

Jeśli możesz rozszerzać komórki, ale nie możesz przełączyć ich w odpowiedni los w odpowiednim czasie, twój proces zatrzyma się na etapie różnicowania. To jest kluczowy punkt: syntetyczne obwody genowe dają ci wewnątrzkomórkową kontrolę nad zaangażowaniem, czasem, pamięcią i mieszanką linii, gdzie same zmiany w mediach często pozostawiają heterogeniczne, częściowo zaangażowane populacje.
Gdybym budował przepływ pracy różnicowania mięsa hodowanego, od razu wziąłbym cztery punkty z tego artykułu:
- Zacznij od natywnej sieci, a nie od konstruktu. Użyj analizy trajektorii snRNA-seq, , wnioskowania GRN i profilowania miRNA, aby znaleźć, gdzie komórki się zatrzymują, dryfują lub rozgałęziają w niewłaściwy los.
-
Dopasuj typ obwodu do problemu procesu.
Przełącznik toggle switch pasuje do blokady, projekt feedforward lub pasmowo-przepustowy pasuje do kontroli czasu, bramka logiczna pasuje do bramkowania wielosygnałowego, a miSFITs pasują do stopniowanego wyjścia. - Projektuj z myślą o niskim wycieku, niskim hałasie i bezpieczeństwie od pierwszego dnia. Części ortogonalne, negatywna autoregulacja, iFFL, transgeny cm oraz indukowalny moduł zabijania lub zatrzymania wzrostu są częścią konstrukcji, a nie dodatkiem.
- Waliduj w warunkach odpowiednich dla skali na wczesnym etapie. Obwód, który działa w 2D, może się zmienić w 3D, na mikronośniku lub w zawiesinie mieszanej z powodu gradientów induktora, ograniczeń tlenu i ścinania.
Artykuł zwraca również uwagę na praktyczny aspekt istotny dla zespołów procesowych: kontrola pojedynczej linii i kontrola proporcji to różne zadania. Kaseta Tet-On MyoD może wspierać wejście miogeniczne, ale produkty cięte w całości wymagają kontroli proporcji mięśni, tłuszczu i ECM, , co zazwyczaj oznacza sprzężenie zwrotne, sygnalizację parakrynną i bardziej intensywne badanie klonów.
Kilka liczb ilustruje tę wiadomość:
- Standardowe różnicowanie miogenne może zatrzymać się przy współczynnikach fuzji około 50–60%
- Inżynieryjne GRN w iPSC zwiększyły różnicowanie do docelowej linii z 52% do 81%
- Syntetyczne obwody w zmodyfikowanych MSC napędzały różnicowanie sercowe do 76%
- Niektóre linie porcine Tet-On-PAX7 utrzymywały wysoką potencjał miogenny powyżej 40 pasaży
