

CRISPR muuttaa viljellyn lihan tuotantoa ratkaisemalla merkittävän haasteen: solustressin teollisissa bioreaktoreissa. Tämä työkalu mahdollistaa tarkat geneettiset muokkaukset parantaakseen solujen selviytymistä, pidentääkseen lisääntymistä ja vähentääkseen vanhenemista ankarissa olosuhteissa. Esimerkiksi geenien, kuten TP53 ja PTEN, poistaminen on pidentänyt primaaristen vs. kuolemattomien solulinjojen viljelyaikoja 100:sta 200 päivään ja lisännyt solujen runsautta 1 000-kertaiseksi 30 päivässä. Kuitenkin nämä muutokset voivat vaikuttaa erilaistumiseen, mikä vaatii huolellista optimointia.
Artikkelin keskeisiä havaintoja ovat:
- Stressitekijät bioreaktoreissa: Leikkausvoimat, ravinne-epätasapainot ja oksidatiivinen stressi vähentävät solujen elinkelpoisuutta.
- CRISPR-strategiat: Geenien poisto (TP53, PTEN) ja aktivoinnit (HIF1A) kohdistuvat tiettyihin stressivasteisiin.
- Validointi: Muokatut solut käyvät läpi genomisia, proteomisia ja toiminnallisia testejä suorituskyvyn ja erilaistumispotentiaalin varmistamiseksi.
-
Laajentaminen: Siirtyminen bioreaktorin olosuhteisiin edellyttää optimoituja väliaineita ja laitteita, ja alustat kuten
Cellbase tarjoavat räätälöityjä resursseja.
CRISPR:n tarkkuus mahdollistaa stressinkestävien solulinjojen kehittämisen, mutta kasvun ja erilaistumisen tasapainottaminen on kriittistä skaalautuvan viljellyn lihan tuotannossa.
Bioreaktorin stressiprofiilien kartoittaminen geneettistä suunnittelua varten
Keskeisten bioreaktorin stressitekijöiden tunnistaminen
Ennen CRISPR-muokkauksen aloittamista on tärkeää kartoittaa bioreaktorin stressiprofiilit geneettisen suunnittelun ohjaamiseksi. Bioreaktoreiden stressitekijät aiheuttavat spesifisiä soluvastauksia, jotka on ymmärrettävä hyvin sopivien geneettisten kohteiden valitsemiseksi.
Mekaaninen ja hydrodynaaminen stressi on yksi välittömimmistä haasteista. Sekoitetutankkibioreaktorit luovat leikkausvoimia, jotka voivat vahingoittaa solukalvoja ja häiritä solujen signalointireittejä [5][2]. Ravinto- ja aineenvaihdunnalliset stressit ovat myös merkittävässä roolissa, usein johtuen epätasaisesta ravinteiden otosta. Ravinteiden gradientit 3D-rakenteissa ja ammoniakin kertyminen lisäävät aineenvaihdunnallista rasitusta [3][5][6]. Lisäksi pH:n vaihtelut ja kohonneet lämpötilat voivat vähentää solujen lisääntymisnopeutta ja jopa ohjata soluja ennenaikaiseen erilaistumiseen [3][2].
Muut stressitekijät, kuten oksidatiivinen, mitokondriaalinen ja ER-stressi, haastavat edelleen solujen elinkelpoisuutta. Oksidatiivinen stressi muuttuu erityisen vakavaksi siirryttäessä seerumittomaan viljelyalustaan, sillä luonnollisten antioksidanttien puuttuminen tekee soluista alttiimpia reaktiivisille happilajeille [4]. Solu tasolla, mitokondriaalinen stressi ja endoplasmakalvoston (ER) stressi syntyvät, kun bioprosessin olosuhteet poikkeavat optimaalisista rajoistaan [6]. Xiaoyan Guo Neurodegeneratiivisten sairauksien instituutista UCSF:tä korostaa tätä dynamiikkaa:
"Erilaisten fysiologisten ja ympäristöllisten stressitekijöiden läsnä ollessa solut käynnistävät nopeasti stressivasteet solujen homeostaasin palauttamiseksi." [6]
Kartoittamalla nämä stressitekijät ennakoivasti, sen sijaan että reagoitaisiin ongelmiin niiden ilmetessä, tutkijat voivat määritellä tarkat geenitekniikan tavoitteet.Tämä systemaattinen lähestymistapa varmistaa, että CRISPR-strategiat kohdistuvat tehokkaasti stressinkestävien solulinjojen kehittämiseen.
