

Viljellyn lihan tuotannon tutkijoille apoptoosin minimointi on olennaista solujen elinkelpoisuuden ja tuottavuuden parantamiseksi bioreaktoreissa. Stressitekijät, kuten ravinteiden ehtyminen, osmoottiset epätasapainot ja jätteen kertyminen, aiheuttavat usein solukuolemaa, mikä vähentää saantoja. Anti-apoptoottiset geenit voivat lieventää näitä haasteita pidentämällä solujen elinikää viljelyn aikana. Tässä on nopea yleiskatsaus tärkeimmistä geeneistä ja niiden rooleista:
- BCL-2: Estää mitokondrioiden huokosten muodostumisen, mikä estää apoptoosin alkamisen. Tehokas erilaistumattomille soluille, mutta vaatii huolellista tasapainoa pro-apoptoottisten proteiinien kanssa.
- BCL-xL: Suojaa soluja erilaistumisen aikana ja tukee energian aineenvaihduntaa. Ihanteellinen korkean stressin vaiheisiin bioreaktoreissa.
- MCL-1: Tarjoaa nopean vasteen ravinteiden muutoksiin ja pysyy vakaana erilaistumisen aikana. Toimii hyvin yhdessä muiden geenien kanssa.
- BIRC5 (Survivin) : Estää kaspaaseja estääkseen apoptoosin alavirtaan. Tukee lisääntymistä nopeasti jakautuvissa soluissa.
- XIAP: Tehokas kaspaasi-inhibiittori, joka on tehokas äärimmäisissä stressiolosuhteissa, kuten tiheissä viljelmissä. Näiden olosuhteiden seuranta vaatii antureiden valitsemista viljellyn lihan bioreaktoreille ravintoaineiden tasojen ja jätteen kertymisen seuraamiseksi reaaliajassa.
Nopea vertailu
| Geeni | Avaintarkoitus | Stabiilisuus erilaistumisen aikana | Paras käyttötapaus |
|---|---|---|---|
| BCL-2 | Estää varhaisen apoptoosin (BAX/BAK) | Stabiili | Erilaistumattomien solujen säilyttäminen |
| BCL-xL | Estää kaspaasien aktivoitumisen, tukee aineenvaihduntaa | Vaihekohtainen | Erilaistuvat solut stressin alla |
| MCL-1 | Nopea reagointi ravintomuutoksiin | Stabiili | Monivaiheinen selviytyminen |
| BIRC5 | Estää kaspaaseja alavirtaan | Vähenee erilaistumisen myötä | Nopeasti jakautuvat solut |
| XIAP | Laaja kaspaasi-inhibitio | Vakaa | Korkean stressin bioreaktorin olosuhteet |
1.BCL-2
BCL-2 on hyvin tutkittu anti-apoptoottinen geeni, joka näyttelee keskeistä roolia solunsisäisessä (mitokondriaalisessa) apoptoosipolussa. Tämä polku on merkittävä solukuoleman mekanismi, joka usein käynnistyy viljellyissä lihassoluissa bioreaktorin stressitekijöiden, kuten ravinnepulan tai alhaisen happitason, vaikutuksesta.
BCL-2 toimii sitoutumalla ja neutraloimalla pro-apoptoottisia proteiineja, kuten BAX ja BAK. Tämä toiminta estää mitokondriaalisten huokosten muodostumisen, pysäyttäen sytokromi c:n vapautumisen ja apoptoosin alavirran kaskadin. Tämä mekanismi on ratkaisevan tärkeä solujen elinkelpoisen eliniän pidentämiseksi viljellyn lihan tuotannossa. Kuten Rønning SB et al. selittävät:
"Bcl-2:n ja Bax:n välinen suhde määrää solujen herkkyyden apoptoosille."[5]
Mitokondriaalisen roolinsa lisäksi BCL-2 sijaitsee myös endoplasmakalvostossa (ER).Tässä se vähentää kalsiumtasoja ja estää IP3-reseptorin välittämän kalsiumin vapautumisen, lieventäen kalsiumin aiheuttamaa apoptoosia – yleinen ongelma tiheissä bioreaktoriviljelmissä[4]. Näiden skaalaushaasteiden hallinta on teollisuuden ensisijainen tavoite. Tämä kaksoispaikannus mahdollistaa BCL-2:n suojata soluja useilta apoptoosin laukaisijoilta.