- Około 20% ludzkich pluripotencjalnych komórek macierzystych może nosić mutacje związane z rakiem, dlatego moduły bezpieczeństwa indukowane są ważne
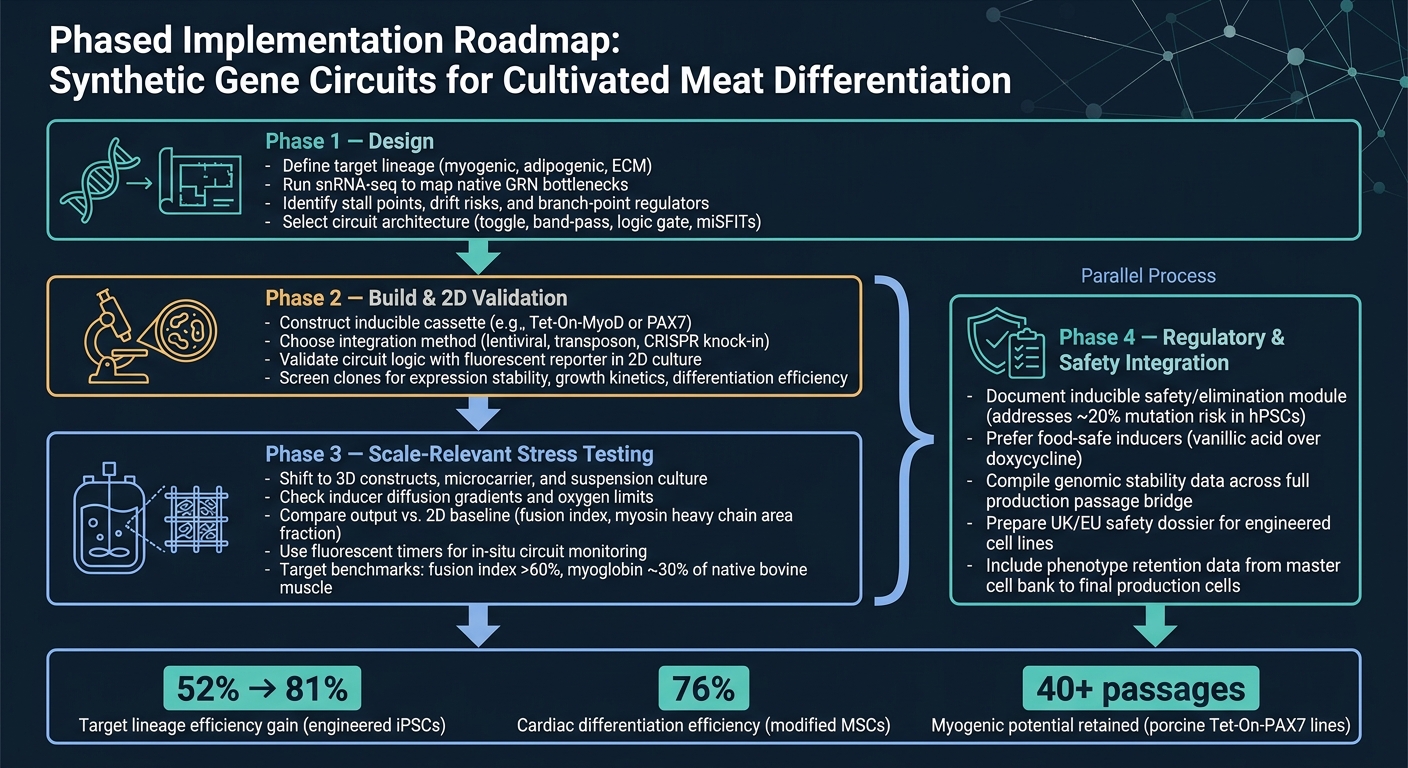
Przepływ pracy syntetycznych obwodów genowych dla różnicowania mięsa hodowlanego
Historie naukowców: Michael Elowitz, obwody genetyczne w żywych komórkach
Szybkie porównanie
| Temat | Co artykuł mówi w prostych słowach |
|---|---|
| Główne wąskie gardło | Różnicowanie, nie ekspansja |
| Dlaczego obwody pomagają | Dodają kontrolę progową i pamięć stanu komórki wewnątrz komórki |
| Najlepszy pierwszy krok | Mapowanie natywnej kontroli losu za pomocą snRNA-seq i pokrewnych narzędzi |
| Główne opcje obwodów | Przełącznik, sprzężenie do przodu, pasmowo-przepustowy, bramki logiczne, miSFITs, CRISPRa/i |
| Przykład miogenezy | Tet-On-MyoD do oddzielenia wzrostu od końcowego różnicowania |
| Adipogeneza / kontrola ECM | miSFITs i projekty kokardowe dla stopniowanego wyjścia typu PPARG/BMP4 |
| Wyzwanie całkowitego cięcia | Kontrola proporcji w mięśniach, tłuszczu i tkance łącznej |
| Ryzyko skalowania | Zachowanie 2D może nie być takie samo w 3D lub bioreaktorach |
| Wybory integracyjne | Lentivirus, transpozony, CRISPR knock-in, wektory episomalne |
| Punkt regulacyjny | Linie inżynieryjne potrzebują szerszego pakietu bezpieczeństwa; induktory bezpieczne dla żywności, takie jak kwas wanilinowy, są preferowane nad DOX, gdzie to możliwe |
Więc, mówiąc prosto: to nie jest tylko artykuł o projektowaniu obwodów. Czytałbym to jako przewodnik po łączeniu architektury obwodów, biologii linii, wyboru klonów, wydajności bioreaktorów i dokumentacji bezpieczeństwa UK/UE w jedną strategię różnicowania.
Czytaj dalej, jeśli chcesz poznać pełną ścieżkę od mapowania sieci natywnej do wyboru konstrukcji, kontroli skalowania i dopasowania regulacyjnego.
2. Zasady projektowania obwodów kontrolujących różnicowanie
2.1 Zmapuj natywną sieć losów komórek przed zaprojektowaniem obwodu
Zanim zaprojektujesz obwód, potrzebujesz jasnego obrazu tego, co komórka już robi.
Sekwencjonowanie RNA pojedynczego jądra (snRNA-seq) to dobry punkt wyjścia. Może pokazać subpopulacje w stanie spoczynku, w tym komórki rezerwowe oznaczone przez NOTCH2 i HEYL , i wskazać cele ścieżek, które mogą poprawić różnicowanie [3].