Omics-datan käyttö stressiin reagoivien geenien löytämiseksi
Stressiympäristön karakterisoinnin jälkeen seuraava vaihe on tunnistaa geenit, jotka reagoivat näihin olosuhteisiin. Työkalut kuten transkriptomiikka (RNA-seq) ja proteomiikka ovat korvaamattomia geeniekspression ja proteiinimäärien muutosten seuraamisessa, kun solut siirtyvät terveistä, varhaisvaiheen tiloista stressaantuneisiin, myöhäisvaiheen olosuhteisiin [1][6]. Kuitenkin, vaikka nämä menetelmät tallentavat alavirran vaikutuksia, ne usein epäonnistuvat tunnistamaan ylävirran säätelijöitä, jotka ajavat näitä muutoksia [6].
Yhdistetyt CRISPR knockout -seulonnat täyttävät tämän aukon.Järjestelmällisesti häiritsemällä tuhansia geenejä suuressa solupopulaatiossa, nämä seulonnat paljastavat, mitkä geenimuutokset antavat kasvuedun stressin alla, paljastaen kriittisiä säätelykeskuksia [1][6]. Esimerkiksi kohdistamalla geeneihin kuten TP53 ja PTEN on osoitettu kääntävän molekulaarisia ikääntymismerkkejä, jotka johtuvat pitkäaikaisesta viljelystressistä. Tämä mahdollistaa myöhäisvaiheen solujen ylläpitää transkriptioprofiilia, joka on samanlainen kuin varhaisvaiheen villityyppisoluilla [1].
Käyttämällä hierarkkista klusterointia, tutkijat voivat ryhmitellä geenejä niiden ilmentymismuutosten perusteella ajan myötä, eristäen moduuleja, jotka liittyvät prosesseihin kuten solusyklin eteneminen ja proteiinisynteesi. Nämä prosessit tyypillisesti heikkenevät, kun bioreaktorin aiheuttama vanheneminen ottaa vallan [1]. Kun yhdistetään reittien rikastusanalyysiin (työkaluilla kuten gprofiler2 ), nämä moduulit voidaan liittää tiettyihin biologisiin reitteihin, kuten TGFβ-signaalointiin tai kondrogeeniseen erilaistumiseen, jotka voivat aktiivisesti rajoittaa solujen laajentumista [1].
Alla oleva taulukko esittelee kunkin menetelmän panoksen kattavan stressikartan rakentamiseen:
