

Si vous pouvez étendre les cellules mais ne pouvez pas les orienter vers le bon destin au bon moment, votre processus s'arrêtera à la différenciation. C'est le point central ici : les circuits génétiques synthétiques vous donnent un contrôle intra-cellulaire sur l'engagement, le timing, la mémoire et le mélange de lignées, là où les changements de milieu laissent souvent des populations hétérogènes, partiellement engagées.
Si je devais construire un flux de travail de différenciation de viande cultivée, je tirerais quatre points de cet article immédiatement :
- Commencez avec le réseau natif, pas la construction. Utilisez l'analyse de trajectoire snRNA-seq, , l'inférence GRN et le profilage miRNA pour identifier où les cellules s'arrêtent, dérivent ou se dirigent vers le mauvais destin.
-
Associez le type de circuit au problème du processus.
Un interrupteur à bascule convient au verrouillage, un design feedforward ou passe-bande convient au contrôle du timing, une porte logique convient à la gestion de signaux multiples, et les miSFITs conviennent à une sortie graduée. - Concevoir pour une faible fuite, un faible bruit et la sécurité dès le premier jour. Les pièces orthogonales, l'autorégulation négative, les iFFLs, les transgènes cm et un module d'arrêt de croissance ou de mise à mort inductible font partie de la construction, pas une réflexion après coup.
- Valider dans des conditions pertinentes à l'échelle dès le début. Un circuit qui fonctionne en 2D peut changer en 3D, microporteur ou suspension agitée en raison des gradients d'inducteur, des limites d'oxygène et du cisaillement.
L'article souligne également un point pratique qui importe pour les équipes de processus : le contrôle de lignée unique et le contrôle de ratio sont des tâches différentes. Une cassette Tet-On MyoD peut favoriser l'entrée myogénique, mais les produits découpés nécessitent le contrôle des proportions de muscle, de graisse et de MEC, ce qui signifie généralement rétroaction, signalisation paracrine et un dépistage plus rigoureux des clones.
Quelques chiffres ancrent ce message :
- La différenciation myogénique standard peut s'arrêter avec des indices de fusion d'environ 50–60%
- Les GRNs conçus dans les iPSCs ont augmenté la différenciation de la lignée cible de 52% à 81%
- Les circuits synthétiques dans les MSCs modifiés ont conduit la différenciation cardiaque à 76%
