lisääntyvät tavanomaisissa olosuhteissa, kun doksisykliiniä (DOX) ei ole läsnä. Lisää DOX, ja piiri ohjaa myogeenisen sitoutumisen.Tutkijat useista kiinalaisista laitoksista käyttivät tätä lähestymistapaa kanan alkion fibroblasteissa ja raportoivat tehokkaasta myotubusten muodostumisesta induktion jälkeen [4].
Kaksivakaa kytkin tarjoaa tiukemman tilan hallinnan. Järjestelmät, jotka on rakennettu keskinäisesti estävistä repressoreista, kuten E-KRAB ja Pip-KRAB, voivat muuttaa lyhyen DOX-pulssin vakaaksi myogeeniseksi ohjelmaksi [6]. Käytännössä tämä tarkoittaa, että erilaistunut tila pysyy lukittuna induktion jälkeen sen sijaan, että se häviäisi signaalin poistamisen jälkeen. Negatiivisen autoregulaation lisääminen auttaa myös vähentämään stokastista kohinaa ja rajoittamaan vuotavaa erilaistumista laajennusvaiheen aikana [6] .
Tämä on tärkeää, koska tavanomainen in vitro myogeeninen erilaistuminen usein pysähtyy kesken. Fuusioindeksit ovat yleensä vain noin 50–60%, jättäen suuren fuusioitumattoman reservipopulaation [3]. Piiripohjainen MyoD-aktivaatio voi parantaa sitoutumista, mutta pelkkä sitoutuminen ei takaa tasaista fuusiota. Kun MyoD-piirit yhdistetään MEK-, NOTCH- ja RXR-modulaatioon, lähes täydellinen fuusio 2D-kulttuurissa tulee mahdolliseksi [3]. Piiri hallitsee linjan sisäänpääsyn ajoitusta; pienet molekyylit edistävät puhtaampaa ja tasaisempaa solusyklin lopetusta.
3.2 Adipogeeninen ja fibrogeneeninen hallinta koostumukselle ja rakenteelle
Pelkkä lihas ei riitä. Kun myogeeninen hallinta on paikoillaan, seuraava kysymys on koostumus: kuinka paljon rasvaa muodostuu, kuinka paljon ECM:ää kertyy ja milloin nämä ohjelmat käynnistyvät. Tässä binäärinen ON/OFF-ohjaus on usein liian karkea. Tiimit tarvitsevat yleensä asteittaista ulostuloa, erityisesti adipogeenisen-fibrogeneenisen haarautumispisteen ympärillä.
miSFITs tarjoaa käytännöllisen tavan säätää ilmentymistä askelittain.Asettamalla mutatoituja miRNA-kohdesivustoja - esimerkiksi miR-17-sivustoja - ulostulogeeneihin, kuten PPARG tai BMP4, tutkijat voivat valita ilmentymistasoja varianttikirjastosta [5]. Tämä tekee adiposyytti-induktiosta enemmän himmenninkytkimen kaltaisen kuin valokytkimen. Sen sijaan, että solut työnnettäisiin kaikki-tai-ei-mitään-vasteeseen, tiimit voivat titrata adipogeneesiä tarkemmin [5].
Fibroblastit eivät myöskään ole vain sivustakatsojia tässä. Ne toimittavat ECM-proteiineja, jotka muokkaavat tekstuuria [10]. Tämä tekee fibrogeneettisestä hallinnasta osan tuotesuunnittelua, ei vain sivuseikkaa. Piirit voivat auttaa hallitsemaan siirtymää fibrogeneettisten ja adipogeneettisten tilojen välillä, ja viljellyssä siipikarjassa suora PPARG-aktivointi fibroblasteissa voi olla tarpeen merkittävän rasvakertymän aikaansaamiseksi [10].
Rusettiarkkitehtuuri sopii tähän ongelmaan hyvin, koska se pitää aistimisen ja ulostulon erillään. Aistimiskerros lukee solun nykyisen tilan, kun taas ulostulokerros säätää PPARG:tä, CEBPA:ta tai muita linjaregulaattoreita. Tämä erottelu auttaa estämään adipogeenisten tai sidekudosohjelmien käynnistymisen ennen kuin solut ovat saavuttaneet oikean kehitysvaiheen.