| Menetelmä | Pääasiallinen käyttö | Keskeinen tulos |
|---|---|---|
| Transkriptomiikka (RNA-seq) | mRNA-ilmentymismuutosten mittaaminen | Differentially expressed genes (DEGs) stressaantuneiden ja stressaamattomien solujen välillä [1] |
| Proteomiikka | Proteiinien runsauden mittaaminen | Käännöstulokset, jotka on kartoitettu tiettyihin stressitekijöihin [6] |
| Pooled CRISPR -seulonta | Toiminnallinen geenien häirintä | Ylävirran säätelykeskukset ja elintärkeät geenit [1][6] |
| PCA & Hierarkkinen klusterointi | Datan visualisointi ja ryhmittely | Solujen tilan muutokset ja yhteissäädellyt stressivastepolut [1] |
sbb-itb-ffee270
Solulinjojen muokkaus CRISPR-Cas9 - Vinkkejä ja niksejä menestyksen maksimoimiseksi
CRISPR-strategiat stressinkestävien solulinjojen muokkaamiseen
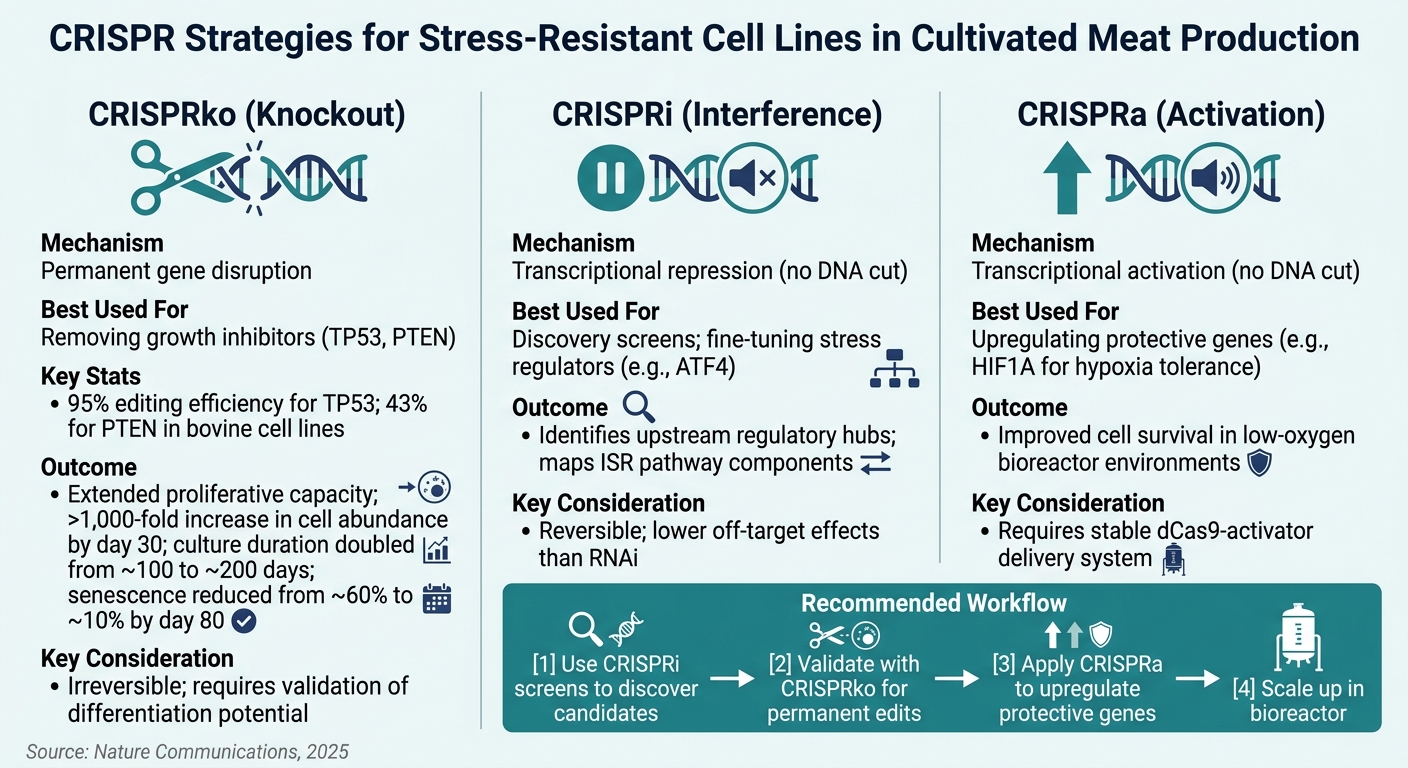
CRISPR-tekniikat stressinkestävien solulinjojen kehittämiseen viljellyssä lihassa
Avaingeenit ja -reitit stressinkestävyyteen
Yksityiskohtaisen stressikartan avulla seuraava askel on kohdegeenien tunnistaminen muokkausta varten.Kohteiden valinta riippuu ensisijaisesta stressitekijästä, joka vaikuttaa solujen suorituskykyyn.
Replikaattinen vanheneminen on merkittävä este viljellyn lihan tuotannossa, koska se rajoittaa solujen lisääntymistä. Noin 25% solulähteistä tällä alalla ovat mesenkymaalisia kantasoluja (MSCs), jotka kohtaavat peruuttamattoman kasvun pysähtymisen toistuvan passauksen jälkeen [1] . TP53 geenin, joka koodaa p53-kasvaimen suppressoriproteiinia, poisto puuttuu suoraan tähän ongelmaan. Tutkimukset naudan MSC:issä osoittavat, että TP53 poisto laajentaa merkittävästi solujen lisääntymiskykyä, jolloin ne voivat jakautua paljon pidemmälle kuin muokkaamattomat linjat [1]. Samoin PTEN poisto parantaa PI3K/AKT/mTOR reittiä, lisäten stressinsietokykyä [1].