BCL-2:n molekyylirakenne, joka koostuu kahdeksasta alfa-heliksin nipusta ja neljästä hyvin määritellystä BH-domeenista, tekee siitä e
Kuitenkin on olemassa kriittinen varoitus: BCL-2:n ja pro-apoptoottisten proteiinien, kuten BAX:n, välistä tasapainoa on hallittava huolellisesti. Jopa korkeat BCL-2:n ilmentymistasot eivät välttämättä estä apoptoosia, jos pro-apoptoottiset signaalit vahvistuvat liikaa[2]. Tämän tasapainon seuranta on olennaista optimaalisen solujen elinkelpoisuuden saavuttamiseksi.
sbb-itb-ffee270
2. BCL-xL
BCL-xL, koodattu BCL2L1 geenillä, on keskeisessä asemassa BCL-2-perheessä paikantumalla ulkoiseen mitokondrion kalvoon ja estämällä apoptoosia. Se saavuttaa tämän vastustamalla pro-apoptoottisia proteiineja, kuten BAX ja BAK.Lisäksi se estää pilkotun kaspaasi-3:n (CASP3), joka on välttämätön solukuoleman pysäyttämiseksi. Tämä mekanismi on erityisen arvokas tiheissä bioreaktoriviljelmissä , missä metabolinen stressi voi uhata solujen elinkykyä.
Kiinnostavaa on, että BCL-xL:n aktiivisuus liittyy tiettyihin erilaistumisvaiheisiin. Tiettyjen vaiheiden aikana sen ilmentyminen lisääntyy, kun taas muut anti-apoptoottiset proteiinit, kuten BCL-2 ja MCL-1, pysyvät muuttumattomina. Tämä korostaa sen merkitystä solujen selviytymisen ylläpitämisessä erilaistumisen aikana. Kuten on todettu Cell Death & Disease:
"BCL-xL/BCL2L1 on kriittinen anti-apoptoottinen proteiini, joka edistää erilaistuvien... solujen selviytymistä." [2]
Apoptoosin roolin lisäksi BCL-xL tukee solujen energiametaboliaa. Se parantaa sekä glykolyysiä että oksidatiivista fosforylaatiota, varmistaen korkean metabolisen aktiivisuuden.BCL-xL:n estämisen on osoitettu vähentävän metabolisten geenien ilmentymistä ja alentavan sekä perus- että maksimaalista mitokondrioiden hengitystä. Tämä toiminto on erityisen tärkeä viljellyille lihasoluille, jotka ovat riippuvaisia jatkuvasta metabolisesta tuotannosta.
BCL-xL on erittäin yhteensopiva geenieditointistrategioiden kanssa, joita käytetään yleisesti viljellyn lihan tutkimuksessa. Tekniikat, kuten lentivirusvälitteinen siirto, mahdollistavat BCL2L1-geenin vakaan integroinnin, kun taas doksisykliini-indusoitavat CRISPR/Cas9-järjestelmät tarjoavat tarkan ajallisen kontrollin sen ilmentymiselle [2] [6]. Tämä tarkkuuden taso hallitaan usein kehittyneen bioprosessinohjausohjelmiston. kautta. Nämä ominaisuudet tekevät BCL-xL:stä vahvan ehdokkaan parantamaan solulinjojen elinkelpoisuutta viljellyn lihan tuotannossa.
Differentiatiivaiheissa, joissa on korkeat metaboliset vaatimukset, BCL-xL voi olla tehokkaampi kuin BCL-2.Tutkijat voivat käyttää estäjää WEHI-539 testatakseen solulinjan riippuvuutta BCL-xL:stä ennen pysyvien geneettisten muutosten tekemistä [2]. Lisäksi BCL-xL:n ja MCL-1:n yhteisilmaisu voisi parantaa solujen selviytymistä, sillä näiden proteiinien on havaittu toimivan synergisesti joissakin resistenttityyppisissä soluissa [6].