3.3 Monilinjasuhteen hallinta ja palautteen aistiminen
Suhteenhallintapiirit käsittelevät eri ongelmaa: ei sitä, tapahtuuko erilaistuminen, vaan pysyykö lopullinen populaatiomix siellä missä sen pitäisi. Kokonaisleikkaustuotteissa lihaksen, rasvan ja ECM:n saaminen oikeisiin suhteisiin on yhtä tärkeää kuin minkä tahansa linjan erilaistuminen.
Nämä järjestelmät rakentavat palautesäätöä soluihin itseensä. Tilakohtaiset promoottorit rajoittavat signaaliproteiinien ilmentymisen soluihin, jotka ovat jo sitoutuneet tiettyyn linjaan.Synteettiset parakriiniset moduulit antavat sitoutuneiden myogeenisten solujen vapauttaa estävän signaalin, joka tukahduttaa lähellä olevien solujen adipogeenisen sitoutumisen. Logiikka muistuttaa lateraalista inhibitointia Delta-Notch-kehitysjärjestelmissä [1][6] . Missä haarautuminen muuttuu monimutkaisemmaksi, monisyöttöportit voivat yhdistää solunulkoisia vihjeitä sisäisten tilasignaalien kanssa [9].
miSFITit toimivat myös tässä vaiheessa. Säätämällä BMP4:n tai muiden morfogeeneiden ulostulon voimakkuutta, tiimit voivat muuttaa linjatasapainoa ilman, että ylävirran päätöksentekologiikkaa tarvitsee kirjoittaa uudelleen. Suunnitelluissa geenisäätelyverkoissa tämän tyyppinen ohjaus lisäsi kohdelinjan erilaistumistehokkuutta 52% kontrolliryhmissä 81% suunnitelluissa iPS-soluissa [2]. Muokatuissa mesenkymaalisissa kantasoluissa synteettiset piirit ohjasivat sydämen erilaistumistehokkuuden 76% [2].
Alla oleva taulukko vertaa yhden linjan ja suhdeohjauksen lähestymistapoja tuotannossa tärkeimpien seikkojen perusteella.
| Ominaisuus | Yksilinjainen piiri | Suhteensäädinpiiri |
|---|---|---|
| Monimutkaisuus | Matala; tyypillisesti yksi indusoitava promoottori ja säätelijä [4] | Korkea; vaatii logiikkaportteja ja parakriinista viestintää [6][9] |
| Seurantataakka | Matala; seuraa yleensä yhtä raportointigeeniä [4] | Korkea; vaatii useiden linjamerkkien seuraamista [5] |
| Vakaus | Kohtalainen; altis heterogeenisyydelle ja varasolu muodostumiselle [3] | Korkea; käyttää palautetta ja lateraalista inhibitointia tasapainottamaan populaatioita [1] |
| Tuotantoarvo | Korkea biomassalle; rajallinen monimutkaiselle kudosrakenteelle [10] | Välttämätön kokonaisille leikkeleille, jotka tarvitsevat lihasta, rasvaa ja ECM:ää [4] |
Suhdeohjauspiirit lisäävät raskaamman validointikuorman.Mutta niiden sisäänrakennettua palautetta on vaikea sovittaa yhteen pelkän prosessinhallinnan kanssa, mikä lisää painetta kloonivalinnalle ja prosessitestaamiselle.
4. Rakenteesta prosessiin: validointi, skaalaus ja sääntely-yhteensopivuus
4.1 Integraatiostrategia ja kloonivalinta vakaaseen suorituskykyyn
Piirisuunnittelun jälkeen alkaa vaikea osa: saada piiri soluihin tavalla, joka pysyy vakaana tuotantomittakaavan läpikuljetuksen aikana.