Metabolisten ja mitokondriaalisten stressien käsittelemiseksi integroitu stressivaste (ISR) on kriittinen reitti. Transkriptiotekijä ATF4 näyttelee keskeistä roolia mitokondriaalisten stressivasteiden koordinoinnissa, ja CRISPR-seulonnat ovat olleet keskeisiä sen ylävirran säätelijöiden kartoittamisessa [6] . Kuten Xiaoyan Guo ja Martin Kampmann Kalifornian yliopistosta, San Franciscosta, selittävät:
"Puolueettomat geneettiset seulonnat, jotka perustuvat transkriptionaaliseen tai translaationaaliseen raporttiin, ovat tehokkaita lähestymistapoja tunnistaa tietyn stressivasteen säätelytekijöitä." [6]
Myös TGFβ-reitti ansaitsee huomiota, erityisesti naudan MSC-solujen laajentamisessa. CRISPR-seulonnat ovat osoittaneet, että TGFβ-vetoinen kondrogeeninen erilaistuminen tukahduttaa solujen lisääntymistä.Tämän reitin tukahduttaminen auttaa pitämään solut erilaistumattomassa, laajennettavassa tilassa [1]. Hypoksisissa olosuhteissa, joita usein esiintyy tiheissä 3D-tukirakenteiden, ytimissä, HIF1A:n aktivoiminen CRISPRa:lla parantaa solujen selviytymistä vähähappisissa ympäristöissä. Nämä muutokset varustavat solut selviytymään teollisen mittakaavan bioreaktoreiden.
dynaamisissa olosuhteissa.Kuitenkin on tärkeää huomata, että muokkaukset, jotka maksimoivat solujen lisääntymisen - kuten TP53-knockoutit - voivat vähentää solujen kykyä erilaistua lihas- tai rasvakudokseksi. Tämä kasvu- ja erilaistumispotentiaalin välinen kompromissi on tasapainotettava huolellisesti, kun suunnitellaan insinööristrategiaa [1].
| Stressitekijä | Keskeinen geenikohde | CRISPR-strategia | Tulos |
|---|---|---|---|
| Replikatiivinen vanheneminen | TP53 | Knockout | Laajennettu proliferaatiokapasiteetti; lisääntynyt solumäärä |
| Ravinto/kasvustressi | PTEN | Knockout | Tehostunut PI3K/AKT/mTOR-signalointi; parantunut selviytyminen |
| Mitokondriaalinen stressi | ATF4 | CRISPRi / reportteri | Ylävirran säätelyreittien tunnistaminen |
| Hypoksia | HIF1A | CRISPRa (aktivointi) | Lisääntynyt selviytyminen vähähappisissa bioreaktoriympäristöissä |
| Kondrogeneettinen ajautuminen | TGFβ-reitti | Knockout / repressio | Erilaistumattoman, proliferoivan tilan ylläpito naudan MSC-soluissa |
Kun avaingeenit on tunnistettu, oikean CRISPR-tekniikan valinta on seuraava kriittinen vaihe.
CRISPR-muokkaustekniikoiden vertailu
CRISPR-menetelmän valinta määrittää geneettisten muutosten tarkkuuden ja pysyvyyden. Jokaisella lähestymistavalla on omat vahvuutensa riippuen siitä, onko tavoitteena pysyvä muutos, palautuva säätö vai tutkimuksellinen seulonta.
CRISPR knockout (CRISPRko) on ensisijainen menetelmä geenien pysyvään poistamiseen käytöstä. Se on ihanteellinen kohteille kuten TP53 ja PTEN, joissa tarvitaan täydellinen toiminnan menetys. Validointitutkimukset ovat osoittaneet, että CRISPRko saavuttaa 95% muokkaustehokkuuden TP53 ja 43% PTEN naudan solulinjoissa [1] . Nämä vaihtelut korostavat kohdekohtaisen tehokkuuden testaamisen tärkeyttä ennen laajamittaista muokkausta.
CRISPR-interferenssi (CRISPRi) tarjoaa palautuvan geenin tukahduttamisen, mikä tekee siitä ihanteellisen löytövaiheisiin.Se myös vähentää kohteen ulkopuolisia vaikutuksia verrattuna RNAi:hin [6]. Toisaalta, CRISPR-aktivaatio (CRISPRa) toimii yliekspressoimalla suojaavia geenejä, kuten hypoksiansietoon ( HIF1A) tai antioksidanttisuojaukseen liittyviä geenejä, parantaakseen stressinsietokykyä.