3. MCL-1
MCL-1 (Myeloid Cell Leukaemia-1) on keskeisessä asemassa säätelemässä sisäistä apoptoottista reittiä. Se sijaitsee ulommalla mitokondrion kalvolla ja estää apoptoosin sitomalla ja eristämällä pro-apoptoottiset proteiinit BAX ja BAK, estäen niiden oligomerisaation ja sitä seuraavan kalvon läpäisevyyden. Tämä toiminta estää sytokromi c:n vapautumisen, pysäyttäen apoptoottisen kaskadin ennen kuin se saavuttaa toimeenpanovaiheen [8] . Lisäksi MCL-1 sitoutuu BH3-only -proteiineihin - kuten Bim, PUMA ja NOXA - korkealla affiniteetilla [8]. Kuten BCL-2 ja BCL-xL, MCL-1 on elintärkeä apoptoottisten signaalien vastustamisessa, erityisesti bioreaktoristressin aikana.
Yksi MCL-1:n ainutlaatuisista ominaisuuksista on sen lyhyt puoliintumisaika, mikä tekee sen ilmentymisestä erittäin herkän ravinteiden saatavuudelle ja metabolisille signaaleille, erityisesti AMPK/mTOR-reitin kautta. Tutkimukset osoittavat, että 25% kalorinsaannin vähentäminen voi vähentää MCL-1:n translaatiota noin 39% ± 10% [7] . Tämä herkkyys on erityisen merkityksellistä viljellyn lihan tuotannossa, jossa kasvatusalustan koostumuksen vaihtelut tai ravinteiden ehtyminen suurimittaisissa suspensioviljelmissä (jotka vaativat huolellista tuotannon mittakaavan suunnittelua) voivat merkittävästi alentaa MCL-1-tasoja.Tällaiset vähennykset heikentävät solujen elinkelpoisuutta, mikä heikentää anti-apoptoottisten strategioiden avulla saavutettuja parannuksia IVCC:ssä (integroitu elinkelpoisten solujen pitoisuus). Tämän lieventämiseksi seerumittomat väliaineformuloinnit, jotka tukevat vahvaa mTORC1-aktiivisuutta, ovat välttämättömiä [7] .
Toinen huomionarvoinen piirre MCL-1:ssä on sen vakaus erilaistumisen aikana. Haiman progenitorimallissa MCL-1:n ilmentyminen pysyi vakaana koko 17 päivän erilaistumisprotokollan ajan, toisin kuin BCL-xL, joka osoitti vaihekohtaista vaihtelua [2]. Tämä vakaus tekee MCL-1:stä erityisen edullisen viljellyn lihan sovelluksissa, joissa solujen on selviydyttävä useista kypsymisvaiheista ilman tarkasti ajoitettuja interventioita.
Geenieditointityökaluja voidaan käyttää MCL-1:n muokkaamiseen, aivan kuten muita anti-apoptoottisia geenejä, mikä tekee siitä monipuolisen kohteen solulinjojen suunnittelussa.
Kun MCL-1:ä käytetään yhdessä muiden anti-apoptoottisten geenien kanssa, se tarjoaa lisäetuja. Esimerkiksi MCL-1:n yhdistäminen BCL-xL:n kanssa on osoittanut synergistisiä vaikutuksia - molempien proteiinien samanaikainen estäminen vähensi selviytymislääkkeiden EC50-arvoa noin 10 μM:sta alle 20 nM:iin [6]. Tämä lähestymistapa voi merkittävästi parantaa solujen selviytymistä viljellyn lihan tuotannon korkean stressin vaiheissa.
4. BIRC5 (Survivin)
BIRC5, jota usein kutsutaan Surviviniksi, on osa Apoptoosin Estäjä (IAP) proteiiniperhettä [2]. Toisin kuin BCL-2 perheen proteiinit, jotka toimivat mitokondrion kalvolla estääkseen apoptoosin alkamisen, BIRC5 toimii pidemmällä alavirtaan. Se estää apoptoosin suorittamisesta vastaavat kaspaasit, toimien tehokkaasti viimeisenä puolustuslinjana ohjelmoitua solukuolemaa vastaan [10].