- Certaines lignées porcine Tet-On-PAX7 ont conservé un potentiel myogénique élevé au-delà de 40 passages
- Environ 20% des cellules souches pluripotentes humaines peuvent porter des mutations liées au cancer, c'est pourquoi les modules de sécurité inductibles sont importants
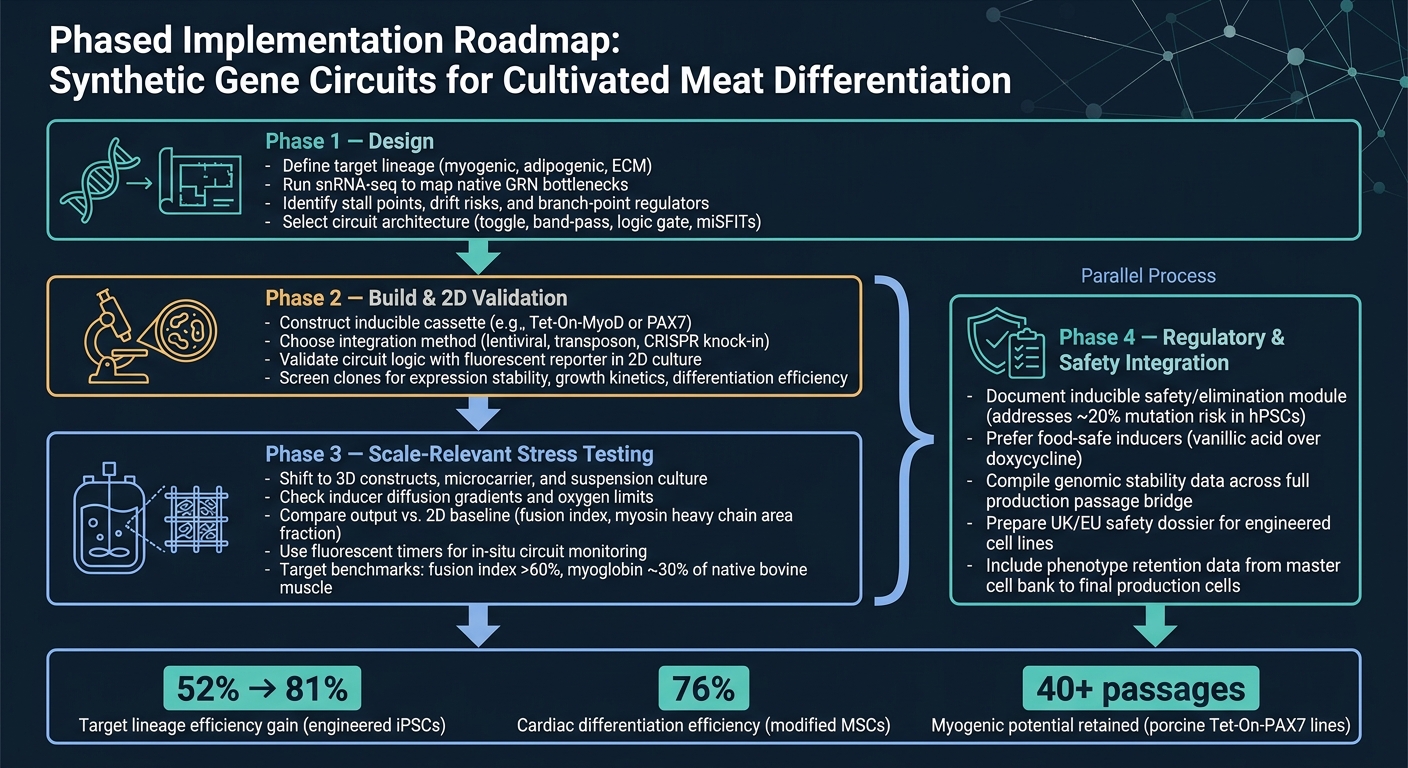
Flux de travail des circuits génétiques synthétiques pour la différenciation de la viande cultivée
Histoires de scientifiques : Michael Elowitz, Circuits génétiques dans les cellules vivantes
Comparaison rapide
| Sujet | Ce que l'article dit en termes simples |
|---|---|
| Goulot d'étranglement principal | Diversification, pas expansion |
| Pourquoi les circuits aident | Ils ajoutent un contrôle de seuil et une mémoire d'état cellulaire à l'intérieur de la cellule |
| Meilleur premier pas | Cartographier le contrôle du destin natif avec snRNA-seq et outils connexes |
| Principales options de circuit | Bascule, feedforward, passe-bande, portes logiques, miSFITs, CRISPRa/i |
| Exemple de myogenèse | Tet-On-MyoD pour séparer la croissance de la différenciation terminale |
| Adipogenèse / Contrôle de la MEC | miSFITs et conceptions en nœud papillon pour une sortie graduée de type PPARG/BMP4 |
| Défi de la coupe entière | Contrôle du ratio entre muscle, graisse et tissu conjonctif |
| Risque d'échelle | Le comportement en 2D peut ne pas se maintenir en 3D ou dans des bioréacteurs |
| Choix d'intégration | Lentivirus, transposons, CRISPR knock-in, vecteurs épisomaux |
| Point réglementaire | Les lignées modifiées nécessitent un ensemble de sécurité plus large ; les inducteurs sûrs pour l'alimentation tels que l'acide vanillique sont préférés à la DOX lorsque cela est possible |
Donc, en termes simples : ceci n'est pas seulement un article de conception de circuit. Je le lirais comme un guide pour lier l'architecture des circuits, la biologie de la lignée, la sélection des clones, la performance des bioréacteurs et la documentation de sécurité UK/UE en une stratégie de différenciation.
Lisez la suite si vous voulez le chemin complet de la cartographie du réseau natif au choix de la construction, aux vérifications de montée en échelle et à l'adéquation réglementaire.
2. Principes de conception pour les circuits de contrôle de la différenciation
2.1 Cartographier le réseau de destin cellulaire natif avant de concevoir le circuit
Avant de concevoir un circuit, vous avez besoin d'une image claire de ce que la cellule fait déjà.
