

Pour les chercheurs en production de viande cultivée, minimiser l'apoptose est essentiel pour améliorer la viabilité cellulaire et la productivité dans les bioréacteurs. Les facteurs de stress tels que l'épuisement des nutriments, les déséquilibres osmotiques et l'accumulation de déchets déclenchent souvent la mort cellulaire, réduisant les rendements. Les gènes anti-apoptotiques peuvent atténuer ces défis en prolongeant la durée de vie des cellules pendant la culture. Voici un aperçu rapide des principaux gènes et de leurs rôles :
- BCL-2: Empêche la formation de pores mitochondriaux, bloquant l'apoptose à son initiation. Efficace pour les cellules non différenciées mais nécessite un équilibre soigneux avec les protéines pro-apoptotiques.
- BCL-xL: Protège les cellules pendant la différenciation et soutient le métabolisme énergétique. Idéal pour les phases de stress élevé dans les bioréacteurs.
- MCL-1: Offre une réponse rapide aux changements nutritionnels et reste stable pendant la différenciation. Fonctionne bien en combinaison avec d'autres gènes.
- BIRC5 (Survivin) : Inhibe les caspases pour bloquer l'apoptose en aval. Soutient la prolifération dans les cellules à division rapide.
- XIAP: Un puissant inhibiteur de caspases efficace dans des conditions de stress extrême, telles que les cultures à haute densité. La surveillance de ces conditions nécessite la sélection de capteurs pour les bioréacteurs de viande cultivée afin de suivre les niveaux de nutriments et l'accumulation de déchets en temps réel.
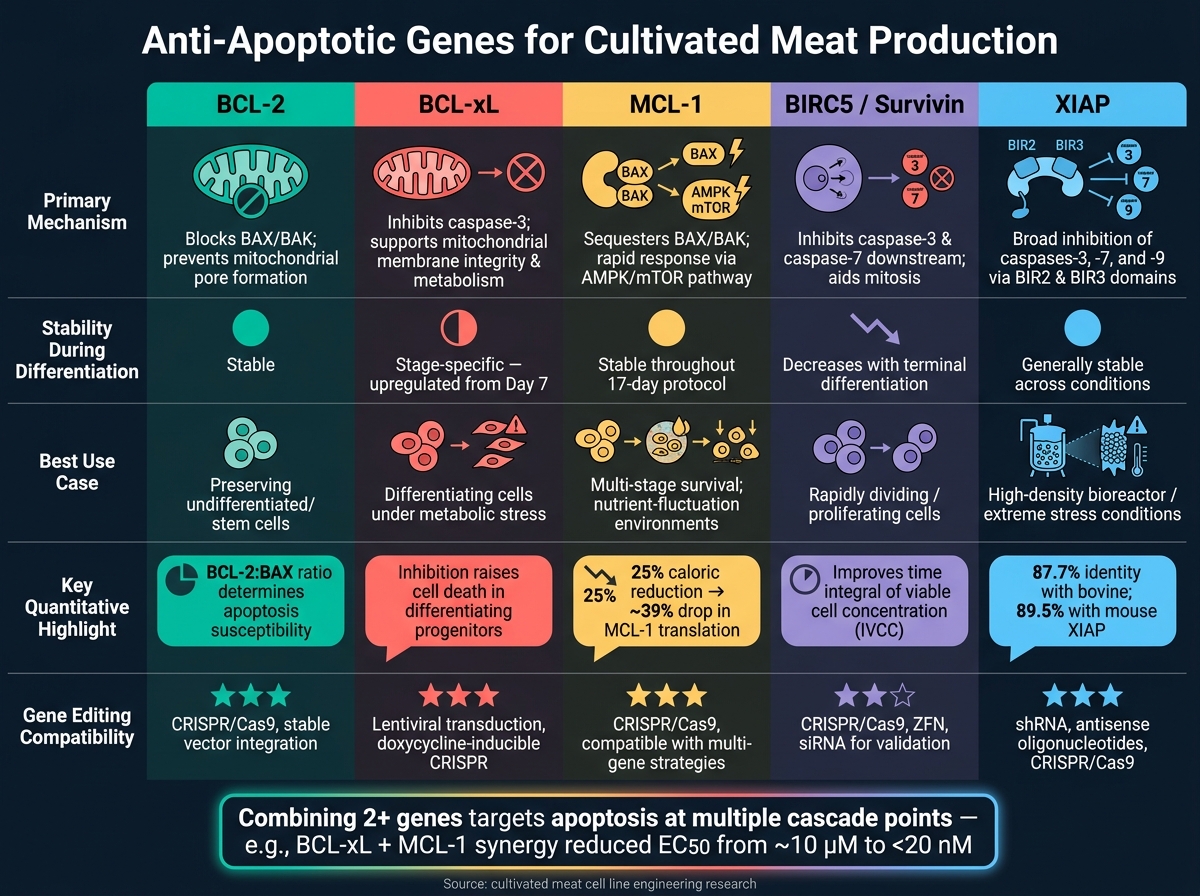
Comparaison rapide
| Gène | Rôle clé | Stabilité pendant la différenciation | Meilleur cas d'utilisation |
|---|---|---|---|
| BCL-2 | Bloque l'apoptose précoce (BAX/BAK) | Stable | Préservation des cellules non différenciées |
| BCL-xL | Prévient l'activation des caspases, soutient le métabolisme | Spécifique à l'étape | Cellules en différenciation sous stress |
| MCL-1 | Réponse rapide aux changements nutritionnels | Stable | Survie multi-étapes |
| BIRC5 | Inhibe les caspases en aval | Diminue avec la différenciation | Cellules à division rapide |
| XIAP | Inhibition large des caspases | Stable | Conditions de bioréacteur à haute tension |
1. BCL-2