Suspensioviljelmissä stressitekijät, kuten ravinteiden ehtyminen, aineenvaihduntajätteiden kertyminen ja mekaaninen leikkausjännitys, voivat laukaista apoptoosin. Estämällä kaspaasiaktiivisuutta tässä myöhemmässä vaiheessa BIRC5:n yliekspressio auttaa pidentämään solujen elinkelpoisuutta ja tuottavuutta. Tämä parantaa elinkelpoisten solujen pitoisuuden aikaintegraalia - keskeinen mittari soluviljelyn suorituskyvyn optimoinnissa [9] . Eric Baek, tutkija KAIST:ssa, selittää:
"Elinkelpoisten solujen pitoisuuden aikaintegraalin parantaminen voittamalla solukuolema, nimittäin apoptoosi, on yksi laajimmin käytetyistä strategioista terapeuttisten proteiinien [ja solujen] tehokkaaseen tuotantoon." [9]
Tämä alavirran interventio on osoittautunut parantavan bioreaktorin tuottoa viljellyissä lihan solulinjoissa, mukaan lukien sian satelliittisolut ja naudan myoblastit.
Tehokkain strategia sisältää kombinatorisen suunnittelun, yhdistämällä BIRC5 mitokondrioiden suojaajien, kuten BCL-2 tai BCL-xL, kanssa. Professori Michael Betenbaugh Johns Hopkinsin yliopistosta korostaa tätä lähestymistapaa:
"Strategiat, jotka estävät solukuoleman useissa kohdissa kaskadia pitkin, voivat rajoittaa näiden apoptoosin signaalien voimistumista." [10]
Yhdistämällä BIRC5:n kaspaasi-inhibition ylävirran mitokondriosuojaukseen, tutkijat voivat luoda monikerroksisen puolustuksen apoptoosia vastaan.
BIRC5 integroituu myös saumattomasti geenieditointityönkulkuihin.CRISPR/Cas9 on johtava menetelmä stabiilien solulinjojen luomiseksi yliekspressiolla [9], vaikka sinkkisorminukleaasit tarjoavat tarkan vaihtoehdon. siRNA:ta voidaan käyttää reittien validointiin ennen sitoutumista genomiseen integraatioon [9].
5. XIAP
XIAP (X-linked inhibitor of apoptosis) tunnustetaan tehokkaimmaksi kaspaasi-inhibiittoriksi IAP (inhibitor of apoptosis protein) -perheessä. Yhdessä geenien kuten BCL-2 ja MCL-1 kanssa, XIAP:lla on kriittinen rooli apoptoosin kohdentamisessa sen toteutusvaiheessa. Kuten korostettu Genes & Development :
"XIAP:ta pidetään tehokkaimpana kaspaasi-inhibiittorina in vitro." [12]
XIAP käyttää kahta erillistä mekanismia apoptoosin estämiseen. Ensinnäkin sen BIR2-domeeni ja linkkerialue estävät efektorikaspaasit-3 ja -7.Toiseksi, sen BIR3-domeeni estää kaspaasi-9:n, pysäyttäen tehokkaasti mitokondrioiden sisäisen apoptoottisen reitin. Lisäksi sen C-terminaalinen RING-domeeni helpottaa kohdekaspaasien ubikitiinointia ja sitä seuraavaa proteasomaalista hajoamista [11]. Puuttumalla sekä sisäisiin että ulkoisiin apoptoottisiin reitteihin, XIAP osoittautuu erittäin tehokkaaksi käsitellessään apoptoosin laukaisijoita, kuten ravinnepuutetta, aineenvaihduntatuotteita ja mekaanista stressiä - tekijöitä, joita tavallisesti kohdataan viljellyn lihan tuotantojärjestelmissä. Sen toiminnallisuutta parantaa entisestään sen vahva säilyminen lajien välillä.
Esimerkiksi ihmisen XIAP jakaa 87.7% proteiini-identiteetin Bos taurus ( nauta) ja 89.5% kanssa Mus musculus (hiiri) [11] . Tämä korkea samankaltaisuus mahdollistaa nisäkkäiden mallijärjestelmistä saadun tutkimuksen luotettavan soveltamisen solulinjoihin, joita käytetään viljellyn lihan tuotannossa.