Le séquençage de l'ARN à noyau unique (snRNA-seq) est un bon point de départ. Il peut montrer des sous-populations quiescentes, y compris des cellules de réserve marquées par NOTCH2 et HEYL , et indiquer des cibles de voies qui peuvent améliorer la différenciation [3].
À partir de là, l'analyse de trajectoire et l'inférence du réseau de régulation génique (GRN) aident à cartographier l'ordre d'activation des régulateurs et à mettre en évidence où les cellules sont les plus susceptibles de stagner. Dans la myogenèse, la cascade principale passe par MYOD1 et MYOG . Dans l'adipogenèse, les principaux nœuds sont PPARG et CEBPA , avec le destin des progéniteurs fibro-adipogéniques (FAP) agissant comme le principal point de risque de bifurcation. Le tableau ci-dessous résume les principaux régulateurs et goulots d'étranglement.
| Lignée | Régulateurs maîtres clés | Voies de signalisation critiques | Goulots d'étranglement identifiés |
|---|---|---|---|
| Myogénique | MYOD1, MYOG, PAX7 | MEK/ERK, NOTCH, WNT | Formation de cellules de réserve (quiescence) |
| Adipogénique | PPARG, CEBPA, ZFP423 | RXR, TGF-β, BMP | Destin des progéniteurs fibro-adipogéniques (FAP) |
| Pluripotent | OCT4, SOX2, NANOG | FGF, TGF-β/Nodal | Différenciation spontanée / hétérogénéité |
Une autre couche utile est le profilage de l'expression des miARN. Les miARN endogènes tels que miR-302a, liés à la pluripotence, et miR-375, liés à la différenciation, peuvent agir comme des classificateurs internes dans les conceptions de type sense-and-respond. Cela permet au circuit de lire l'état réel de la cellule au lieu de dépendre uniquement des inducteurs externes [5].
Ces goulets d'étranglement devraient guider le choix du circuit. Si le principal problème est la dérive, vous pourriez avoir besoin d'un verrouillage. Si le problème est le timing, une impulsion peut mieux convenir. Si le contrôle du destin dépend de plus d'un signal, une logique multi-entrée a généralement plus de sens.
2.2 Choisissez la bonne architecture de circuit
C'est là que les compromis apparaissent clairement. La bonne architecture dépend de trois points pratiques : à quel point l'engagement doit être permanent, à quel point le timing doit être contrôlé et combien de charge génétique la cellule peut transporter sans problème.
Les bascules bistables conviennent aux cas où l'engagement de la lignée doit rester verrouillé. Le principal problème est le basculement spontané de l'état causé par le bruit transcriptionnel.
Les filtres passe-bande conviennent aux cas où un facteur de transcription est nécessaire uniquement dans une fenêtre de développement définie. Le problème est que les niveaux d'inducteur nécessitent un contrôle strict, sinon le timing dérape.
Les portes logiques ajoutent de la spécificité en nécessitant plus d'une entrée en même temps. Par exemple, la différenciation peut être autorisée uniquement lorsqu'un inducteur exogène est présent et que la cellule présente le bon profil de miARN endogène. Cela aide à réduire le risque d'engagement hors cible.
Le tableau ci-dessous présente les principales architectures et les compromis qui les accompagnent.
| Architecture | Réversibilité | Précision temporelle | Complexité d'intégration | Cas d'utilisation principal | Risque clé |
|---|---|---|---|---|---|
| Interrupteur bistable | Faible (une fois verrouillé) | Modéré | Modéré | Engagement de lignée permanent | Renversement spontané dû au bruit |
| Filtre passe-bande | Élevée (dépendant de la concentration) | Élevée | Élevée | Étapes de développement transitoires | Nécessite un contrôle strict de l'inducteur |
| Porte logique (ET/OU/PAS) | Variable | Modéré | Modéré–élevé | Activation spécifique au type cellulaire | Fuites dans les états OFF |
| Nœud papillon / multi-entrée | Élevé | Modéré | Modéré | Intégration multi-signal | Se base sur la stabilité des miRNA endogènes |
| miSFITs | Élevé | Modéré | Faible–modéré | Ajustement de sortie gradué | Plage dynamique étroite si mal ajustée |
"En minimisant le nombre de couches computationnelles tout en maintenant la fonctionnalité, cette stratégie aborde les obstacles de scalabilité dans l'ingénierie des circuits génétiques." - Nature Communications [9]
Chaque couche réglementaire ajoutée augmente la charge utile et puise dans les ressources cellulaires. En pratique, si deux conceptions font le même travail, la plus simple est généralement la meilleure option lorsque l'échelle compte.
