

If you can expand cells but can’t switch them into the right fate at the right time, your process will stall at differentiation. That is the core point here: synthetic gene circuits give you inside-the-cell control over commitment, timing, memory and lineage mix, where media changes alone often leave heterogeneous, partly committed populations.
If I were building a cultivated meat differentiation workflow, I’d take four points from this article straight away:
- Start with the native network, not the construct. Use snRNA-seq, trajectory analysis, GRN inference and miRNA profiling to find where cells stall, drift or branch into the wrong fate.
-
Match circuit type to the process problem.
A toggle switch suits lock-in, a feedforward or band-pass design suits timing control, a logic gate suits multi-signal gating, and miSFITs suit graded output. - Design for low leak, low noise and safety from day one. Orthogonal parts, negative autoregulation, iFFLs, cm transgenes and an inducible kill or growth-arrest module are part of the build, not an afterthought.
- Validate under scale-relevant conditions early. A circuit that works in 2D may shift in 3D, microcarrier or stirred suspension because of inducer gradients, oxygen limits and shear.
The article also makes a practical point that matters for process teams: single-lineage control and ratio control are different jobs. A Tet-On MyoD cassette may push myogenic entry, but whole-cut products need control of muscle, fat and ECM proportions, which usually means feedback, paracrine signalling and heavier clone screening.
A few figures anchor that message:
- Standard myogenic differentiation can stall with fusion indices of roughly 50–60%
- Engineered GRNs in iPSCs increased target-lineage differentiation from 52% to 81%
- Synthetic circuits in modified MSCs drove cardiac differentiation to 76%
- Some porcine Tet-On-PAX7 lines kept high myogenic potential beyond 40 passages
- Around 20% of human pluripotent stem cells may carry cancer-linked mutations, which is why inducible safety modules matter
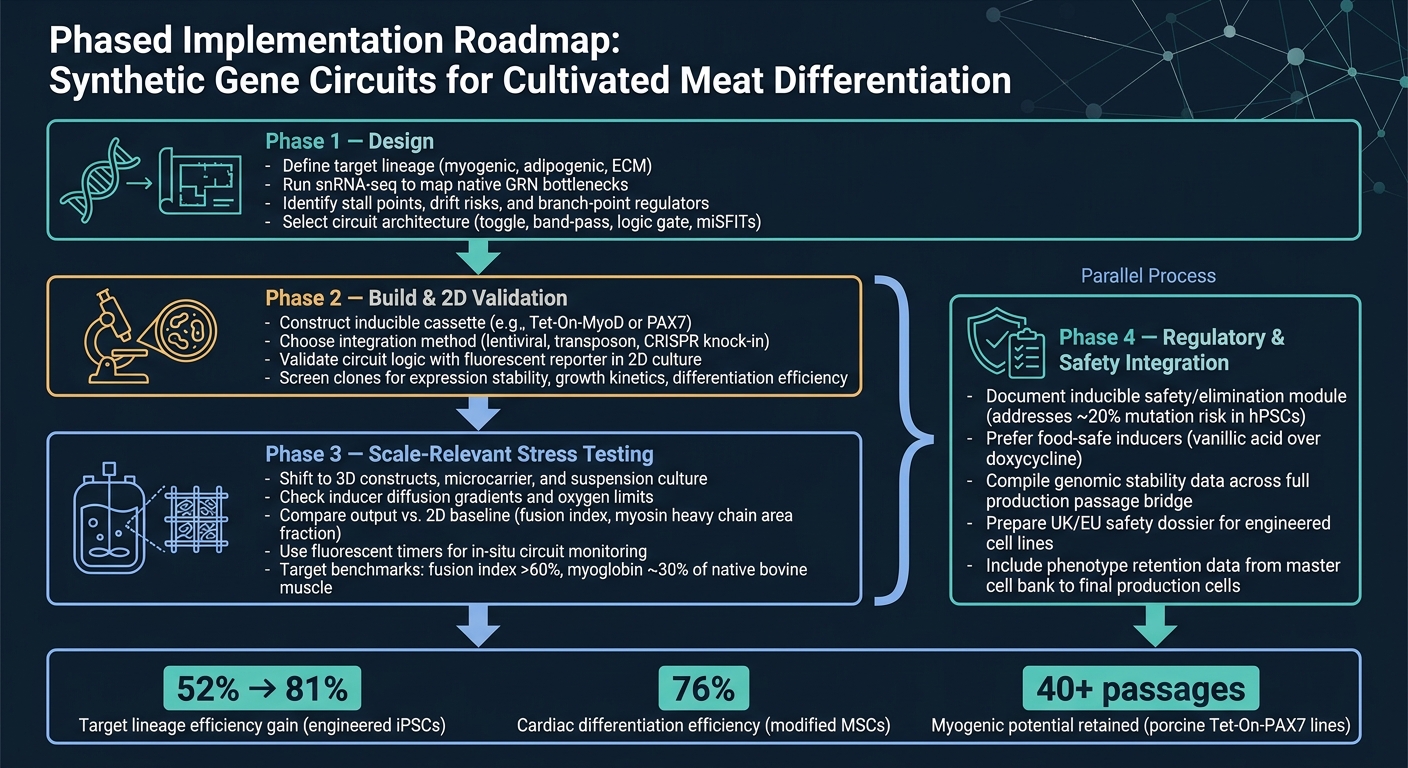
Synthetic Gene Circuit Workflow for Cultivated Meat Differentiation
Scientist Stories: Michael Elowitz, Genetic Circuits in Living Cells
Quick comparison
| Topic | What the article says in plain terms |
|---|---|
| Main bottleneck | Differentiation, not expansion |
| Why circuits help | They add threshold control and cell-state memory inside the cell |
| Best first step | Map native fate control with snRNA-seq and related tools |
| Main circuit options | Toggle, feedforward, band-pass, logic gates, miSFITs, CRISPRa/i |