Lentiviruskuljetus on usein tehokasta ja antaa vakaita integraatteja nopeasti. Mutta integraatio on satunnaista. Tämä tarkoittaa enemmän sääntelyhuomiota sekä riskiä, että ilmentyminen vähenee ajan myötä hiljentymisen vuoksi. Transposonijärjestelmät kuten PiggyBac ja Sleeping Beauty sijoittuvat keskivaiheille. Ne voivat säilyttää suorituskyvyn monien läpikuljetusten ajan, mutta sinun on silti seulottava kopioluku ja insertointikohdat. Sleeping Beauty, esimerkiksi, on käytetty vakauttamaan naudan satelliittisoluja yliekspressoimalla TERT ja CDK4, ja linjat säilyttävät myogeenisen potentiaalin yli 40 passagen ajan [10] . CRISPR knock-in antaa tiukimman hallinnan siitä, mihin konstruktio sijoittuu, ja tarkimman genomisen asetelman, vaikka kloonien valinta on hitaampaa ja läpimeno on alhaisempi.
| Integraatiomenetelmä | Lisäyksen hallinta | Stabiilisuus | Skaalautuvuus | Sääntelyyn liittyvät näkökohdat |
|---|---|---|---|---|
| Lentiviruskuljetus | Matala (satunnainen integraatio) | Korkea, mutta altis hiljentymiselle | Korkea | Suurempi sääntelyvalvonta satunnaisen insertoinnin ja viruksen jäänteiden vuoksi |
| Transposonit (PiggyBac/SB) | Kohtalainen | Korkea monien passagien yli | Korkea | Vaatii kopiomäärän ja insertointipaikkojen seulontaa |
| CRISPR Knock-in | Korkea (paikkakohtainen) | Erittäin korkea | Kohtalainen | Suotuisa; vähentää riskiä häiritä endogeenisia geenejä |
| Episomaaliset vektorit | Ei mitään (ekstrakromosomaalinen) | Matala; voi kadota jakautumisen aikana | Matala | Matala integraatioeste, mutta ei sovellu pitkäaikaiseen laajentamiseen |
Kloonauksen seulonnassa on tehtävä enemmän kuin vain varmistettava, että konstruktio on läsnä.Sen tulisi seurata ilmentymän ajautumista, insertointiprofiilia, kasvukinetiikkaa, erilaistumistehokkuutta ja fenotyypin säilymistä korkealla passagenumerolla. snRNA-seq on hyödyllinen tässä, koska se voi sulkea pois kloonit, jotka ovat rikastuneet Pax7⁺/Ki-67⁻ varantosoluilla - soluilla, jotka poistuvat solusyklistä erilaistumatta - ennen laajentamista [3]. Porcine EPSC:t Tet-On-PAX7-piirillä säilyttivät korkean lihaserilaistumisen 3D-mikrokantaja- ja suspensioviljelmässä yli 40 passagen ajan [8].
4.2 Kuinka piirin käyttäytyminen muuttuu 3D-, mikrokantaja- ja bioreaktoriviljelmässä
Kun sinulla on klooni, seuraava testi on, käyttäytyykö se samalla tavalla 2D:n ulkopuolella. Monissa tapauksissa se ei käyttäydy. Suorituskyky 2D:ssä harvoin siirtyy puhtaasti suspensioon, mikrokantajaan tai tukirakenteeseen, koska diffuusiogradientit, happirajoitukset ja leikkausvoimat muuttavat piirin ulostuloa.
Yksi ensimmäisistä tarkastuksista on indusoijan diffuusio. Sekoitetuissa säiliöbioreaktoreissa pienimolekyyliset indusoijat tarvitsevat tasaisen jakautumisen soluihin. Käytännössä gradientteja voi muodostua, erityisesti tiheissä mikrokantajakulttuureissa ja aggregaattien tai tukirakenteiden ytimissä. Suspensiokulttuuri sopii yleensä paremmin suurimittakaavaiseen viljeltyyn lihaan, koska se tukee suurempaa solutiheyttä ja mahdollistaa tiukemman prosessinhallinnan.
Solujen tilan seuranta vaikeutuu myös järjestelmän laajentuessa. Fluoresoivat raporttisignaalit, jotka on helppo lukea mikroskopialla 2D:ssä, voivat olla vaikeasti erotettavissa läpinäkymättömissä 3D-rakenteissa. Fluoresoivat ajastimet - koettimet, jotka muuttavat emissiota sinisestä punaiseen proteiinin kypsyessä - voivat antaa reaaliaikaista piirin aktivointitietoa in situ [1]. Järkevä reitti on vaiheittainen validointi: ensin 2D:ssä, sitten 3D-rakenteissa, ja lopuksi lopullisissa bioreaktorin olosuhteissa [3][8] .