Une fois l'architecture définie, le travail suivant consiste à la faire tenir sous faible fuite, suppression du bruit et contrôle de sécurité.
2.3 Construire pour la fiabilité, la faible fuite et la sécurité
Un circuit doit rester stable sur une culture prolongée. Une courte période de bonne performance n'est pas suffisante pour une utilisation en production.
Les pièces orthogonales sont la première défense. Les promoteurs, facteurs de transcription et éléments régulateurs qui ne réagissent pas avec la machinerie native aident à limiter les effets hors cible et réduisent la chance que des signaux endogènes activent ou désactivent le circuit. Des promoteurs modifiés à haute étanchéité tels que PCREm ont été utilisés pour réduire les fuites basales dans les systèmes mammaliens inductibles [6] .
L'autorégulation négative mérite également d'être ajoutée lorsque cela est possible. C'est l'un des motifs les plus connus pour réduire le bruit transcriptionnel et produire une réponse plus linéaire à la concentration d'inducteur [6]. Les boucles d'alimentation incohérentes (iFFLs) peuvent ajouter une autre couche de contrôle en filtrant les fluctuations stochastiques, de sorte que les cellules répondent aux signaux soutenus plutôt qu'aux pics de bruit courts.
Les versions à codons modifiés (cm ) des facteurs de transcription synthétiques facilitent également la caractérisation. Elles permettent de séparer l'expression pilotée par le circuit de l'expression génomique endogène (g) lors de la validation [1]. Cela peut sembler être un petit détail, mais cela permet de gagner du temps lorsque vous essayez de déterminer si une lecture provient du circuit ou du génome hôte.
Les modules de sécurité sont requis. Environ 20 % des cellules souches pluripotentes humaines portent des mutations associées au cancer [7]. Donc, si un circuit est intégré dans une lignée dérivée de cellules souches, il devrait inclure un module d'arrêt de croissance ou d'élimination inductible. L'acide vanillique est un inducteur utile à privilégier ici car c'est un additif alimentaire autorisé, ce qui renforce l'argument pour l'utiliser comme déclencheur de circuit dans les lignées cellulaires de viande cultivée [1].
"La biologie synthétique fournit aux ingénieurs les moyens de déployer des circuits pour ajuster facilement et précisément l'expression de plusieurs gènes afin de... éliminer les effets secondaires potentiels hors cible." - npj Systems Biology and Applications [6]
Ces choix établissent les circuits spécifiques à la lignée dans la Section 3.
sbb-itb-ffee270
3. Stratégies de circuits pour la différenciation myogénique, adipogénique et contrôlée par ratio
3.1 Circuits myogéniques qui séparent l'expansion de la différenciation terminale
Une fois l'architecture du circuit établie, la prochaine tâche est le déploiement spécifique à la lignée. Pour la myogenèse, le principal problème est simple à énoncer mais difficile à exécuter : les cellules doivent d'abord proliférer, puis passer à la différenciation sur demande, sans dériver trop tôt dans l'une ou l'autre direction.
Une cassette Tet-On-MyoD est l'une des façons les plus directes de le faire. Dans cette configuration, les cellules prolifèrent dans des conditions standard lorsque la doxycycline (DOX) est absente. Ajoutez du DOX, et le circuit entraîne l'engagement myogénique.Des chercheurs de plusieurs institutions chinoises ont utilisé cette approche dans des fibroblastes embryonnaires de poulet et ont rapporté une formation efficace de myotubes après induction [4].
Un interrupteur bistable offre un contrôle d'état plus strict. Les systèmes construits à partir de répresseurs mutuellement inhibiteurs tels que E-KRAB et Pip-KRAB peuvent transformer une courte impulsion de DOX en un programme myogénique stable [6]. En pratique, cela signifie que l'état différencié reste verrouillé après induction plutôt que de s'estomper une fois le signal retiré. L'ajout d'une autorégulation négative aide également à réduire le bruit stochastique et à limiter la différenciation fuyante pendant la phase d'expansion [6].