BCL-2 est un gène anti-apoptotique bien étudié qui joue un rôle clé dans la voie intrinsèque (mitochondriale) de l'apoptose. Cette voie est un mécanisme majeur de mort cellulaire, souvent déclenché dans les cellules de viande cultivée sous les stress des bioréacteurs tels que les pénuries de nutriments ou les faibles niveaux d'oxygène.
BCL-2 fonctionne en se liant et en neutralisant les protéines pro-apoptotiques comme BAX et BAK. Cette action empêche la formation de pores mitochondriaux, stoppant la libération de cytochrome c et interrompant la cascade en aval de l'apoptose. Ce mécanisme est crucial pour prolonger la durée de vie viable des cellules dans la production de viande cultivée. Comme l'expliquent Rønning SB et al. :
"Le rapport entre Bcl-2 et Bax détermine la susceptibilité des cellules à subir l'apoptose."[5]
Au-delà de son rôle mitochondrial, BCL-2 réside également dans le réticulum endoplasmique (ER).Ici, il réduit les niveaux de calcium et inhibe la libération de calcium médiée par le récepteur IP3, atténuant l'apoptose induite par le calcium – un problème fréquent dans les cultures de bioréacteurs à haute densité[4]. La gestion de ces défis d'échelle est un objectif principal pour l'industrie. Cette double localisation permet à BCL-2 de protéger les cellules contre plusieurs déclencheurs d'apoptose.
La structure moléculaire de BCL-2, composée d'un faisceau de huit hélices alpha et de quatre domaines BH bien définis, en fait un e
Cependant, il y a un avertissement critique : l'équilibre entre BCL-2 et les protéines pro-apoptotiques comme BAX doit être soigneusement géré. Même des niveaux élevés d'expression de BCL-2 peuvent échouer à prévenir l'apoptose si les signaux pro-apoptotiques deviennent trop forts[2]. Surveiller cet équilibre est essentiel pour atteindre une viabilité cellulaire optimale.
sbb-itb-ffee270
2. BCL-xL
BCL-xL, codé par le gène BCL2L1, joue un rôle central dans la famille BCL-2 en se localisant à la membrane externe mitochondriale et en empêchant l'apoptose. Il y parvient en contrant les protéines pro-apoptotiques comme BAX et BAK.De plus, il inhibe la caspase-3 clivée (CASP3), qui est essentielle pour arrêter la mort cellulaire. Ce mécanisme est particulièrement précieux dans les cultures de bioréacteurs à haute densité , où le stress métabolique peut menacer la viabilité cellulaire.