Stamtąd analiza trajektorii i wnioskowanie sieci regulacyjnych genów (GRN) pomagają mapować kolejność aktywacji regulatorów i wskazują, gdzie komórki najprawdopodobniej się zatrzymują. W miogenezie główna kaskada przebiega przez MYOD1 i MYOG. W adipogenezie główne węzły to PPARG i CEBPA, z losem progenitorów fibro-adipogennych (FAP) działającym jako główny punkt ryzyka rozgałęzienia. Poniższa tabela podsumowuje głównych regulatorów i wąskie gardła.
| Linia | Kluczowi regulatorzy główni | Krytyczne szlaki sygnalizacyjne | Zidentyfikowane wąskie gardła |
|---|---|---|---|
| Miogeniczny | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Tworzenie komórek rezerwowych (spoczynkowych) |
| Adipogeniczny | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Los progenitorów fibro-adipogennych (FAP) |
| Pluripotencjalny | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Spontaniczne różnicowanie / heterogeniczność |
Kolejną przydatną warstwą jest profilowanie ekspresji miRNA. Endogenne miRNA, takie jak miR-302a, związane z pluripotencją, oraz miR-375, związane z różnicowaniem, mogą działać jako wewnętrzne klasyfikatory w projektach typu sense-and-respond. To pozwala obwodowi odczytać rzeczywisty stan komórki zamiast polegać wyłącznie na zewnętrznych induktorach [5].
Te wąskie gardła powinny kierować wyborem obwodu. Jeśli głównym problemem jest dryf, może być potrzebna blokada. Jeśli problemem jest czas, lepiej może pasować impuls. Jeśli kontrola losu zależy od więcej niż jednej wskazówki, logika wielo-wejściowa zwykle ma więcej sensu.
2.2 Wybierz odpowiednią architekturę obwodu
To tutaj kompromisy stają się widoczne. Odpowiednia architektura zależy od trzech praktycznych punktów: jak trwałe musi być zobowiązanie, jak ściśle trzeba kontrolować czas i jak duży ładunek genetyczny komórka może przenosić bez problemu.
Przełączniki bistabilne pasują do przypadków, w których zobowiązanie linii musi pozostać zablokowane. Głównym problemem jest spontaniczne przełączanie stanu spowodowane szumem transkrypcyjnym.
Filtry pasmowo-przepustowe pasują do przypadków, gdy czynnik transkrypcyjny jest potrzebny tylko w określonym oknie rozwojowym. Haczyk polega na tym, że poziomy induktora muszą być ściśle kontrolowane, w przeciwnym razie czas się przesuwa.
Bramki logiczne dodają specyficzności, wymagając więcej niż jednego wejścia jednocześnie. Na przykład, różnicowanie może być dozwolone tylko wtedy, gdy obecny jest egzogenny induktor i komórka wykazuje odpowiedni profil endogennego miRNA. To pomaga zmniejszyć ryzyko niecelowego zaangażowania.
Poniższa tabela przedstawia główne architektury i kompromisy, które się z nimi wiążą.
| Architektura | Odwracalność | Precyzja czasowa | Złożoność integracji | Główne zastosowanie | Kluczowe ryzyko |
|---|---|---|---|---|---|
| Przełącznik bistabilny | Niskie (po zablokowaniu) | Umiarkowane | Umiarkowane | Stałe zaangażowanie linii komórkowej | Spontaniczne przełączanie z powodu szumów |
| Filtr pasmowy | Wysokie (zależne od stężenia) | Wysokie | Wysokie | Przejściowe etapy rozwojowe | Wymaga ścisłej kontroli induktora |
| Bramka logiczna (AND/OR/NOT) | Zmienna | Umiarkowane | Umiarkowane–wysokie | Aktywacja specyficzna dla typu komórki | Przecieki w stanach OFF |
| Muszka / multiwejściowy | Wysoki | Umiarkowany | Umiarkowany | Integracja wielosygnałowa | Opiera się na stabilności endogennego miRNA |
| miSFITs | Wysoki | Umiarkowany | Niski–umiarkowany | Stopniowe dostrajanie wyjścia | Wąski zakres dynamiczny, jeśli źle dostrojony |
"Minimalizując liczbę warstw obliczeniowych przy jednoczesnym zachowaniu funkcjonalności, ta strategia rozwiązuje bariery skalowalności w inżynierii obwodów genowych." - Nature Communications [9]
Każda dodana warstwa regulacyjna zwiększa ładunek i wykorzystuje zasoby komórkowe. W praktyce, jeśli dwa projekty wykonują to samo zadanie, prostszy jest zazwyczaj lepszym wyborem, gdy skala ma znaczenie.