Cela est important car la différenciation myogénique standard in vitro s'arrête souvent à mi-chemin. Les indices de fusion sont généralement seulement autour de 50–60%, ce qui laisse une grande population de réserve non fusionnante [3]. L'activation de MyoD par circuit peut améliorer l'engagement, mais l'engagement seul ne garantit pas une fusion uniforme. Lorsque les circuits MyoD sont combinés avec la modulation de MEK, NOTCH et RXR, une fusion presque complète en culture 2D devient possible [3]. Le circuit contrôle le timing de l'entrée de la lignée ; les petites molécules favorisent une sortie du cycle cellulaire plus propre et plus uniforme.
3.2 Contrôle adipogénique et fibrogénique pour la composition et la structure
Le muscle seul ne suffit pas. Une fois le contrôle myogénique en place, le problème suivant est la composition : combien de graisse se forme, combien de MEC est déposée, et quand ces programmes s'activent. Ici, le contrôle binaire ON/OFF est souvent trop brutal. Ce dont les équipes ont généralement besoin, c'est d'une sortie graduée, surtout autour du point de branchement adipogénique-fibrogénique.
Les miSFITs offrent un moyen pratique de régler l'expression par étapes. En plaçant des sites cibles de miARN mutés - par exemple, des sites pour miR-17 - dans la 3′UTR de gènes de sortie tels que PPARG ou BMP4, les chercheurs peuvent sélectionner des niveaux d'expression à partir d'une bibliothèque de variantes [5]. Cela rend l'induction des adipocytes plus semblable à un variateur qu'à un interrupteur. Au lieu de pousser les cellules dans une réponse tout ou rien, les équipes peuvent titrer l'adipogenèse plus soigneusement [5].
Les fibroblastes ne sont pas non plus de simples spectateurs ici. Ils fournissent des protéines ECM qui façonnent la texture [10]. Cela fait du contrôle fibrogénique une partie de la conception du produit, et non juste un problème secondaire. Les circuits peuvent aider à gérer la transition entre les états fibrogénique et adipogénique, et dans la volaille cultivée, l'activation directe de PPARG dans les fibroblastes peut être nécessaire pour générer un dépôt de graisse significatif [10].
Une architecture en nœud papillon convient bien à ce problème car elle maintient la détection et la sortie séparées. La couche de détection lit l'état actuel de la cellule, tandis que la couche de sortie ajuste PPARG, CEBPA ou d'autres régulateurs de lignée. Cette séparation aide à empêcher les programmes adipogéniques ou de tissu conjonctif de s'activer avant que les cellules n'aient atteint le bon stade de développement.
3.3 Contrôle du ratio multi-lignée et détection par rétroaction
Les circuits de contrôle du ratio traitent d'un problème différent : non pas si la différenciation se produit, mais si le mélange final de la population reste là où il devrait être. Pour les produits entiers, obtenir les bonnes proportions de muscle, de graisse et de MEC est tout aussi important que de faire différencier une lignée quelconque.
Ces systèmes intègrent le contrôle par rétroaction dans les cellules elles-mêmes. Les promoteurs spécifiques à l'état restreignent l'expression des protéines de signalisation aux cellules qui se sont déjà engagées dans une lignée donnée.Les modules paracrines synthétiques permettent ensuite aux cellules myogéniques engagées de libérer un signal inhibiteur qui supprime l'engagement adipogénique dans les cellules voisines. La logique ressemble à l'inhibition latérale dans les systèmes de développement Delta-Notch [1][6] . Lorsque la ramification devient plus compliquée, des portes multi-entrées peuvent combiner des signaux extracellulaires avec des signaux d'état interne [9].
Les miSFITs fonctionnent également à ce stade. En ajustant la force de sortie de BMP4 ou d'autres morphogènes, les équipes peuvent modifier l'équilibre des lignées sans réécrire la logique de décision en amont. Dans les réseaux de régulation génique conçus, ce type de contrôle a augmenté l'efficacité de différenciation de la lignée cible de 52 % dans les contrôles à 81 % dans les iPSCs conçus [2]. Dans les cellules souches mésenchymateuses modifiées, les circuits synthétiques ont conduit l'efficacité de différenciation cardiaque à 76 % [2].