| Myogenesis example | Tet-On-MyoD to separate growth from terminal differentiation |
| Adipogenesis / ECM control | miSFITs and bow-tie designs for graded PPARG/BMP4-type output |
| Whole-cut challenge | Ratio control across muscle, fat and connective tissue |
| Scale-up risk | 2D behaviour may not hold in 3D or bioreactors |
| Integration choices | Lentivirus, transposons, CRISPR knock-in, episomal vectors |
| Regulatory point | Engineered lines need a broader safety package; food-safe inducers such as vanillic acid are preferred over DOX where possible |
So, in simple terms: this is not just a circuit design article. I’d read it as a guide to linking circuit architecture, lineage biology, clone selection, bioreactor performance and UK/EU safety documentation into one differentiation strategy.
Read on if you want the full path from native network mapping to construct choice, scale-up checks and regulatory fit.
2. Design principles for differentiation-control circuits
2.1 Map the native cell-fate network before designing the circuit
Before you design a circuit, you need a clear picture of what the cell is already doing.
Single-nucleus RNA sequencing (snRNA-seq) is a good place to start. It can show quiescent subpopulations, including reserve cells marked by NOTCH2 and HEYL, and point to pathway targets that may improve differentiation [3].
From there, trajectory analysis and gene regulatory network (GRN) inference help map the order of regulator activation and highlight where cells are most likely to stall. In myogenesis, the main cascade runs through MYOD1 and MYOG. In adipogenesis, the main nodes are PPARG and CEBPA, with fibro-adipogenic progenitor (FAP) fate acting as the main branch-point risk. The table below summarises the main regulators and bottlenecks.
| Lineage | Key master regulators | Critical signalling pathways | Identified bottlenecks |
|---|---|---|---|
| Myogenic | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Reserve cell formation (quiescence) |
| Adipogenic | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Fibro-adipogenic progenitor (FAP) fate |
| Pluripotent | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Spontaneous differentiation / heterogeneity |
Another useful layer is miRNA expression profiling. Endogenous miRNAs such as miR-302a, linked to pluripotency, and miR-375, linked to differentiation, can act as internal classifiers in sense-and-respond designs. That lets the circuit read the cell’s actual state instead of depending only on external inducers [5].