Fait intéressant, l'activité de BCL-xL s'aligne avec des étapes spécifiques de la différenciation. Pendant certaines phases, son expression augmente, tandis que d'autres protéines anti-apoptotiques, telles que BCL-2 et MCL-1, restent inchangées. Cela souligne son importance dans le maintien de la survie cellulaire pendant la différenciation. Comme noté dans Cell Death & Disease:
"BCL-xL/BCL2L1 est une protéine anti-apoptotique critique qui favorise la survie des cellules en différenciation..." [2]
Au-delà de son rôle dans l'apoptose, BCL-xL soutient le métabolisme énergétique cellulaire. Il améliore à la fois la glycolyse et la phosphorylation oxydative, assurant une activité métabolique élevée.L'inhibition de BCL-xL a montré qu'elle réduit l'expression des gènes métaboliques et diminue à la fois la respiration mitochondriale basale et maximale. Cette fonction est particulièrement importante pour les cellules de viande cultivée, qui dépendent d'une production métabolique soutenue.
BCL-xL est hautement compatible avec les stratégies d'édition génétique couramment utilisées dans la recherche sur la viande cultivée. Des techniques comme la transduction lentivirale permettent une intégration stable du gène BCL2L1, tandis que les systèmes CRISPR/Cas9 induits par la doxycycline offrent un contrôle temporel précis de son expression [2] [6]. Ce niveau de précision est souvent géré par des logiciels avancés de contrôle des bioprocédés. Ces attributs font de BCL-xL un candidat solide pour améliorer la viabilité des lignées cellulaires dans la production de viande cultivée.
Pour les étapes de différenciation avec des exigences métaboliques élevées, BCL-xL peut être plus efficace que BCL-2.Les chercheurs peuvent utiliser l'inhibiteur WEHI-539 pour tester la dépendance d'une lignée cellulaire à BCL-xL avant de procéder à des modifications génétiques permanentes [2]. De plus, la co-expression de BCL-xL avec MCL-1 pourrait améliorer davantage la survie cellulaire, car il a été observé que ces protéines fonctionnent de manière synergique dans certains types de cellules résistantes [6].
3. MCL-1
MCL-1 (Myeloid Cell Leukaemia-1) joue un rôle clé dans la régulation de la voie apoptotique intrinsèque. Située sur la membrane externe mitochondriale, elle empêche l'apoptose en se liant et en séquestrant les protéines pro-apoptotiques BAX et BAK, arrêtant leur oligomérisation et la perméabilisation membranaire subséquente. Cette action bloque la libération du cytochrome c, arrêtant la cascade apoptotique avant qu'elle n'atteigne la phase d'exécution [8] . De plus, MCL-1 se lie aux protéines BH3-only - telles que Bim, PUMA et NOXA - avec une haute affinité [8]. Comme BCL-2 et BCL-xL, MCL-1 est essentiel pour contrer les signaux apoptotiques, surtout pendant le stress du bioréacteur.
Une des caractéristiques uniques de MCL-1 est sa courte demi-vie, rendant son expression très réactive à la disponibilité des nutriments et aux signaux métaboliques, en particulier via la voie AMPK/mTOR. Les études indiquent qu'une réduction de 25 % de l'apport calorique peut diminuer la traduction de MCL-1 d'environ 39 % ± 10 % [7]. Cette sensibilité est particulièrement pertinente pour la production de viande cultivée, où les fluctuations dans la composition du milieu de croissance ou l'épuisement des nutriments lors des cultures en suspension à grande échelle (qui nécessitent une planification de l'échelle de production minutieuse) peuvent réduire significativement les niveaux de MCL-1. De telles réductions compromettent la viabilité cellulaire, sapant les améliorations de la IVCC (concentration cellulaire viable intégrale) obtenues grâce à des stratégies anti-apoptotiques. Pour atténuer cela, des formulations de milieux sans sérum qui soutiennent une activité mTORC1 robuste sont essentielles [7] .