Le tableau ci-dessous compare les approches à lignée unique et à contrôle de ratio en utilisant les points qui comptent le plus en production.
| Caractéristique | Circuit à lignée unique | Circuit de contrôle de ratio |
|---|---|---|
| Complexité | Basse; typiquement un seul promoteur inductible et régulateur [4] | Élevée; nécessite des portes logiques et une signalisation paracrine [6][9] |
| Charge de surveillance | Basse; suit généralement un seul rapporteur [4] | Élevée; nécessite le suivi de plusieurs marqueurs de lignée [5] |
| Robustesse | Modérée; sujette à l'hétérogénéité et à la formation de cellules de réserve [3] | Élevé ; utilise la rétroaction et l'inhibition latérale pour équilibrer les populations [1] |
| Valeur de production | Élevé pour la biomasse ; limité pour la structure tissulaire complexe [10] | Essentiel pour les produits entiers nécessitant muscle, graisse et ECM [4] |
Les circuits de contrôle du ratio ajoutent une charge de validation plus lourde. Mais leur retour d'information intégré est difficile à égaler avec le seul contrôle de processus, ce qui met plus de pression sur la sélection des clones et les tests de processus.
4. Du concept au processus : validation, montée en échelle et adéquation réglementaire
4.1 Stratégie d'intégration et sélection des clones pour une performance stable
Après la conception du circuit, la partie difficile commence : intégrer ce circuit dans les cellules de manière à ce qu'il reste stable à travers le passage à l'échelle de production.
La livraison lentivirale est souvent efficace et donne rapidement des intégrants stables. Mais l'intégration est aléatoire. Cela signifie plus d'attention réglementaire, ainsi qu'un risque de diminution de l'expression au fil du temps en raison de la silenciation. Les systèmes de transposons tels que PiggyBac et Sleeping Beauty se situent au milieu. Ils peuvent maintenir la performance à travers de nombreux passages, mais il est toujours nécessaire de vérifier le nombre de copies et les sites d'insertion. Sleeping Beauty , par exemple, a été utilisé pour immortaliser de manière stable les cellules satellites bovines en surexprimant TERT et CDK4, avec des lignées conservant leur potentiel myogénique au-delà de 40 passages [10] . CRISPR knock-in offre le contrôle le plus strict sur l'emplacement du construct et le cadre génomique le plus précis, bien que la sélection des clones soit plus lente et que le débit soit plus faible.
| Méthode d'intégration | Contrôle d'insertion | Stabilité | Évolutivité | Considérations réglementaires |
|---|---|---|---|---|
| Livraison lentivirale | Faible (intégration aléatoire) | Élevée, mais sujette au silence | Élevée | Examen réglementaire accru en raison de l'insertion aléatoire et des résidus viraux |
| Transposons (PiggyBac/SB) | Modérée | Élevée sur de nombreux passages | Élevée | Nécessite un dépistage du nombre de copies et des sites d'insertion |
| Knock-in CRISPR | Élevée (site spécifique) | Très élevée | Modérée | Favorable; réduit le risque de perturber les gènes endogènes |
| Vecteurs épisomaux | Aucun (extrachromosomique) | Faible ; peut être perdu lors de la division | Faible | Obstacle d'intégration plus faible, mais inadapté pour une expansion à long terme |
Le dépistage des clones doit faire plus que confirmer la présence du construct.Il devrait suivre la dérive d'expression, le profil d'insertion, la cinétique de croissance, l'efficacité de différenciation et la rétention du phénotype à un nombre de passages élevé. La snRNA-seq est utile ici car elle peut exclure les clones enrichis en cellules de réserve Pax7⁺/Ki-67⁻ - cellules qui quittent le cycle cellulaire sans se différencier - avant la montée en échelle [3]. Les EPSCs porcins avec un circuit Tet-On-PAX7 ont conservé une différenciation musculaire élevée en culture sur microporteur 3D et en suspension sur plus de 40 passages [8].