Tässä on nopea vertailu tekniikoista:
| Tekniikka | Mekanismi | Paras käyttö | Keskeinen huomioitava asia |
|---|---|---|---|
| CRISPRko | Pysyvä geenin häirintä | Kasvun estäjien poistaminen (TP53, PTEN) | Peruuttamaton; vaatii erilaistumispotentiaalin validoinnin |
| CRISPRi | Transkriptionaalinen repressio (ei DNA-leikkausta) | Löytöseulonnat; säätelijöiden hienosäätö | Palautuva; vähemmän kohteen ulkopuolisia vaikutuksia kuin RNAi:tä |
| CRISPRa | Transkriptionaalinen aktivaatio (ei DNA-leikkausta) | Suojaavien geenien yliregulaatio (HIF1A) | Tarvitsee vakaan dCas9-aktivaattorin toimitusjärjestelmän |
Joukkueille, jotka ovat alkuvaiheessa kohteiden tunnistamisessa, yhdistetyt CRISPRi-seulonnat tarjoavat kustannustehokkaan tavan löytää stressinkestävyyteen liittyviä geenejä laajassa mittakaavassa.Kun lupaavat ehdokkaat on validoitu, CRISPRko:ta voidaan käyttää pysyviin muokkauksiin, jotka soveltuvat tuotantoon. Nämä lähestymistavat täydentävät toisiaan, ja niiden käyttö peräkkäin nähdään yhä enemmän alan parhaana käytäntönä [1][6].
CRISPR-reagenssien ja bioreaktoritarvikkeiden hankintaan, jotka on räätälöity viljellyn lihan tutkimukseen, alustat kuten
CRISPR-muokattujen solulinjojen toteuttaminen ja validointi
CRISPR-muokkausten suunnittelu ja toimitus
Kun olet tunnistanut kohdegeenit, seuraava askel on CRISPR-muokkausten suunnittelu ja toimitus. Tehokkaan geenin häirinnän varmistamiseksi keskity yksittäisten opas-RNA:iden (sgRNA) luomiseen, jotka kohdistuvat olennaisiin eksoniin. Tämä lähestymistapa lisää todennäköisyyttä geenin täydelliselle poistamiselle sen sijaan, että tuotettaisiin katkaistu, osittain toimiva proteiini.Kaksinkertaisen opas-RNA-strategian käyttö voi merkittävästi parantaa knockout-tehokkuutta, nostaen sen noin 55% yli 95% [8].
Valitsemasi toimitusmenetelmä riippuu solutyypistä. Viljellyille lihasolulinjoille valmiiksi kootut Cas9-ribonukleoproteiinit (RNP:t) ovat usein paras vaihtoehto. Nämä RNP:t ovat ohimeneviä, mikä tarkoittaa, että ne hajoavat nopeasti toimituksen jälkeen, mikä auttaa minimoimaan kohteen ulkopuoliset vaikutukset ja välttämään plasmidi-DNA:n integraation riskin [8] . Tapauksissa, joissa käytetään yhdistettyjä seulontoja tai vaikeasti transfektoitavia primaarisia solulinjoja, lentivirusvälitys on luotettava vaihtoehto. Kun käytetään lentivirusjärjestelmiä, tutkijat pitävät yleensä infektiokertoimen (MOI) alhaisena, noin 0,3, välttääkseen useita integraatioita, jotka voisivat monimutkaistaa jatkoanalyysiä [1].
Parhaiden tulosten saavuttamiseksi varmista, että solut ovat logaritmisessa kasvuvaiheessa ja 70–90% konfluenssissa ennen transfektiota. Toimituksen jälkeen eristä yksittäiset kloonit käyttämällä menetelmiä, kuten rajoittava laimennus tai fluoresenssiaktivoitu solulajittelu (FACS), varmistaaksesi selkeän ja yksiselitteisen validoinnin. Lopuksi muokkaukset on vahvistettava genomisella, proteomisella ja toiminnallisella tasolla onnistumisen varmistamiseksi.
Muokattujen solulinjojen seulonta ja validointi
Perusteellinen validointi on välttämätöntä, kun muokattuja solulinjoja siirretään bioreaktorin olosuhteisiin. Tämä prosessi sisältää seulonnan kolmella tasolla: genomisella, proteomisella ja toiminnallisella. Minkä tahansa näistä vaiheista ohittaminen lisää riskiä valita solulinjoja, jotka voivat epäonnistua tuotanto-olosuhteissa.