Those bottlenecks should drive the circuit choice. If the main problem is drift, you may need a lock-in. If timing is the issue, a pulse may fit better. If fate control depends on more than one cue, multi-input logic usually makes more sense.
2.2 Choose the right circuit architecture
This is where the trade-offs show up in plain view. The right architecture depends on three practical points: how permanent the commitment must be, how tightly timing needs to be controlled, and how much genetic payload the cell can carry without trouble.
Bistable toggle switches fit cases where lineage commitment needs to stay locked. The main problem is spontaneous state flipping caused by transcriptional noise.
Band-pass filters fit cases where a transcription factor is needed only within a defined developmental window. The catch is that inducer levels need tight control, or the timing slips.
Logic gates add specificity by requiring more than one input at the same time. For example, differentiation can be allowed only when an exogenous inducer is present and the cell shows the right endogenous miRNA profile. That helps cut the risk of off-target commitment.
The table below outlines the main architectures and the trade-offs that come with them.
| Architecture | Reversibility | Temporal precision | Integration complexity | Primary use case | Key risk |
|---|---|---|---|---|---|
| Bistable switch | Low (once locked) | Moderate | Moderate | Permanent lineage commitment | Spontaneous flipping due to noise |
| Band-pass filter | High (concentration-dependent) | High | High | Transient developmental stages | Requires tight inducer control |
| Logic gate (AND/OR/NOT) | Variable | Moderate | Moderate–high | Cell-type-specific activation | Leakiness in OFF states |
| Bow-tie / multi-input | High | Moderate | Moderate | Multi-signal integration | Relies on endogenous miRNA stability |
| miSFITs | High | Moderate | Low–moderate | Graded output tuning | Narrow dynamic range if poorly tuned |
"By minimizing the number of computational layers while maintaining functionality, this strategy addresses scalability barriers in gene circuit engineering." - Nature Communications [9]
Each added regulatory layer increases payload and draws on cellular resources. In practice, if two designs do the same job, the simpler one is usually the better bet when scale matters.
Once the architecture is set, the next job is to make it hold up under low leakiness, noise suppression and fail-safe control.
2.3 Build for reliability, low leakiness and safety
A circuit has to stay stable over extended culture. A short burst of good performance is not enough for production use.
Orthogonal parts are the first defence. Promoters, transcription factors and regulatory elements that do not cross-react with native machinery help limit off-target effects and reduce the chance that endogenous signals will switch the circuit on or shut it down. Modified high-tightness promoters such as PCREm have been used to reduce basal leakiness in inducible mammalian systems [6].
Negative autoregulation is also worth adding where possible. It is one of the better-known motifs for reducing transcriptional noise and producing a more linear response to inducer concentration [6]. Incoherent feedforward loops (iFFLs) can add another layer of control by filtering stochastic fluctuations, so cells respond to sustained signals rather than short noise spikes.
Codon-modified (cm) versions of synthetic transcription factors make characterisation easier as well. They let you separate circuit-driven expression from endogenous genomic (g) expression during validation [1]. That may sound like a small detail, but it saves time when you're trying to work out whether a readout comes from the circuit or the host genome.
Safety modules are required. Around 20% of human pluripotent stem cells carry cancer-associated mutations [7]. So, if a circuit is going into a stem-cell-derived line, it should include an inducible growth-arrest or elimination module. Vanillic acid is a useful inducer to prioritise here because it is a licensed food additive, which helps the case for using it as a circuit trigger in cultivated meat cell lines [1].
"Synthetic biology provides engineers with the means to deploy circuits to easily and precisely tune the expression of multiple genes to... eliminate potential adverse off-target side effects." - npj Systems Biology and Applications [6]
These choices set up the lineage-specific circuits in Section 3.
sbb-itb-ffee270
3. Circuit strategies for myogenic, adipogenic and ratio-controlled differentiation
3.1 Myogenic circuits that separate expansion from terminal differentiation
Once the circuit architecture is set, the next job is lineage-specific deployment. For myogenesis, the main problem is simple to state but hard to execute: cells need to proliferate first, then switch into differentiation when asked, without drifting too early in either direction.