4.2 Comment le comportement du circuit change en culture 3D, sur microporteur et en bioréacteur
Une fois que vous avez un clone, le test suivant est de savoir s'il se comporte de la même manière en dehors du 2D. Dans de nombreux cas, ce n'est pas le cas. La performance en 2D se transpose rarement proprement à la suspension, au microporteur ou à la culture sur échafaudage car les gradients de diffusion, les limites d'oxygène et le cisaillement modifient tous la sortie du circuit.
L'un des premiers contrôles est la diffusion de l'inducteur. Dans les bioréacteurs à cuve agitée, les inducteurs à petites molécules doivent atteindre les cellules de manière uniforme. En pratique, des gradients peuvent se former, surtout dans les cultures sur microporteurs denses et à l'intérieur des agrégats ou des noyaux de supports. La culture en suspension est généralement mieux adaptée pour la viande cultivée à grande échelle car elle soutient une densité cellulaire plus élevée et offre un contrôle de processus plus strict.
Le suivi de l'état des cellules devient également plus difficile à mesure que le système s'agrandit. Les signaux de rapporteurs fluorescents qui sont faciles à lire par microscopie en 2D peuvent devenir difficiles à résoudre dans des constructions 3D opaques. Les minuteurs fluorescents - des sondes qui changent d'émission du bleu au rouge à mesure qu'une protéine mûrit - peuvent fournir des données d'activation de circuit en temps réel in situ [1]. La voie raisonnable est la validation par étapes : d'abord en 2D, puis dans des constructions 3D, puis dans les conditions finales du bioréacteur [3][8] .
4.3 Caractérisation, documentation de sécurité alimentaire et considérations UK/UE
Après les tests de processus, la caractérisation doit montrer que la fonction du circuit, le phénotype et la sécurité sont toujours valides. Le noyau de données doit inclure la cytométrie en flux, la qPCR avec séquences cm, les cours de temps RNA-seq, et les lectures fonctionnelles telles que la fraction de la zone de la chaîne lourde de myosine et l'expression de la myoglobine [1]. Les milieux de différenciation optimisés sans sérum ont montré qu'ils pouvaient amener l'expression de la myoglobine à environ 30 % des niveaux trouvés dans le muscle bovin natif [3]. Cela donne aux équipes un point de référence clair plutôt qu'un objectif vague.
Vous devez également documenter les profils de protéines, d'acides aminés et de graisses, ainsi que les caractéristiques sensorielles [10][3].
D'un point de vue réglementaire, le Royaume-Uni et l'UE tracent une ligne claire entre les lignées cellulaires immortalisées spontanément (non-OGM) et les lignées génétiquement modifiées. Ces dernières nécessitent des dossiers de sécurité plus complets [10][3]. Les packages de stabilité doivent montrer la rétention du phénotype et la stabilité génomique tout au long du pont de production complet - de la banque de cellules maîtresses aux cellules de production finales - et les dossiers de traçabilité doivent rendre compte de chaque passage intermédiaire [10]. Si le circuit dépend d'un inducteur chimique, un additif sûr pour l'alimentation ou homologué tel que l'acide vanillique est préférable à la doxycycline [1].
La surveillance génomique de routine est indispensable, et un interrupteur d'élimination ou de suicide inductible devrait être documenté comme une mesure de contrôle des risques essentielle [7]. Sa fonction devrait également être incluse dans le dossier de sécurité, surtout alors que les règles du Royaume-Uni et de l'UE pour la viande cultivée continuent de se préciser.
5. Feuille de route pratique et conclusion
5.1 Une feuille de route de mise en œuvre par étapes pour les équipes de viande cultivée
La voie la plus directe du concept à la production est un flux de travail par étapes.
Phase 1 est la conception. Commencez par définir la lignée cible, puis utilisez le snRNA-seq pour confirmer les principaux goulots d'étranglement avant de choisir une architecture de circuit. Cette étape est importante car un circuit ne peut résoudre que les contraintes que vous avez réellement identifiées.
Phase 2 est la construction et la validation en 2D. Construisez le construct et vérifiez que le circuit se comporte comme prévu en 2D, en utilisant une lecture simple de rapporteur.À ce stade, l'objectif est simple : confirmer que la logique fonctionne avant de passer à des modèles plus complexes et coûteux.