4.3 Karakterisointi, elintarviketurvallisuusdokumentaatio ja UK/EU-huomioitavat asiat
Prosessitestausten jälkeen karakterisoinnin on osoitettava, että piirin toiminta, fenotyyppi ja turvallisuus ovat edelleen voimassa. Ydin datakokonaisuuden tulisi sisältää virtaussytometria, qPCR cm-sekvensseillä, RNA-seq-aikakursseja ja toiminnallisia tuloksia, kuten myosiinin raskasketjun alueen osuus ja myoglobiinin ilmentyminen [1]. Optimoidut seerumittomat erilaistumismediat ovat osoittaneet nostavan myoglobiinin ilmentymisen noin 30% tasolle, joka löytyy alkuperäisestä naudanlihaksesta [3]. Tämä antaa tiimeille selkeän vertailukohdan sen sijaan, että se olisi epämääräinen tavoite.
Sinun on myös dokumentoitava proteiini-, aminohappo- ja rasvaprofiilit sekä aistinvaraiset ominaisuudet [10] [3].
Sääntelyn näkökulmasta Yhdistynyt kuningaskunta ja EU tekevät selkeän eron spontaanisti kuolemattomien (ei-GMO) solulinjojen ja geneettisesti muokattujen linjojen välillä. Jälkimmäiset tarvitsevat laajempia turvallisuusasiakirjoja [10][3]. Stabiilisuuspakettien tulisi osoittaa fenotyypin säilyminen ja genominen stabiilisuus koko tuotantosillan ajan - pääsolupankista lopullisiin tuotantosoluihin - ja jäljitettävyysasiakirjojen on katettava jokainen välivaihe [10]. Jos piiri riippuu kemiallisesta indusoijasta, elintarviketurvallinen tai lisensoitu lisäaine, kuten vanilliinihappo, on suositeltavampi kuin doksisykliini [1].
Rutiininomainen genominen valvonta on välttämätöntä, ja indusoitava itsemurha- tai eliminointikytkin tulisi dokumentoida keskeisenä riskinhallintatoimenpiteenä [7]. Sen toiminta tulisi myös sisällyttää turvallisuusasiakirjaan, erityisesti kun Yhdistyneen kuningaskunnan ja EU:n säännöt viljellylle lihalle muotoutuvat.