A Tet-On-MyoD cassette is one of the most direct ways to do this. In this setup, cells proliferate under standard conditions when doxycycline (DOX) is absent. Add DOX, and the circuit drives myogenic commitment. Researchers at multiple Chinese institutions used this approach in chicken embryonic fibroblasts and reported efficient myotube formation after induction [4].
A bistable toggle switch offers tighter state control. Systems built from mutually inhibitory repressors such as E-KRAB and Pip-KRAB can turn a short DOX pulse into a stable myogenic programme [6]. In practice, that means the differentiated state stays locked in after induction rather than fading once the signal is removed. Adding negative autoregulation also helps cut stochastic noise and limit leaky differentiation during the expansion phase [6].
This matters because standard in vitro myogenic differentiation often stalls part-way. Fusion indices are usually only around 50–60%, which leaves a large non-fusing reserve population [3]. Circuit-driven MyoD activation can improve commitment, but commitment alone does not guarantee uniform fusion. When MyoD circuits are combined with MEK, NOTCH and RXR modulation, near-complete fusion in 2D culture becomes possible [3]. The circuit controls the timing of lineage entry; the small molecules push a cleaner and more uniform cell-cycle exit.
3.2 Adipogenic and fibrogenic control for composition and structure
Muscle alone is not enough. Once myogenic control is in place, the next issue is composition: how much fat forms, how much ECM is deposited, and when those programmes switch on. Here, binary ON/OFF control is often too blunt. What teams usually need is graded output, especially around the adipogenic-fibrogenic branch point.
miSFITs give a practical way to tune expression in steps. By placing mutated miRNA target sites - for example, sites for miR-17 - in the 3′UTR of output genes such as PPARG or BMP4, researchers can select expression levels from a variant library [5]. That makes adipocyte induction more like a dimmer switch than a light switch. Instead of pushing cells into an all-or-nothing response, teams can titrate adipogenesis more carefully [5].
Fibroblasts are also not just bystanders here. They supply ECM proteins that shape texture [10]. That makes fibrogenic control part of product design, not just a side issue. Circuits can help manage the shift between fibrogenic and adipogenic states, and in cultivated poultry, direct PPARG activation in fibroblasts may be needed to generate meaningful fat deposition [10].
A bow-tie architecture fits this problem well because it keeps sensing and output separate. The sensing layer reads the cell’s current state, while the output layer adjusts PPARG, CEBPA or other lineage regulators. That separation helps stop adipogenic or connective tissue programmes from turning on before cells have reached the right developmental stage.
3.3 Multi-lineage ratio control and feedback sensing
Ratio-control circuits deal with a different problem: not whether differentiation happens, but whether the final population mix stays where it should. For whole-cut products, getting muscle, fat and ECM into the right proportions matters just as much as getting any one lineage to differentiate.
These systems build feedback control into the cells themselves. State-specific promoters restrict signalling protein expression to cells that have already committed to a given lineage. Synthetic paracrine modules then let committed myogenic cells release an inhibitory signal that suppresses adipogenic commitment in nearby cells. The logic resembles lateral inhibition in Delta-Notch developmental systems [1][6]. Where branching gets more complicated, multi-input gates can combine extracellular cues with internal state signals [9].
miSFITs also work at this stage. By tuning the output strength of BMP4 or other morphogens, teams can shift lineage balance without rewriting the upstream decision logic. In engineered gene regulatory networks, this type of control increased target lineage differentiation efficiency from 52% in controls to 81% in engineered iPSCs [2]. In modified mesenchymal stem cells, synthetic circuits drove cardiac differentiation efficiency to 76% [2].
The table below compares single-lineage and ratio-control approaches using the points that matter most in production.
| Feature | Single-lineage circuit | Ratio-control circuit |
|---|---|---|
| Complexity | Low; typically a single inducible promoter and regulator [4] | High; requires logic gates and paracrine signalling [6][9] |
| Monitoring burden | Low; usually follows a single reporter [4] | High; requires tracking multiple lineage markers [5] |
| Robustness | Moderate; prone to heterogeneity and reserve cell formation [3] | High; uses feedback and lateral inhibition to balance populations [1] |
| Production value | High for biomass; limited for complex tissue structure [10] | Essential for whole-cut products needing muscle, fat and ECM [4] |
Ratio-control circuits add a heavier validation load. But their built-in feedback is hard to match with process control alone, which puts more pressure on clone selection and process testing.