Une autre caractéristique notable de MCL-1 est sa stabilité pendant la différenciation. Dans les modèles de progéniteurs pancréatiques, l'expression de MCL-1 est restée stable tout au long d'un protocole de différenciation de 17 jours, contrairement à BCL-xL, qui a montré une variation dépendante du stade [2]. Cette stabilité rend MCL-1 particulièrement avantageux pour les applications de viande cultivée, où les cellules doivent survivre à plusieurs étapes de maturation sans nécessiter d'interventions précisément chronométrées.&
Les outils d'édition génétique peuvent être utilisés pour modifier MCL-1, tout comme d'autres gènes anti-apoptotiques, en faisant une cible polyvalente pour l'ingénierie des lignées cellulaires.
Lorsqu'il est utilisé en combinaison avec d'autres gènes anti-apoptotiques, MCL-1 offre des avantages supplémentaires. Par exemple, l'association de MCL-1 avec BCL-xL a montré des effets synergiques - l'inhibition simultanée des deux protéines a réduit l'EC50 des médicaments de survie d'environ 10 μM à moins de 20 nM [6]. Cette approche peut améliorer considérablement la survie cellulaire pendant les phases de stress intense de la production de viande cultivée.
4. BIRC5 (Survivine)
BIRC5, souvent appelé Survivine, est un membre de la famille des protéines Inhibiteur de l'Apoptose (IAP) [2]. Contrairement aux protéines de la famille BCL-2, qui agissent au niveau de la membrane mitochondriale pour empêcher l'initiation de l'apoptose, BIRC5 opère plus en aval. Il bloque les caspases responsables de l'exécution de l'apoptose, servant effectivement de dernière ligne de défense contre la mort cellulaire programmée [10].
Dans les cultures en suspension, des facteurs de stress tels que l'épuisement des nutriments, l'accumulation de déchets métaboliques et le stress de cisaillement mécanique peuvent déclencher l'apoptose. En inhibant l'activité des caspases à ce stade avancé, la surexpression de BIRC5 aide à prolonger la viabilité et la productivité des cellules. Cela se traduit par une amélioration de l'intégrale temporelle de la concentration cellulaire viable - une métrique clé pour optimiser la performance des cultures cellulaires [9] . Eric Baek, un chercheur à KAIST, explique:
"Améliorer l'intégrale temporelle de la concentration cellulaire viable en surmontant la mort cellulaire, notamment l'apoptose, est l'une des stratégies les plus largement utilisées pour la production efficace de protéines thérapeutiques [et de cellules]." [9]
Cette intervention en aval a montré qu'elle améliore les rendements des bioréacteurs dans les lignées cellulaires de viande cultivée, y compris les cellules satellites porcines et les myoblastes bovins.
La stratégie la plus efficace implique l'ingénierie combinatoire, en associant BIRC5 à des protecteurs mitochondriaux comme BCL-2 ou BCL-xL. Le professeur Michael Betenbaugh de l'Université Johns Hopkins souligne cette approche :
"Les stratégies qui bloquent la mort cellulaire à plusieurs points le long de la cascade peuvent limiter l'amplification de ces signaux d'apoptose." [10]
En combinant l'inhibition des caspases par BIRC5 avec une protection mitochondriale en amont, les chercheurs peuvent établir une défense à plusieurs niveaux contre l'apoptose.
BIRC5 s'intègre également parfaitement dans les flux de travail d'édition génétique.CRISPR/Cas9 est la méthode principale pour créer des lignées cellulaires stables avec surexpression [9], bien que les nucléases à doigts de zinc offrent une alternative précise. L'ARNsi peut être utilisé pour la validation des voies avant de s'engager dans l'intégration génomique [9].