Genomisella tasolla alkuperäinen seulonta voidaan suorittaa käyttämällä epäsuhtatestejä, kuten T7E1 tai Surveyor, jotka antavat nopean arvion muokkaustiheydestä solupoolissa.Tarkkaa vahvistusta varten seuraa Sanger-sekvensointi tai seuraavan sukupolven sekvensointi (NGS) tunnistaaksesi kloonit, joilla on bialleelisia häiritseviä indelsejä [7][8]. Proteomiikan validointi, joka suoritetaan tyypillisesti Western blot -analyysillä, varmistaa kohdeproteiinin täydellisen puuttumisen. Esimerkiksi vuonna 2025 tehdyssä tutkimuksessa osoitettiin, että TP53:n poisto johti yli 1 000-kertaiseen solumäärän kasvuun 30. päivään mennessä kilpailullisessa seulonnassa, mikä käytännössä kaksinkertaisti viljelykeston 100 päivästä noin 200 päivään [1].
Toiminnallinen validointi on yhtä tärkeää. Metabolista elinkelpoisuutta ja lisääntymisnopeuksia voidaan arvioida Alamar Blue -testeillä, samalla kun seurataan populaation kaksinkertaistumisaikaa (PDT) pitkiä ajanjaksoja - jopa 200 päivää - auttaen tunnistamaan solulinjat, jotka ovat voittaneet replikaation vanhenemisen [1]. Solulinjat, jotka on suunniteltu kestämään hypoksista tai mitokondriaalista stressiä, voidaan vahvistaa FACS-pohjaisilla raportointikokeilla, että solut reagoivat oikein matalan hapen tai ravinteiden rajoittamissa olosuhteissa [6]. Lisäksi TP53- tai PTEN-poistetut solulinjat tulisi testata niiden kyvystä säilyttää erilaistumispotentiaali. Virtaussytometria mesenkymaalisten kantasolujen (MSC) markkereille, kuten CD29 ja CD44, voi vahvistaa, että nämä solut säilyttävät kantasolumaisuutensa [1].
| Validointitaso | Menetelmä | Tarkoitus |
|---|---|---|
| Genomiikka | Sanger-sekvensointi / NGS | Vahvista bialleeliset häiritsevät indelit [7][8] |
| Proteomiikka | Western Blot | Varmista kohdeproteiinin täydellinen puuttuminen [7][8] |
| Fenotyyppinen | Virtsanäytevirtaus (CD29/CD44) | Tarkista MSC-merkkien ja kantasolukyvyn säilyminen [1] |
| Toiminnallinen | Alamar Blue / PDT-seuranta | Arvioi kasvukinetiikka ja metabolinen terveys [1] |
| Stressi | FACS-pohjaiset raportointianalyysit | Tutki stressivasteen käyttäytymistä haastavissa olosuhteissa [6] |
Ennen muokatun solulinjan laajentamista, suorita STR-profilointi solujen identiteetin varmistamiseksi ja tee mykoplasmatesti kontaminaation poissulkemiseksi [7]. Kolmen kuukauden ajan kestää yleensä luoda validoitu knockout-solulinja, ja tietyt työnkulun vaiheet saattavat vaatia toistoa.
Laajentaminen: Stressinkestävien solulinjojen siirtäminen tuotantoon
Muokattujen solulinjojen siirtäminen bioreaktorin olosuhteisiin
Kun solulinjat on validoitu, niiden on siirryttävä laboratoriomittakaavan adherenttikulttuureista suspensiojärjestelmiin, kuten sekoitussäiliöbioreaktoreihin, ilmankohotusreaktoreihin tai pyöriväseinäisiin astioihin - jokainen näistä pystyy tukemaan teollisen mittakaavan viljellyn lihan tuotantoa [2].
Adherenttiriippuvaisille soluille, kuten naudan mesenkymaalisille kantasoluille (bMSC), laminin-511-pinnoitettujen mikrokantajien käyttö tarjoaa käytännöllisen polun suspensiokulttuuriin [3]. Tämän siirtymän aikana on tärkeää seurata MSC-markkereita, kuten CD29 ja CD44, jotta varmistetaan solujen erilaistumispotentiaalin säilyminen [1].