4. From construct to process: validation, scale-up and regulatory fit
4.1 Integration strategy and clone selection for stable performance
After circuit design, the hard part starts: getting that circuit into cells in a way that stays stable through production-scale passaging.
Lentiviral delivery is often efficient and gives stable integrants fast. But integration is random. That means more regulatory attention, plus a risk that expression drops over time because of silencing. Transposon systems such as PiggyBac and Sleeping Beauty sit in the middle. They can hold performance across many passages, but you still need to screen for copy number and insertion sites. Sleeping Beauty, for example, has been used to stably immortalise bovine satellite cells by overexpressing TERT and CDK4, with lines retaining myogenic potential beyond 40 passages [10]. CRISPR knock-in gives the tightest control over where the construct lands and the most precise genomic setting, although clone picking is slower and throughput is lower.
| Integration Method | Insertion Control | Stability | Scalability | Regulatory Considerations |
|---|---|---|---|---|
| Lentiviral Delivery | Low (random integration) | High, but prone to silencing | High | Greater regulatory scrutiny due to random insertion and viral remnants |
| Transposons (PiggyBac/SB) | Moderate | High over many passages | High | Requires screening for copy number and insertion sites |
| CRISPR Knock-in | High (site-specific) | Very high | Moderate | Favourable; reduces risk of disrupting endogenous genes |
| Episomal Vectors | None (extrachromosomal) | Low; can be lost during division | Low | Lower integration hurdle, but unsuitable for long-term expansion |
Clone screening needs to do more than confirm the construct is present. It should track expression drift, insertion profile, growth kinetics, differentiation efficiency, and phenotype retention at high passage number. snRNA-seq is useful here because it can rule out clones enriched for Pax7⁺/Ki-67⁻ reserve cells - cells that leave the cell cycle without differentiating - before scale-up [3]. Porcine EPSCs with a Tet-On-PAX7 circuit retained high muscle differentiation in 3D microcarrier and suspension culture over 40 passages [8].
4.2 How circuit behaviour changes in 3D, microcarrier and bioreactor culture
Once you have a clone, the next test is whether it behaves the same way outside 2D. In many cases, it doesn't. Performance in 2D rarely carries across cleanly to suspension, microcarrier, or scaffold culture because diffusion gradients, oxygen limits, and shear all shift circuit output.
One of the first checks is inducer diffusion. In stirred-tank bioreactors, small-molecule inducers need to reach cells evenly. In practice, gradients can form, especially in dense microcarrier cultures and inside aggregates or scaffold cores. Suspension culture is usually a better fit for large-scale cultivated meat because it supports higher cell density and gives tighter process control.
Monitoring cell state also gets harder as the system scales. Fluorescent reporter signals that are easy to read by microscopy in 2D can become difficult to resolve in opaque 3D constructs. Fluorescent timers - probes that shift emission from blue to red as a protein matures - can give real-time circuit-activation data in situ [1]. The sensible route is staged validation: first in 2D, then in 3D constructs, then under the final bioreactor conditions [3][8].
4.3 Characterisation, food-safety documentation and UK/EU considerations
After process testing, characterisation has to show that circuit function, phenotype, and safety still hold up. The core data stack should include flow cytometry, qPCR with cm sequences, RNA-seq time courses, and functional readouts such as myosin heavy chain area fraction and myoglobin expression [1]. Optimised serum-free differentiation media have been shown to bring myoglobin expression to approximately 30% of levels found in native bovine muscle [3]. That gives teams a clear benchmark rather than a vague target.
You also need to document protein, amino acid, and fat profiles, along with sensory traits [10][3].