Gdy architektura jest ustalona, kolejnym zadaniem jest zapewnienie jej stabilności przy niskiej przeciekalności, tłumieniu szumów i kontroli awaryjnej.
2.3 Buduj dla niezawodności, niskiej przeciekalności i bezpieczeństwa
Obwód musi pozostać stabilny przez dłuższy czas hodowli. Krótkotrwały dobry wynik nie wystarcza do zastosowań produkcyjnych.
Części ortogonalne są pierwszą linią obrony. Promotory, czynniki transkrypcyjne i elementy regulacyjne, które nie reagują krzyżowo z natywną maszynerią, pomagają ograniczyć efekty uboczne i zmniejszyć szansę, że endogenne sygnały włączą lub wyłączą obwód.Zmodyfikowane promotory o wysokiej szczelności, takie jak PCREm, zostały użyte do zmniejszenia podstawowego wycieku w indukowalnych systemach ssaków [6] .
Negatywna autoregulacja również jest warta dodania tam, gdzie to możliwe. Jest to jeden z bardziej znanych motywów redukujących szum transkrypcyjny i zapewniających bardziej liniową odpowiedź na stężenie induktora [6]. Niespójne pętle sprzężenia zwrotnego (iFFLs) mogą dodać kolejną warstwę kontroli poprzez filtrowanie stochastycznych fluktuacji, dzięki czemu komórki reagują na utrzymujące się sygnały, a nie na krótkie szczyty szumu.
Wersje zmodyfikowane kodonowo (cm) syntetycznych czynników transkrypcyjnych również ułatwiają charakteryzację. Pozwalają one oddzielić ekspresję napędzaną przez obwód od endogennej ekspresji genomowej ( g) podczas walidacji [1]. To może wydawać się drobnym szczegółem, ale oszczędza czas, gdy próbujesz ustalić, czy odczyt pochodzi z obwodu, czy z genomu gospodarza.
Moduły bezpieczeństwa są wymagane. Około 20% ludzkich pluripotencjalnych komórek macierzystych nosi mutacje związane z rakiem [7]. Dlatego, jeśli obwód ma być wprowadzony do linii pochodzącej z komórek macierzystych, powinien zawierać indukowalny moduł zatrzymania wzrostu lub eliminacji. Kwas wanilinowy jest tutaj użytecznym induktorem do priorytetyzacji, ponieważ jest licencjonowanym dodatkiem do żywności, co wspiera jego użycie jako wyzwalacza obwodu w liniach komórek mięsa hodowlanego [1].
"Biologia syntetyczna zapewnia inżynierom środki do wdrażania obwodów w celu łatwego i precyzyjnego dostrajania ekspresji wielu genów, aby... wyeliminować potencjalne niekorzystne skutki uboczne poza celem." - npj Systems Biology and Applications [6]
Te wybory ustanawiają specyficzne dla linii obwody w Sekcji 3.
sbb-itb-ffee270
3. Strategie obwodów dla różnicowania miogenicznego, adipogennego i kontrolowanego stosunku
3.1 Obwody miogeniczne, które oddzielają ekspansję od terminalnego różnicowania
Gdy architektura obwodu jest ustalona, następnym zadaniem jest specyficzne dla linii wdrożenie. Dla miogenezy główny problem jest prosty do sformułowania, ale trudny do wykonania: komórki muszą najpierw proliferować, a następnie przełączyć się na różnicowanie na żądanie, bez zbyt wczesnego dryfowania w którymkolwiek kierunku.
Kasetka Tet-On-MyoD jest jednym z najbardziej bezpośrednich sposobów na to. W tym układzie komórki proliferują w standardowych warunkach, gdy brak jest doksycykliny (DOX). Dodaj DOX, a obwód napędza zaangażowanie miogeniczne.Badacze z wielu chińskich instytucji zastosowali to podejście w fibroblastach zarodkowych kurcząt i zgłosili wydajną formację miotub po indukcji [4].
Dwustanowy przełącznik bistabilny oferuje ściślejszą kontrolę stanu. Systemy zbudowane z wzajemnie hamujących represorów, takich jak E-KRAB i Pip-KRAB, mogą przekształcić krótki impuls DOX w stabilny program miogeniczny [6]. W praktyce oznacza to, że stan zróżnicowany pozostaje zablokowany po indukcji, zamiast zanikać po usunięciu sygnału. Dodanie negatywnej autoregulacji również pomaga zmniejszyć szum losowy i ograniczyć przecieki różnicowania podczas fazy ekspansji [6] .