Kriittinen askel skaalaamisessa on median uudelleenmuotoilu. Seerumipohjainen media tulisi korvata kemiallisesti määritellyillä, seerumittomilla koostumuksilla, jotka on rikastettu lipideillä, ei-välttämättömillä aminohapoilla ja antioksidanteilla solujen elinkelpoisuuden ylläpitämiseksi laajamittaisissa olosuhteissa [4]. Huomattavasti, CRISPR-muokatut solulinjat TP53- ja PTEN-poistogeenien kanssa ovat paremmin varustettuja tähän siirtymään. Tutkimus, joka julkaistiin Nature Communications (2025), osoitti, että nämä muokkaukset pidentivät bMSC:iden proliferaatiokautta noin 100 päivästä yli 200 päivään, samalla kun ne vähensivät vanhenemista noin 60% vain 10% päivään 80 mennessä [1].
"TP53- ja PTEN-poistogeenit lisäsivät merkittävästi proliferaatiota ja viivästyttivät vanhenemista." - Nature Communications [1]
Siirtymävaiheen aikana työkalut, kuten Alamar Blue -testit ja qRT-PCR, ovat olennaisia solujen elinkelpoisuuden seuraamisessa ja geneettisten muutosten vakauden varmistamisessa. Nämä CRISPR-muokatut naudansolulinjat ovat osoittaneet keskimääräisen 12% parannuksen kaksinkertaistumisnopeuksissa, ja jotkut ovat saavuttaneet 50% kasvun 50. päivään mennessä [1]. Kun solut osoittavat vakaata suorituskykyä bioreaktorin olosuhteissa, voidaan keskittyä tarvittavien erikoislaitteiden hankintaan mittakaavan kasvattamiseksi.
Laitteiden ja Materiaalien Hankinta Mittakaavan Kasvattamiseksi
Tuotantotason bioreaktorikäyttöön siirtyminen tuo mukanaan merkittäviä haasteita hankinnassa. Solujen sopeutumisen varmistamisen jälkeen tarvittavien materiaalien ja laitteiden hankinta tulee ensisijaiseksi.Tuotteet, kuten kertakäyttöiset sekoitussäiliöbioreaktorit, validoidut mikrokantajat, seerumittomat väliainekomponentit ja FACS-järjestelmät jatkuvaan kloonien seurantaan, ovat erittäin erikoistuneita ja usein yleisten laboratoriotoimittajien ulottumattomissa.
Alustat kuten
Päätelmä
CRISPR-tekniikka on siirtynyt tutkimustyökalusta käytännölliseksi menetelmäksi solulinjojen muokkaamiseen viljellyn lihan tuotannossa. Kohdistamalla keskeisiin säätelijöihin, kuten TP53 ja PTEN, tutkijat ovat merkittävästi pidentäneet solujen lisääntymistä, käytännössä kaksinkertaistaen tyypillisen viljelykeston [1]. Tämä edistysaskel laajentaa viljellyn lihan skaalautuvan tuotannon.
Kuitenkin matka muokatuista solulinjoista täysimittaiseen tuotantoon vaatii perusteellista validointia jokaisessa vaiheessa. On varmistettava, että muokatut solut säilyttävät kykynsä erilaistua lihas- ja rasvakudokseksi, mikä on yhtä kriittistä kuin nopean lisääntymisen saavuttaminen. Ilman tätä, edes nopeimmin kasvavat solulinjat eivät olisi kaupallisesti elinkelpoisia [1]. Tämä korostaa tarvetta tiukoille validointiprosesseille, jotta voidaan varmistaa, että parantunut lisääntyminen johtaa merkityksellisiin tuotantotuloksiin.
Nature Communications vahvistaa tämän lähestymistavan toteamalla:
"Nämä havainnot osoittavat CRISPR-seulonnan hyödyllisyyden naudan kantasolujen ominaisuuksien optimoimisessa ja tarjoavat polun kohti laajamittaisempaa viljellyn lihan tuotantoa tulevaisuudessa." [1]
Huolimatta näistä edistysaskeleista, käytännön haasteet, kuten hankinta, voivat hidastaa edistymistä. Riippuvuus yleistoimittajista sgRNA-kirjastoille, kertakäyttöisille bioreaktoreille ja seerumittomille kasvatusalustoille aiheuttaa usein yhteensopivuusongelmia ja viivästyksiä. Alustat kuten
Sopivien materiaalien saatavuus on yhtä tärkeää kuin itse geenitekniikka. Kuten Nature Communications on todennut, vaikka viljelty liha tarjoaa lupaavan vaihtoehdon perinteiselle lihalle, skaalautuvuus ja kustannustehokkuus ovat edelleen merkittäviä esteitä. CRISPR-pohjainen suunnittelu, kun se yhdistetään kurinalaiseen bioprosessisuunnitteluun ja tehostettuun hankintaan alustojen, kuten
UKK
Mitkä bioreaktorin stressitekijät tulisi kartoittaa ennen CRISPR-kohteiden valintaa?