Phase 3 est le test de résistance à l'échelle pertinente. Passez aux systèmes 3D et aux conditions pertinentes pour les bioréacteurs, puis comparez les résultats avec la base de référence 2D. C'est là que de nombreux designs commencent à montrer leurs points faibles, surtout lorsque le transfert de masse, le cisaillement et les effets de matrice entrent en jeu.
Phase 4 est l'intégration réglementaire et de sécurité, et elle doit se dérouler en parallèle avec la Phase 3. Le travail de sécurité et de réglementation ne doit pas attendre la fin. Exécutez-le parallèlement à la montée en échelle, y compris la documentation pour tout module de sécurité inductible.
5.2 Approvisionnement en outils et matériaux habilitants via Cellbase
Une fois le flux de travail établi, l'approvisionnement devient souvent l'étape limitante.
- lignes cellulaires
- milieux sans sérum et chimiquement définis
- échafaudages
- composants de bioréacteur
- capteurs
- équipements analytiques
Un accès fiable à des matériaux compatibles à chaque étape a un effet direct sur la rapidité avec laquelle le comportement des circuits peut être caractérisé dans des conditions pertinentes à l'échelle.
5.3 Points clés à retenir
Les circuits génétiques synthétiques offrent aux équipes de viande cultivée un contrôle programmable sur le timing, les seuils et l'équilibre des lignées que les protocoles basés uniquement sur les milieux ne peuvent égaler. Le choix de l'architecture façonne la réversibilité, la fuite et la sécurité.Les systèmes inductibles sont généralement préférés car ils offrent un contrôle conditionnel et une charge métabolique plus faible [6].
"La boîte à outils de biologie synthétique peut être utilisée pour établir des lignées cellulaires avec une expression génique modulable, qui, lorsqu'elle est couplée avec PAT et la modélisation computationnelle, peut permettre aux systèmes de contrôle en boucle fermée de fournir un rendement et une qualité de produit optimaux." - npj Systems Biology and Applications [6]
Le déploiement réussi n'est pas seulement un problème de biologie. Il dépend d'un couplage étroit entre l'ingénierie des circuits, la conception des bioprocédés, la documentation réglementaire et l'approvisionnement.
FAQs
Comment les circuits génétiques synthétiques améliorent-ils la cohérence de la différenciation?
Les circuits génétiques synthétiques peuvent rendre la différenciation plus cohérente car ils vous donnent un contrôle programmé sur le comportement cellulaire et l'engagement de la lignée.En pratique, cela signifie utiliser des opérations logiques modulaires pour ajuster l'expression des gènes et des facteurs de transcription avec un timing précis.
Ce timing est important. Il aide les cellules à traverser des changements d'état définis dans le bon ordre, au lieu de dériver vers des destins mixtes ou indésirables. Il réduit également la différenciation hors cible et diminue le bruit à travers la culture.
Le résultat est simple : des populations cellulaires plus uniformes, stables et matures pour la production de viande cultivée.
Quel design de circuit convient au contrôle myogénique ou adipogénique ?
Dans la recherche sur la viande cultivée, les mêmes fibroblastes de poulet peuvent être poussés dans l'une ou l'autre lignée. La myogenèse suit un ensemble de protocoles d'induction, tandis que l'adipogenèse peut être activée en exposant les cellules à des intrants tels que le sérum de poulet ou les acides gras.
À partir de là, ces destins cellulaires peuvent être contrôlés étape par étape à l'intérieur de structures en hydrogel 3D pour construire des structures de viande avec des ratios définis de graisse et de collagène.
Pourquoi les circuits génétiques se comportent-ils souvent différemment en culture 3D ?
En culture 3D, les circuits génétiques se comportent souvent différemment parce que les cellules traitent des entrées physiques et structurelles qui n'existent tout simplement pas dans les monocouches 2D. Ces entrées incluent la tension mécanique, le stress de cisaillement, la rigidité de la matrice et la densité cellulaire locale.
Ces signaux peuvent modifier les voies de signalisation telles que Notch. Ils peuvent également changer la façon dont les circuits synthétiques détectent la force et coordonnent les réponses en aval, y compris l'adhésion cellule-cellule et la morphogenèse tissulaire.