XIAP:ia voidaan säädellä käyttämällä työkaluja, kuten shRNA, antisense-oligonukleotideja tai CRISPR/Cas9 [11]. Äärimmäisen stressin alaisena sen RING-domeeni voi indusoida itseubikytinaation [12], kun taas endogeeniset estäjät, kuten SMAC/DIABLO ja HTRA2, voivat syrjäyttää XIAP:in kaspaaseista [11][13]. Nämä havainnot tekevät XIAP:ista houkuttelevan kohteen geenieditointimenetelmille, joiden tavoitteena on optimoida solulinjoja viljellyn lihan kehittämiseksi.Anti-apoptoottisten geenien vertailu yhdellä silmäyksellä
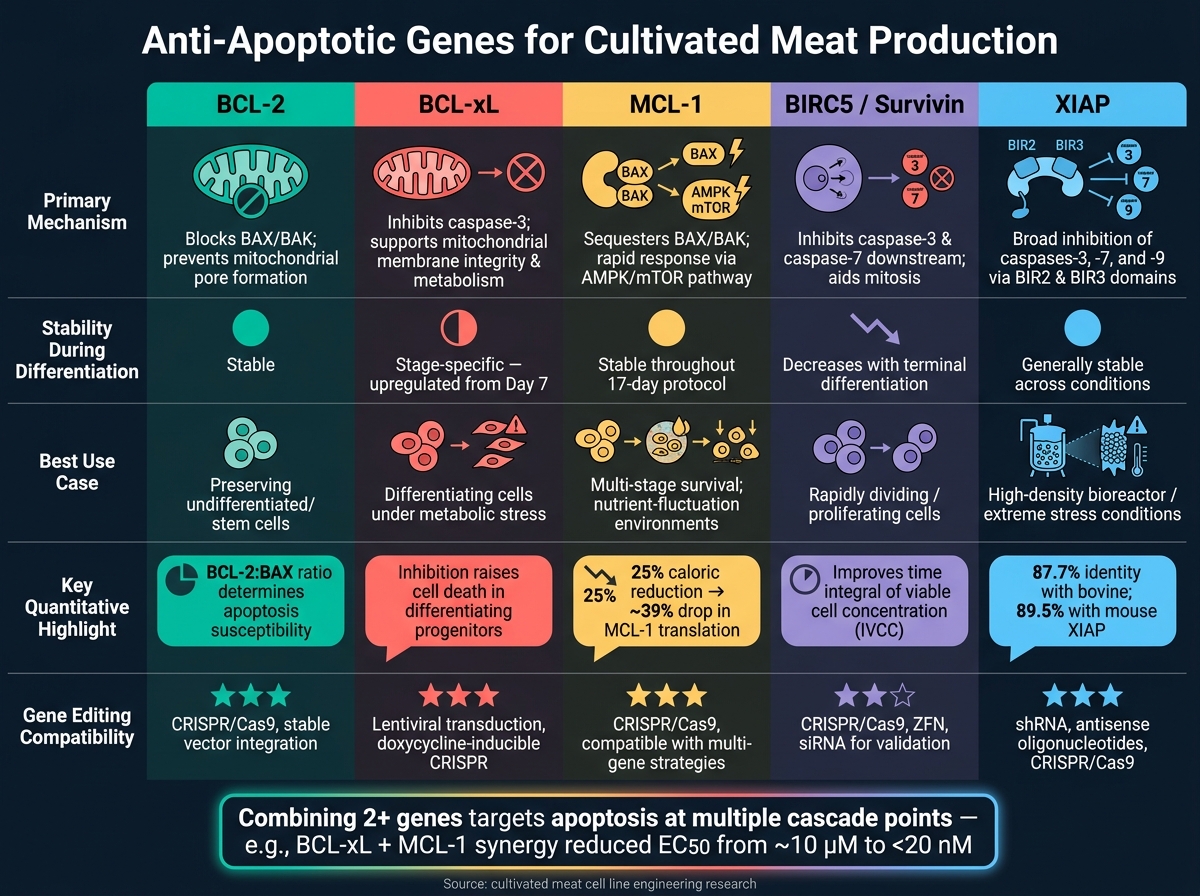
Anti-apoptoottiset geenit viljellylle lihalle: rinnakkainen vertailu
Kun työskentelet viljellyn lihan tuotannon parissa, eri anti-apoptoottisten geenien toiminnan ymmärtäminen voi auttaa hienosäätämään solulinjojen suunnittelua. Jokaisella geenillä on oma erityinen mekanisminsa, käyttäytymisensä erilaistumisen aikana ja mahdolliset sovellukset. Alla oleva taulukko tiivistää nämä erot, mikä helpottaa päätöksentekoa siitä, mikä geeni - tai geenien yhdistelmä - saattaa toimia parhaiten tarpeisiisi.