From a regulatory point of view, the UK and EU draw a clear line between spontaneously immortalised (non-GMO) cell lines and genetically engineered lines. The latter need broader safety dossiers [10][3]. Stability packages should show phenotype retention and genomic stability across the full production bridge - from master cell bank to final production cells - and traceability records need to account for every passage in between [10]. If the circuit depends on a chemical inducer, a food-safe or licensed additive such as vanillic acid is preferable to doxycycline [1].
Routine genomic surveillance is a must, and an inducible suicide or elimination switch should be documented as a core risk-control measure [7]. Its function should also be included in the safety dossier, especially as UK and EU rules for cultivated meat continue to take shape.
5. Practical roadmap and conclusion
5.1 A phased implementation roadmap for cultivated meat teams
The cleanest route from concept to production is a staged workflow.
Phase 1 is design. Start by defining the target lineage, then use snRNA-seq to confirm the main bottlenecks before you pick a circuit architecture. That step matters because a circuit can only solve the constraints you’ve actually identified.
Phase 2 is build and 2D validation. Build the construct and check that the circuit behaves as intended in 2D, using a simple reporter readout. At this stage, the goal is straightforward: confirm the logic works before moving into harder and more costly models.
Phase 3 is scale-relevant stress-testing. Shift into 3D systems and bioreactor-relevant conditions, then compare output against the 2D baseline. This is where many designs start to show their weak points, especially when mass transfer, shear, and matrix effects come into play.
Phase 4 is regulatory and safety integration, and it should run in parallel with Phase 3. Safety and regulatory work shouldn’t wait until the end. Run it alongside scale-up, including documentation for any inducible safety module.
5.2 Sourcing enabling tools and materials through Cellbase
Once the workflow is set, sourcing often becomes the rate-limiting step.
Cellbase is a specialised B2B marketplace built only for the cultivated meat industry. Procurement teams and R&D scientists can use Cellbase to find verified suppliers for the tools and materials that synthetic circuit workflows depend on, including:
- cell lines
- serum-free and chemically defined media
- scaffolds
- bioreactor components
- sensors
- analytical equipment
Reliable access to compatible materials at each stage has a direct effect on how fast circuit behaviour can be characterised under scale-relevant conditions.
5.3 Key takeaways
Synthetic gene circuits give cultivated meat teams programmable control over timing, thresholds, and lineage balance that media-only protocols cannot match. Architecture choice shapes reversibility, leakiness, and safety. Inducible systems are usually preferred because they provide conditional control and a lower metabolic load [6].
"The synthetic biology toolkit can be used to establish cell lines with tuneable gene expression, which, when coupled with PAT and computational modelling, can enable closed-loop control systems to deliver optimal product yield and quality." - npj Systems Biology and Applications [6]
Successful deployment is not a biology problem alone. It depends on tight coupling between circuit engineering, bioprocess design, regulatory documentation, and procurement.
FAQs
How do synthetic gene circuits improve differentiation consistency?
Synthetic gene circuits can make differentiation more consistent because they give you programmed control over cell behaviour and lineage commitment. In practice, that means using modular logic operations to tune gene and transcription factor expression with tight timing.
That timing matters. It helps cells move through defined state changes in the right order, instead of drifting into mixed or unwanted fates. It also cuts down off-target differentiation and reduces noise across the culture.
The payoff is straightforward: more uniform, stable, and mature cell populations for cultivated meat production.
Which circuit design suits myogenic or adipogenic control?
In cultivated meat research, the same chicken fibroblasts can be pushed into either lineage. Myogenesis follows one set of induction protocols, while adipogenesis can be switched on by exposing the cells to inputs such as chicken serum or fatty acids.
From there, these cell fates can be controlled step by step inside 3D hydrogel scaffolds to build meat structures with defined fat and collagen ratios.
Why do gene circuits often behave differently in 3D culture?
In 3D culture, gene circuits often behave differently because cells are dealing with physical and structural inputs that simply don't exist in 2D monolayers. Those inputs include mechanical tension, shear stress, matrix stiffness, and local cell density.
These cues can shift signalling pathways such as Notch. They can also change how synthetic circuits detect force and coordinate downstream responses, including cell-cell adhesion and tissue morphogenesis.