To ma znaczenie, ponieważ standardowe różnicowanie miogeniczne in vitro często zatrzymuje się w połowie. Wskaźniki fuzji wynoszą zwykle tylko około 50–60%, co pozostawia dużą populację rezerwową, która się nie łączy [3]. Aktywacja MyoD sterowana obwodami może poprawić zaangażowanie, ale samo zaangażowanie nie gwarantuje jednolitego połączenia. Gdy obwody MyoD są połączone z modulacją MEK, NOTCH i RXR, możliwe staje się niemal całkowite połączenie w kulturze 2D [3]. Obwód kontroluje czas wejścia w linię; małe cząsteczki wspomagają czystsze i bardziej jednolite wyjście z cyklu komórkowego.
3.2 Kontrola adipogenna i fibrogenna dla składu i struktury
Sam mięsień nie wystarczy. Gdy kontrola miogeniczna jest już na miejscu, kolejnym problemem jest skład: ile tłuszczu się tworzy, ile ECM jest odkładane i kiedy te programy się włączają. Tutaj, binarna kontrola WŁ./WYŁ. jest często zbyt prymitywna. To, czego zespoły zwykle potrzebują, to stopniowane wyjście, zwłaszcza w punkcie rozgałęzienia adipogenno-fibrogennego.
miSFITs dają praktyczny sposób na stopniowe dostrajanie ekspresji.Poprzez umieszczenie zmutowanych miejsc docelowych miRNA - na przykład miejsc dla miR-17 - w 3′UTR genów wyjściowych, takich jak PPARG lub BMP4, badacze mogą wybierać poziomy ekspresji z biblioteki wariantów [5]. To sprawia, że indukcja adipocytów jest bardziej jak ściemniacz niż przełącznik światła. Zamiast popychać komórki do reakcji typu wszystko albo nic, zespoły mogą bardziej precyzyjnie dawkować adipogenezę [5].
Fibroblasty również nie są tutaj tylko obserwatorami. Dostarczają białka ECM, które kształtują teksturę [10]. To sprawia, że kontrola fibrogeniczna jest częścią projektowania produktu, a nie tylko kwestią poboczną. Obwody mogą pomóc zarządzać przejściem między stanami fibrogenicznymi i adipogenicznymi, a w hodowanym drobiu, bezpośrednia aktywacja PPARG w fibroblastach może być potrzebna do uzyskania znaczącego odkładania tłuszczu [10].
Architektura typu muszka dobrze pasuje do tego problemu, ponieważ oddziela warstwę sensoryczną od wyjściowej. Warstwa sensoryczna odczytuje aktualny stan komórki, podczas gdy warstwa wyjściowa dostosowuje PPARG, CEBPA lub inne regulatory linii. To oddzielenie pomaga zapobiegać włączaniu się programów adipogennych lub tkanki łącznej, zanim komórki osiągną odpowiedni etap rozwoju.
3.3 Kontrola stosunku wieloliniowego i sprzężenie zwrotne
Obwody kontroli stosunku zajmują się innym problemem: nie tym, czy różnicowanie zachodzi, ale czy ostateczna mieszanka populacji pozostaje tam, gdzie powinna. Dla produktów typu whole-cut, uzyskanie odpowiednich proporcji mięśni, tłuszczu i ECM jest tak samo ważne, jak doprowadzenie do różnicowania dowolnej linii.
Te systemy wbudowują kontrolę sprzężenia zwrotnego w same komórki. Promotory specyficzne dla stanu ograniczają ekspresję białek sygnalizacyjnych do komórek, które już zobowiązały się do danej linii.Syntetyczne moduły parakrynne pozwalają następnie zaangażowanym komórkom miogennym uwalniać sygnał hamujący, który tłumi zaangażowanie adipogenne w pobliskich komórkach. Logika przypomina inhibicję boczną w systemach rozwojowych Delta-Notch [1][6]. Gdy rozgałęzienie staje się bardziej skomplikowane, bramki wielowejściowe mogą łączyć sygnały zewnątrzkomórkowe z sygnałami stanu wewnętrznego [9].
miSFITs działają również na tym etapie. Poprzez dostosowanie siły wyjściowej BMP4 lub innych morfogenów, zespoły mogą przesuwać równowagę linii bez przepisania logiki decyzji w górę strumienia. W inżynieryjnych sieciach regulacji genów, tego typu kontrola zwiększyła efektywność różnicowania docelowej linii z 52% w kontrolach do 81% w inżynieryjnych iPSC [2]. W zmodyfikowanych mezenchymalnych komórkach macierzystych, syntetyczne obwody zwiększyły efektywność różnicowania kardiologicznego do 76% [2].