| Geeni | Ensisijainen mekanismi | Ilmentymän vakaus | Raportoitu elinkelpoisuuden vaikutus | Muokkausyhteensopivuus |
|---|---|---|---|---|
| BCL-2 | Estää pro-apoptoottisen BAX/BAK:n ja varmistaa erilaistumattomien solujen selviytymisen [2] | Pysyy suhteellisen vakaana erilaistumisen aikana [2] | Välttämätön alkuperäisen kantasolupoolin säilyttämiseksi [2] | Korkea yhteensopivuus muokkaustyökalujen kanssa |
| BCL-xL | Estää pilkotun kaspaasi-3:n; ylläpitää mitokondrioiden kalvon eheyttä ja aineenvaihduntaa [2] | Ylösreguloituu erilaistumisen päivänä 7 alkaen [2] | Kriittinen erilaistuvien progenitorien tukemisessa; sen estäminen lisää solukuolemaa [2] | Korkea yhteensopivuus muokkaustyökalujen kanssa |
| MCL-1 | Moduloi pro-apoptoottisia signaaleja osana BCL-2-perhettä [2] | Ilmentyminen pysyy vakaana linjaspesifikaation aikana [2] | Tarjoaa laajoja selviytymisetuja, mutta puuttuu vaihekohtaisia vaikutuksia kuten BCL-xL [2] | Korkea yhteensopivuus muokkaustyökalujen kanssa |
| BIRC5 (Survivin) | Estää kaspaasi-3 ja kaspaasi-7; auttaa kromosomien jakautumisessa mitoosin aikana | Korkea proliferoituvissa soluissa; vähenee terminaalisen erilaistumisen myötä | Tukee selviytymistä ja proliferaatiota nopeasti jakautuvissa soluissa | Yhteensopiva sekä shRNA knockdownin että CRISPR-editoinnin kanssa |
| XIAP | Estää useita kaspaaseja, tarjoten laajaa apoptoottista suojaa | Yleisesti vakaa eri olosuhteissa | Erityisen tehokas stressin alla, kuten suuritiheyksisissä bioreaktoriolosuhteissa | Korkea yhteensopivuus editointityökalujen kanssa |
BCL-xL erottuu edukseen kaksoisroolistaan solujen selviytymisen edistämisessä ja aineenvaihdunnan tukemisessa, erityisesti kriittisen erilaistumisvaiheen aikana, jolloin pro-apoptoottiset proteiinit kuten BAK luonnollisesti vähenevät.BCL-2, toisaalta on ihanteellinen erilaistumattomien solujen säilyttämiseen, kun taas XIAP tarjoaa laajaa suojaa, erityisesti stressaavissa ympäristöissä, kuten tiheissä viljelmissä.
Mikään yksittäinen geeni ei toimi parhaiten kaikissa tilanteissa. Esimerkiksi BIRC5 on erityisen hyödyllinen tilanteissa, joissa tarvitaan nopeaa solujen jakautumista. Käytännössä kahden tai useamman geenin yhdistäminen tarjoaa usein tehokkaimman suojan, käsitellen useita apoptoottisia laukaisimia samanaikaisesti.
Nämä havainnot tarjoavat perustan näiden geenien sisällyttämiselle solulinjojen suunnittelustrategioihin viljellyn lihan tuotantoa varten. Tämä sisältää oikeiden viljellyn lihan syötteiden valitsemisen skaalautuvuuden varmistamiseksi.