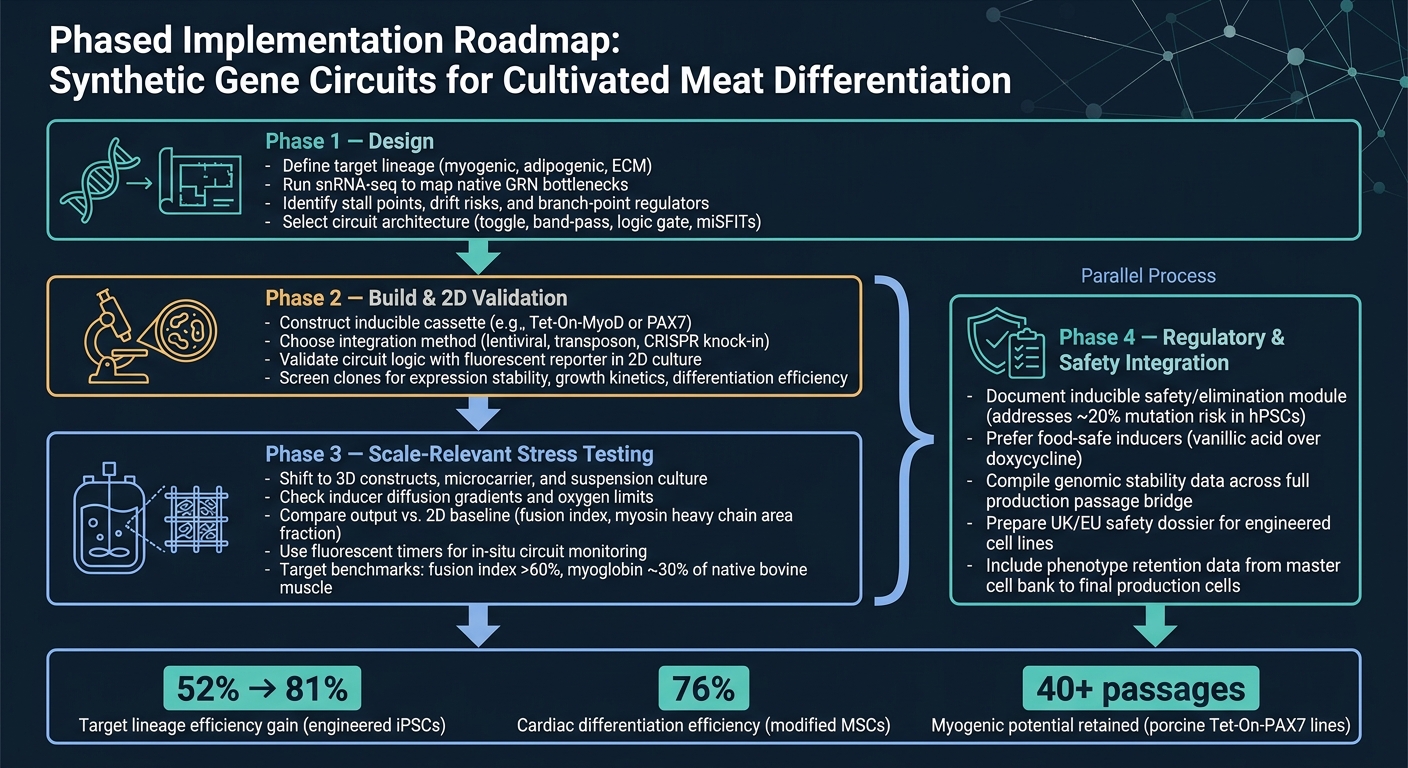
5. Käytännön tiekartta ja johtopäätös
5.1 Vaiheittainen toteutussuunnitelma viljellyn lihan tiimeille
Puhdas reitti konseptista tuotantoon on vaiheittainen työnkulku.
Vaihe 1 on suunnittelu. Aloita määrittelemällä kohdelinja, käytä sitten snRNA-seq:tä vahvistaaksesi tärkeimmät pullonkaulat ennen kuin valitset piiriarkkitehtuurin. Tämä vaihe on tärkeä, koska piiri voi ratkaista vain ne rajoitteet, jotka olet todella tunnistanut.
Vaihe 2 on rakentaminen ja 2D-validointi. Rakenna konstruktio ja tarkista, että piiri käyttäytyy tarkoitetulla tavalla 2D:ssä, käyttäen yksinkertaista raporttilukemaa.Tässä vaiheessa tavoite on yksinkertainen: vahvista logiikan toimivuus ennen siirtymistä vaikeampiin ja kalliimpiin malleihin.
Vaihe 3 on mittakaavaan liittyvä rasitustestaus. Siirry 3D-järjestelmiin ja bioreaktoreihin liittyviin olosuhteisiin, sitten vertaa tuloksia 2D-peruslinjaan. Tässä vaiheessa monet suunnitelmat alkavat paljastaa heikkoutensa, erityisesti kun massansiirto, leikkaus ja matriisivaikutukset tulevat esiin.
Vaihe 4 on sääntely- ja turvallisuusintegraatio, ja sen tulisi edetä rinnakkain Vaiheen 3 kanssa. Turvallisuus- ja sääntelytyötä ei pitäisi jättää loppuun. Suorita se rinnakkain mittakaavan kasvattamisen kanssa, mukaan lukien dokumentointi kaikille indusoitaville turvallisuusmoduuleille.
5.2 Työkalujen ja materiaalien hankinta Cellbase
Kun työnkulku on määritetty, hankinnasta tulee usein nopeutta rajoittava vaihe.