Tabela poniżej porównuje podejścia z pojedynczym rodowodem i kontrolą proporcji, używając najważniejszych punktów w produkcji.
| Funkcja | Obwód jednoliniowy | Obwód z kontrolą stosunku |
|---|---|---|
| Złożoność | Niska; zazwyczaj pojedynczy promotor indukowalny i regulator [4] | Wysoka; wymaga bramek logicznych i sygnalizacji parakrynnej [6][9] |
| Obciążenie monitorowania | Niskie; zazwyczaj śledzi pojedynczy reporter [4] | Wysokie; wymaga śledzenia wielu markerów linii [5] |
| Odporność | Umiarkowana; podatna na heterogeniczność i tworzenie komórek rezerwowych [3] | Wysoki; wykorzystuje sprzężenie zwrotne i hamowanie boczne do równoważenia populacji [1] |
| Wartość produkcji | Wysoka dla biomasy; ograniczona dla złożonej struktury tkankowej [10] | Niezbędne dla produktów z całych kawałków wymagających mięśni, tłuszczu i ECM [4] |
Obwody kontrolujące proporcje dodają większe obciążenie walidacyjne.Ale ich wbudowana informacja zwrotna jest trudna do dopasowania tylko za pomocą kontroli procesu, co zwiększa presję na wybór klonów i testowanie procesów.
4. Od konstrukcji do procesu: walidacja, skalowanie i dopasowanie regulacyjne
4.1 Strategia integracji i wybór klonów dla stabilnej wydajności
Po zaprojektowaniu obwodu zaczyna się trudna część: wprowadzenie tego obwodu do komórek w sposób, który pozostaje stabilny podczas pasażowania na skalę produkcyjną.
Dostarczenie lentiwirusowe jest często wydajne i szybko daje stabilne integranty. Ale integracja jest losowa. To oznacza większą uwagę regulacyjną, a także ryzyko, że ekspresja spadnie z czasem z powodu wyciszenia. Systemy transpozonowe takie jak PiggyBac i Sleeping Beauty znajdują się pośrodku. Mogą utrzymać wydajność przez wiele pasaży, ale nadal trzeba przeprowadzić screening liczby kopii i miejsc insercji. Śpiąca Królewna , na przykład, została użyta do stabilnego immortalizowania komórek satelitarnych bydła poprzez nadekspresję TERT i CDK4, z liniami zachowującymi potencjał miogeniczny powyżej 40 pasaży [10]. CRISPR knock-in zapewnia najściślejszą kontrolę nad miejscem, w którym konstrukcja się osadza, oraz najdokładniejsze ustawienie genomowe, chociaż wybór klonów jest wolniejszy, a przepustowość niższa.
| Metoda integracji | Kontrola wstawiania | Stabilność | Skalowalność | Rozważania regulacyjne |
|---|---|---|---|---|
| Dostarczenie lentiwirusowe | Niskie (losowa integracja) | Wysoka, ale podatna na wyciszanie | Wysoka | Większa kontrola regulacyjna z powodu losowego wstawiania i pozostałości wirusowych |
| Transpozony (PiggyBac/SB) | Umiarkowane | Wysoka przez wiele pasaży | Wysoka | Wymaga badania liczby kopii i miejsc wstawienia |
| CRISPR Knock-in | Wysokie (specyficzne dla miejsca) | Bardzo wysoka | Umiarkowana | Korzystne; zmniejsza ryzyko zakłócenia genów endogennych |
| Wektory episomalne | Brak (pozachromosomalne) | Niskie; mogą zostać utracone podczas podziału | Niskie | Niższa przeszkoda integracyjna, ale nieodpowiednie do długoterminowej ekspansji |
Przesiewanie klonów musi zrobić więcej niż tylko potwierdzić obecność konstruktu.Powinno śledzić dryf ekspresji, profil insercji, kinetykę wzrostu, wydajność różnicowania i zachowanie fenotypu przy wysokiej liczbie pasaży. snRNA-seq jest tutaj przydatne, ponieważ może wykluczyć klony wzbogacone w komórki rezerwowe Pax7⁺/Ki-67⁻ - komórki, które opuszczają cykl komórkowy bez różnicowania - przed skalowaniem [3]. Wieprzowe EPSC z obwodem Tet-On-PAX7 zachowały wysokie różnicowanie mięśni w 3D na mikronośnikach i w kulturze zawiesinowej przez ponad 40 pasaży [8].