Näiden geenien käyttö viljellyn lihan solulinjojen suunnittelussa
Solujen elinkelpoisuuden parantamiseksi viljellyn lihan tuotannossa on keskeisten geenien strateginen integrointi ratkaisevan tärkeää.Ei riitä, että tunnistetaan anti-apoptoottisia geenejä - niiden tehokas sisällyttäminen solulinjoihin on se, mikä tekee eron. Kaksi päästrategiaa ovat yleisesti käytössä: anti-apoptoottisten geenien yliekspressio kuten BCL-2, BCL-xL, ja MCL-1 solujen selviytymisen parantamiseksi tai pro-apoptoottisten geenien poisto kuten BAX, BAK , ja BOK solukuoleman ajureiden poistamiseksi. Näiden lähestymistapojen yhdistäminen johtaa usein solulinjoihin, jotka soveltuvat paremmin laajamittaiseen tuotantoon [1].
Modernit geenieditointityökalut kuten CRISPR/Cas9 mahdollistavat samanaikaiset muokkaukset, kuten Bak1, Bax, ja Bok poiston yhdessä vaiheessa. Vaihtoehtoja kuten ZFNs tai RNA-interferenssi voidaan käyttää tilapäisesti vähentämään kaspaasien aktiivisuutta (e.g. kaspaseja-3, -7, -8 ja -9). Yli-ilmentämisstrategioissa synteettiset promoottorit varmistavat tasaisen ja korkean geenien, kuten BCL-2, ilmentymistason mittakaavan kasvattamisen aikana, mikä on kriittistä solujen suorituskyvyn ylläpitämiseksi syöttöerä- tai jatkuvissa viljelyjärjestelmissä. Nämä yhdistetyt menetelmät vahvistavat solulinjojen kehitystä viljellyn lihan sovelluksissa.
Tällaiset geneettiset muutokset vaikuttavat suoraan parantuneeseen integroidun elinkelpoisen solupitoisuuden (IVCC) , keskeiseen mittariin viljellyn lihan tuotannossa. Solukuolema on voimakkainta erilaistumisen ensimmäisten viiden päivän aikana, mikä tekee varhaisista interventioista geenien, kuten BCL-2 tai BCL-xL, kanssa olennaisia. Tutkimus, joka on julkaistu Cell Death & Disease, korostaa, että BCL-xL-ilmentyminen lisääntyy solujen erilaistuessa, mikä osoittaa, että kypsämmät progenitorit ovat voimakkaasti riippuvaisia sen suojaavasta roolista [2] . Seuraamalla BCL-2-perheen geenien ilmentymistasoja kasvuvaiheiden aikana, interventiot voidaan ajoittaa tarkasti maksimaalisen vaikutuksen saavuttamiseksi.
"Vakiinnuttamalla solulinjoja, jotka yliekspressoivat antiapoptoottisia geenejä tai alasreguloivat proapoptoottisia geenejä, lopputuotteen saantoa voidaan parantaa, kun solut tulevat kestävämmiksi ympäristön stressitekijöitä vastaan." - Gyun Min Lee et al. [1]
Bioreaktoripohjaisessa tuotannossa solut on myös muokattava kestämään hyperosmoottista stressiä ja ravinteiden puutetta. Ennen skaalausta on olennaista validoida geneettiset muokkaukset käyttämällä työkaluja, kuten Western blot tai FACS. Tutkijoille, jotka etsivät erikoistuneita solulinjoja tai geneettisiä materiaaleja, jotka on räätälöity tiheän bioreaktorin ympäristöihin, alustat kuten
Päätelmä
Anti-apoptoottisten geenien valinta viljellyille lihasolulinjoille vaatii räätälöityä lähestymistapaa. Geenit kuten BCL-2 , BCL-xL, ja MCL-1 jokainen näyttelevät ainutlaatuisia rooleja solujen suojaamisessa, mutta niiden menestys riippuu tekijöistä kuten solutyyppi, kehitysvaihe ja tuotannon aikana kohdatut erityiset stressit. Kuten tutkimuksessa korostetaan:
"tasapaino anti-apoptoottisten ja pro-apoptoottisten jäsenten välillä lopulta määrää, elääkö vai kuoleeko solu" [2]
Elossapysymisen lisäksi anti-apoptoottinen geenimuokkaus säilyttää myös aineenvaihduntatoiminnot. Esimerkiksi proteiinit kuten BCL-xL ovat läheisesti sidoksissa glykolyysin ja oksidatiivisen fosforylaation ylläpitämiseen. Kuitenkin huonosti toteutetut interventiot voivat häiritä näitä kriittisiä prosesseja [2]. Varmistamalla, että suunnitellut solulinjat säilyttävät tarkoitetun identiteettinsä ja aineenvaihduntatoimintansa koko tuotannon ajan, on ratkaiseva, vaikkakin joskus unohdettu vaihe. Nämä oivallukset muokkaavat solulinjojen suunnittelun tulevaisuutta.