Kun valitaan CRISPR-kohteita stressinkestävien solulinjojen kehittämiseksi viljellyn lihan tuotannossa, on tärkeää arvioida ensisijaiset bioreaktorin stressitekijät, jotka vaikuttavat solujen kasvuun ja selviytymiseen. Näitä stressitekijöitä ovat:
- Leikkausjännitys: Bioreaktoreissa olevat solut altistuvat usein mekaanisille voimille sekoituksen ja ilmastuksen vuoksi. Pitkäaikainen leikkausjännitys voi vahingoittaa solukalvoja ja heikentää kasvua.
- Happitasot: Optimaalisten happipitoisuuksien ylläpitäminen on elintärkeää. Liian vähäinen happi voi rajoittaa energiantuotantoa, kun taas liiallinen happi voi johtaa oksidatiiviseen stressiin.
- Ravinteiden saatavuus: Solut tarvitsevat jatkuvaa ravinteiden saantia. Mikä tahansa epätasapaino tai puute voi haitata lisääntymistä ja tuottavuutta.
- pH-vaihtelut: Solut menestyvät kapealla pH-alueella. Poikkeamat voivat häiritä aineenvaihduntaprosesseja ja entsyymiaktiivisuutta.
- Lämpötilan vaihtelut: Jopa pienet muutokset lämpötilassa voivat vaikuttaa solujen toimintoihin, aiheuttaen stressiä tai vähentäen elinkelpoisuutta.
- Jätteen kertyminen: Aineenvaihdunnan sivutuotteet, jos niitä ei poisteta tehokkaasti, voivat muuttua myrkyllisiksi ja estää solujen kasvua.
Ymmärtämällä perusteellisesti nämä stressitekijät, tutkijat voivat tunnistaa kriittiset stressivastepolut. Tämä tieto mahdollistaa kohdennetut geneettiset muutokset CRISPR-tekniikalla, parantaen solulinjojen kestävyyttä ja varmistaen vahvemman suorituskyvyn bioreaktoriolosuhteissa.
Kuinka tasapainotan nopeammat kasvueditoinnit lihas- ja rasvadifferentiaation kanssa?
Nopean kasvun tasapainottaminen lihas- ja rasvadifferentiaation kanssa viljellyn lihan tuotannossa vaatii huolellista genetiikan ja viljelyolosuhteiden hallintaa. CRISPR-teknologia on keskeisessä roolissa tässä, mahdollistaen geenien, kuten TP53 ja PTEN. tarkennetut muokkaukset. Nämä säädöt voivat edistää solujen lisääntymistä samalla säilyttäen solujen kyvyn erilaistua lihas- ja rasvakudokseksi.
Viljelyolosuhteiden hienosäätö ja geeniekspression säätely ovat yhtä kriittisiä halutun tasapainon saavuttamiseksi. Resurssit, kuten
Mikä on vähimmäisvalidaatio, joka tarvitaan ennen bioreaktorin skaalausta?
Ennen siirtymistä bioreaktoreihin on tärkeää varmistaa, että geneettisesti muokatut solulinjat säilyttävät vakaat ja toivotut ominaisuudet, kuten parantuneet kasvunopeudet, stressinsietokyvyn ja erilaistumiskyvyn. Tämä validointiprosessi tulisi arvioida geneettistä vakautta ja varmistaa johdonmukainen suorituskyky bioprosessiolosuhteissa. Moni-omisten analyysien ja stressivasteprofiilien tukevat tiedot ovat avainasemassa tässä arvioinnissa. Korkean läpimenon CRISPR-seulonnan avulla voidaan tunnistaa geneettisiä muokkauksia, jotka parantavat solujen lisääntymistä ja elinikää, mikä tekee näistä solulinjoista sopivampia skaalautuvaan viljellyn lihan tuotantoon.