- solulinjat
- seerumittomat ja kemiallisesti määritellyt väliaineet
- tukirakenteet
- bioreaktorikomponentit
- anturit
- analyyttiset laitteet
Luotettava pääsy yhteensopiviin materiaaleihin jokaisessa vaiheessa vaikuttaa suoraan siihen, kuinka nopeasti piirikäyttäytymistä voidaan karakterisoida mittakaavaan liittyvissä olosuhteissa.
5.3 Keskeiset havainnot
Synteettiset geenipiirit antavat viljellyn lihan tiimeille ohjelmoitavan hallinnan ajoitukseen, kynnysarvoihin ja sukulinjan tasapainoon, mitä pelkät väliaineprotokollat eivät voi saavuttaa. Arkkitehtuurin valinta muokkaa palautuvuutta, vuotoisuutta ja turvallisuutta.Indusoitavat järjestelmät ovat yleensä suositumpia, koska ne tarjoavat ehdollista hallintaa ja pienemmän metabolisen kuormituksen [6].
"Synteettisen biologian työkalupakkia voidaan käyttää solulinjojen luomiseen, joissa on säädettävä geeniekspressio, mikä yhdessä PAT:n ja laskennallisen mallinnuksen kanssa voi mahdollistaa suljetun kierron ohjausjärjestelmät, jotka tuottavat optimaalisen tuotteen saannon ja laadun." - npj Systems Biology and Applications [6]
Onnistunut käyttöönotto ei ole pelkästään biologinen ongelma. Se riippuu tiukasta yhteydestä piirielementtien suunnittelun, bioprosessisuunnittelun, sääntelydokumentaation ja hankinnan välillä.
Usein kysytyt kysymykset
Kuinka synteettiset geenipiirit parantavat erilaistumisen johdonmukaisuutta?
Synteettiset geenipiirit voivat tehdä erilaistumisesta johdonmukaisempaa, koska ne antavat sinulle ohjelmoidun hallinnan solukäyttäytymiseen ja linjasitoutumiseen.Käytännössä tämä tarkoittaa modulaaristen logiikkaoperaatioiden käyttöä geenien ja transkriptiotekijöiden ilmentymisen säätämiseen tarkalla ajoituksella.
Tuo ajoitus on tärkeä. Se auttaa soluja siirtymään määritellyissä tilamuutoksissa oikeassa järjestyksessä sen sijaan, että ne ajautuisivat sekoittuneisiin tai ei-toivottuihin kohtaloihin. Se myös vähentää kohdentamatonta erilaistumista ja vähentää melua koko viljelmässä.
Hyöty on yksinkertainen: yhtenäisemmät, vakaammat ja kypsät solupopulaatiot viljellyn lihan tuotantoon.
Mikä piirikaavio sopii myogeeniseen tai adipogeeniseen kontrolliin?
Viljellyn lihan tutkimuksessa samoja kanan fibroblasteja voidaan ohjata kumpaankin linjaan. Myogeneesi noudattaa yhtä induktioprotokollien sarjaa, kun taas adipogeneesi voidaan käynnistää altistamalla solut syötteille, kuten kanan seerumille tai rasvahapoille.
Siitä lähtien näitä solujen kohtaloita voidaan hallita vaihe vaiheelta 3D-hydrogeelirakenteissa, jotta voidaan rakentaa liharakenteita, joilla on määritellyt rasva- ja kollageenisuhteet.
Miksi geenipiirit käyttäytyvät usein eri tavalla 3D-kulttuurissa?
3D-kulttuurissa, geenipiirit käyttäytyvät usein eri tavalla, koska solut käsittelevät fyysisiä ja rakenteellisia syötteitä, joita ei yksinkertaisesti ole olemassa 2D-monokerroksissa. Näihin syötteisiin kuuluvat mekaaninen jännitys, leikkausjännitys, matriisin jäykkyys ja paikallinen solutiheys.
Nämä vihjeet voivat muuttaa signaalireittejä, kuten Notch. Ne voivat myös muuttaa, miten synteettiset piirit havaitsevat voiman ja koordinoivat alavirran vastauksia, mukaan lukien solujen välinen tarttuminen ja kudoksen morfogeneesi.