4.2 Jak zmienia się zachowanie obwodu w kulturze 3D, na mikronośnikach i w bioreaktorze
Gdy masz klon, następnym testem jest sprawdzenie, czy zachowuje się tak samo poza 2D. W wielu przypadkach tak nie jest. Wydajność w 2D rzadko przenosi się czysto na zawiesinę, mikronośniki lub kulturę na rusztowaniu, ponieważ gradienty dyfuzji, ograniczenia tlenu i ścinanie zmieniają wydajność obwodu.
Jednym z pierwszych testów jest dyfuzja induktora. W bioreaktorach z mieszadłem, małocząsteczkowe induktory muszą równomiernie docierać do komórek. W praktyce mogą tworzyć się gradienty, zwłaszcza w gęstych kulturach na mikronośnikach oraz wewnątrz agregatów lub rdzeni rusztowań. Hodowla w zawiesinie zazwyczaj lepiej nadaje się do produkcji mięsa hodowanego na dużą skalę, ponieważ wspiera wyższą gęstość komórek i zapewnia lepszą kontrolę procesu.
Monitorowanie stanu komórek staje się również trudniejsze wraz ze skalowaniem systemu. Sygnały reporterów fluorescencyjnych, które są łatwe do odczytania za pomocą mikroskopii w 2D, mogą być trudne do rozróżnienia w nieprzezroczystych konstrukcjach 3D. Fluorescencyjne timery - sondy, które zmieniają emisję z niebieskiej na czerwoną w miarę dojrzewania białka - mogą dostarczać danych o aktywacji obwodów w czasie rzeczywistym in situ [1]. Rozsądna droga to etapowa walidacja: najpierw w 2D, potem w konstrukcjach 3D, a następnie w końcowych warunkach bioreaktora [3][8].
4.3 Charakterystyka, dokumentacja bezpieczeństwa żywności i rozważania dotyczące UK/UE
Po testach procesowych, charakterystyka musi wykazać, że funkcja obwodu, fenotyp i bezpieczeństwo nadal są zachowane. Podstawowy zestaw danych powinien obejmować cytometrię przepływową, qPCR z sekwencjami cm, kursy czasowe RNA-seq oraz funkcjonalne odczyty, takie jak frakcja powierzchni łańcucha ciężkiego miozyny i ekspresja mioglobiny [1]. Optymalizowane media różnicujące bez surowicy wykazały, że ekspresja mioglobiny osiąga około 30% poziomów występujących w natywnych mięśniach bydła [3]. To daje zespołom jasny punkt odniesienia, a nie nieokreślony cel.
Musisz również udokumentować profile białek, aminokwasów i tłuszczów, wraz z cechami sensorycznymi [10][3].
Z punktu widzenia regulacji, Wielka Brytania i UE wyraźnie rozróżniają linie komórkowe spontanicznie unieśmiertelnione (nie-GMO) od linii genetycznie modyfikowanych. Te ostatnie wymagają szerszych dossier bezpieczeństwa [10][3]. Pakiety stabilności powinny wykazywać zachowanie fenotypu i stabilność genomową w całym procesie produkcji - od głównego banku komórek do końcowych komórek produkcyjnych - a zapisy śledzenia muszą uwzględniać każdy etap pośredni [10]. Jeśli obwód zależy od chemicznego induktora, preferowany jest dodatek bezpieczny dla żywności lub licencjonowany, taki jak kwas wanilinowy, zamiast doksycykliny [1].
Rutynowy nadzór genomowy jest koniecznością, a indukowalny przełącznik samobójczy lub eliminacyjny powinien być udokumentowany jako podstawowy środek kontroli ryzyka [7]. Jego funkcja powinna być również uwzględniona w dokumentacji bezpieczeństwa, zwłaszcza że przepisy dotyczące mięsa hodowlanego w Wielkiej Brytanii i UE nadal się kształtują.
5. Praktyczna mapa drogowa i wnioski
5.1 Stopniowa mapa drogowa wdrożenia dla zespołów zajmujących się mięsem hodowlanym
Najczystsza droga od koncepcji do produkcji to etapowy przepływ pracy.
Faza 1 to projektowanie. Zacznij od zdefiniowania docelowej linii, a następnie użyj snRNA-seq, aby potwierdzić główne wąskie gardła, zanim wybierzesz architekturę obwodu. Ten krok ma znaczenie, ponieważ obwód może rozwiązać tylko te ograniczenia, które faktycznie zidentyfikowałeś.
Faza 2 to budowa i walidacja 2D. Zbuduj konstrukcję i sprawdź, czy obwód zachowuje się zgodnie z oczekiwaniami w 2D, używając prostego odczytu raportera.Na tym etapie cel jest prosty: potwierdzić, że logika działa, zanim przejdziemy do trudniejszych i bardziej kosztownych modeli.