Uudet monigeeniset lähestymistavat ovat nousemassa, jotka yhdistävät suojaavien geenien yliekspression CRISPR-poistojen kanssa pro-apoptoottisista geeneistä, kuten BAX, BAK1, ja BOK, luodakseen kestävämpiä solulinjoja teolliseen käyttöön [1]. Metabolisen profiloinnin työkalut, kuten bioenergeettiset testit, ovat tulossa välttämättömiksi vahvistamaan, että nämä geneettiset muutokset parantavat solujen kokonaisvaltaista suorituskykyä. Tutkijoille, jotka hankkivat sikojen solulinjoja, geneettisiä materiaaleja tai bioreaktorilaitteita,UKK
Minkä anti-apoptoottisen geenin kanssa minun pitäisi aloittaa solulinjalleni?
BCL-2 on usein suositeltu aloituspiste solulinjojen kanssa työskenneltäessä. Tämä hyvin tutkittu anti-apoptoottinen geeni tunnetaan kyvystään parantaa solujen selviytymistä, mikä tekee siitä suositun vaihtoehdon viljellyn lihan tutkimuksessa. Sen rooli solujen elinkelpoisuuden tukemisessa tekee siitä käytännöllisen valinnan alkuvaiheen kokeisiin.
Onko parempi yliekspressoida anti-apoptoottisia geenejä vai poistaa pro-apoptoottisia geenejä?
Viljellyn lihan tuotannossa anti-apoptoottisten geenien, kuten BCL-2-perheen jäsenten, kuten BCL-xL, ilmentämisen lisääminen tuottaa yleensä parempia tuloksia kuin pro-apoptoottisten geenien poistaminen. Tämä strategia tukee sekä solujen selviytymistä että lisääntymistä - keskeisiä tekijöitä tuotannon laajentamisessa - samalla kun säilytetään solun luonnolliset säätelyjärjestelmät.
Lisäämällä anti-apoptoottisten geenien aktiivisuutta solut saavat suuremman vastustuskyvyn apoptoosia vastaan, erityisesti stressaavissa olosuhteissa. Tämä tekee siitä luotettavamman ja turvallisemman lähestymistavan solujen elinkelpoisuuden ylläpitämiseksi viljelyprosessin aikana.
Kuinka voin varmistaa, että anti-apoptoottinen muokkaus parantaa IVCC:tä bioreaktorissani?
Määrittääksesi, parantaako anti-apoptoottinen geenimuokkaus in vitro solujen elinkelpoisuutta ja lisääntymistä (IVCC), tarvitset systemaattisen lähestymistavan:
- Arvioi elinkelpoisuus- ja lisääntymisnopeudet: Käytä menetelmiä, kuten solujen laskemista tai virtaussytometriaa, mitataksesi näitä nopeuksia sekä ennen geenimuokkausta että sen jälkeen.
- Vahvista geeniekspressio : Tekniikoilla, kuten qPCR tai Western blotting, voidaan varmistaa kohdegeenin onnistunut ilmentyminen.
- Seuraa apoptoosin markkereita: Tarkista markkerit, kuten kaspaasiaktiivisuus, varmistaaksesi, että muokkaus vähentää tehokkaasti apoptoosia.
Jotta arviointi olisi täydellinen, on tärkeää testata muokattujen solujen pitkäaikainen stabiilisuus ja lisääntyminen bioreaktorissa. Tämä varmistaa, että parannukset säilyvät useiden viljelysyklien ajan.