Faza 3 to testowanie wytrzymałościowe na skalę. Przejście do systemów 3D i warunków odpowiednich dla bioreaktorów, a następnie porównanie wyników z bazą 2D. To tutaj wiele projektów zaczyna ujawniać swoje słabe punkty, zwłaszcza gdy w grę wchodzą transfer masy, ścinanie i efekty matrycowe.
Faza 4 to integracja regulacyjna i bezpieczeństwa, i powinna być prowadzona równolegle z Fazą 3. Prace nad bezpieczeństwem i regulacjami nie powinny czekać do końca. Należy je prowadzić równolegle z rozwojem skali, w tym dokumentację dla wszelkich indukowalnych modułów bezpieczeństwa.
5.2 Pozyskiwanie narzędzi i materiałów umożliwiających przez Cellbase
Gdy przepływ pracy jest ustalony, pozyskiwanie często staje się krokiem ograniczającym tempo.
- linie komórkowe
- media bezsurowicze i chemicznie zdefiniowane
- rusztowania
- komponenty bioreaktorów
- czujniki
- sprzęt analityczny
Niezawodny dostęp do kompatybilnych materiałów na każdym etapie ma bezpośredni wpływ na to, jak szybko można scharakteryzować zachowanie obwodów w warunkach istotnych dla skali.
5.3 Kluczowe wnioski
Syntetyczne obwody genowe dają zespołom zajmującym się mięsem hodowlanym programowalną kontrolę nad czasem, progami i równowagą linii, której protokoły oparte wyłącznie na mediach nie mogą dorównać. Wybór architektury kształtuje odwracalność, przecieki i bezpieczeństwo.Systemy indukowane są zazwyczaj preferowane, ponieważ zapewniają warunkową kontrolę i niższe obciążenie metaboliczne [6].
"Zestaw narzędzi biologii syntetycznej może być użyty do ustanowienia linii komórkowych z regulowaną ekspresją genów, które, w połączeniu z PAT i modelowaniem komputerowym, mogą umożliwić systemy sterowania w pętli zamkniętej do dostarczania optymalnej wydajności i jakości produktu." - npj Systems Biology and Applications [6]
Udane wdrożenie nie jest tylko problemem biologicznym. Zależy od ścisłego powiązania między inżynierią obwodów, projektowaniem bioprocesów, dokumentacją regulacyjną i zaopatrzeniem.
FAQs
Jak syntetyczne obwody genowe poprawiają spójność różnicowania?
Syntetyczne obwody genowe mogą uczynić różnicowanie bardziej spójnym, ponieważ dają zaprogramowaną kontrolę nad zachowaniem komórek i zaangażowaniem linii.W praktyce oznacza to wykorzystanie modułowych operacji logicznych do regulacji ekspresji genów i czynników transkrypcyjnych z precyzyjnym
timingiemTen timing ma znaczenie. Pomaga komórkom przechodzić przez zdefiniowane zmiany stanów we właściwej kolejności, zamiast dryfować w mieszane lub niepożądane losy. Zmniejsza również różnicowanie poza celem i redukuje szum w całej kulturze.
Korzyść jest prosta: bardziej jednolite, stabilne i dojrzałe populacje komórek do produkcji mięsa hodowlanego.
Jaki projekt obwodu pasuje do kontroli miogenicznej lub adipogenicznej?
W badaniach nad mięsem hodowlanym te same fibroblasty kurczaka mogą być skierowane do dowolnej linii. Miogeneza podąża za jednym zestawem protokołów indukcji, podczas gdy adipogeneza może być włączona poprzez wystawienie komórek na działanie takich czynników jak surowica z kurczaka lub kwasy tłuszczowe.
Stamtąd losy komórek można kontrolować krok po kroku wewnątrz trójwymiarowych rusztowań hydrożelowych, aby budować struktury mięsa z określonymi proporcjami tłuszczu i kolagenu.
Dlaczego obwody genowe często zachowują się inaczej w kulturze 3D?
W kulturze 3D , obwody genowe często zachowują się inaczej, ponieważ komórki mają do czynienia z fizycznymi i strukturalnymi bodźcami, które po prostu nie istnieją w dwuwymiarowych monowarstwowych. Te bodźce obejmują napięcie mechaniczne, naprężenie ścinające, sztywność macierzy i lokalną gęstość komórek.
Te sygnały mogą zmieniać szlaki sygnalizacyjne, takie jak Notch. Mogą również zmieniać sposób, w jaki syntetyczne obwody wykrywają siłę i koordynują odpowiedzi w dół strumienia, w tym adhezję komórka-komórka i morfogenezę tkanek.