5. XIAP
XIAP (inhibiteur de l'apoptose lié à l'X) est reconnu comme l'inhibiteur de caspase le plus puissant au sein de la famille IAP (protéine inhibitrice de l'apoptose). Aux côtés de gènes comme BCL-2 et MCL-1, XIAP joue un rôle critique dans le ciblage de l'apoptose à sa phase d'exécution. Comme souligné dans Genes & Development :
"XIAP est considéré comme l'inhibiteur de caspase le plus puissant in vitro." [12]
XIAP utilise deux mécanismes distincts pour inhiber l'apoptose. Tout d'abord, son domaine BIR2 et sa région de liaison bloquent les caspases effectrices-3 et -7.Deuxièmement, son domaine BIR3 inhibe la caspase-9, arrêtant efficacement la voie apoptotique mitochondriale intrinsèque. De plus, son domaine RING C-terminal facilite l'ubiquitination et la dégradation protéasomale subséquente des caspases cibles [11]. En intervenant dans les voies apoptotiques intrinsèques et extrinsèques, XIAP s'avère très efficace pour traiter les déclencheurs d'apoptose tels que les pénuries de nutriments, les sous-produits métaboliques et le stress mécanique - des facteurs couramment rencontrés dans les systèmes de production de viande cultivée. Sa fonctionnalité est encore renforcée par sa forte conservation à travers les espèces.
Par exemple, XIAP humain partage 87,7 % d'identité protéique avec Bos taurus (bovin) et 89,5 % avec Mus musculus (souris) [11] . Cette grande similarité permet d'appliquer de manière fiable les recherches des systèmes modèles mammifères aux lignées cellulaires utilisées dans la production de viande cultivée.
XIAP peut être régulé à l'aide d'outils tels que shRNA, oligonucléotides antisens ou CRISPR/Cas9 [11]. Sous un stress extrême, son domaine RING peut induire une auto-ubiquitination [12], tandis que des inhibiteurs endogènes comme SMAC/DIABLO et HTRA2 peuvent déplacer XIAP des caspases [11][13]. Ces découvertes font de XIAP une cible attrayante pour les approches d'édition génétique visant à optimiser les lignées cellulaires pour le développement de viande cultivée.
Comparaison des gènes anti-apoptotiques en un coup d'œil
Gènes anti-apoptotiques pour la viande cultivée : comparaison côte à côte
Lors de la production de viande cultivée, comprendre comment fonctionnent différents gènes anti-apoptotiques peut aider à affiner l'ingénierie des lignées cellulaires. Chaque gène a son propre mécanisme distinct, son comportement pendant la différenciation et ses applications potentielles. Le tableau ci-dessous résume ces différences, facilitant ainsi la décision sur le gène - ou la combinaison de gènes - qui pourrait le mieux répondre à vos besoins.
| Gène | Mécanisme Principal | Stabilité de l'Expression | Impact Signalé sur la Viabilité | Compatibilité avec l'Édition |
|---|---|---|---|---|
| BCL-2 | Bloque BAX/BAK pro-apoptotiques et assure la survie des cellules non différenciées [2] | Reste relativement stable pendant la différenciation [2] | Essentiel pour préserver le pool initial de cellules souches [2] | Haute compatibilité avec les outils d'édition |
| BCL-xL | Inhibe la caspase-3 clivée; maintient l'intégrité de la membrane mitochondriale et le métabolisme [2] | Surexprimé à partir du jour 7 de la différenciation [2] | Critique pour soutenir les progéniteurs différenciants ; son inhibition augmente la mort cellulaire [2] | Haute compatibilité avec les outils d'édition |
| MCL-1 | Module les signaux pro-apoptotiques en tant que membre de la famille BCL-2 [2] | L'expression reste stable pendant la spécification de la lignée [2] | Offre de larges avantages de survie mais manque d'effets spécifiques à une étape comme BCL-xL [2] | Haute compatibilité avec les outils d'édition |
| BIRC5 (Survivin) | Bloque les caspases-3 et caspases-7 ; aide à la ségrégation chromosomique pendant la mitose | Élevé dans les cellules proliférantes ; diminue avec la différenciation terminale | Soutient la survie et la prolifération dans les cellules à division rapide | Compatible avec à la fois l'inhibition par shRNA et l'édition CRISPR |
| XIAP | Inhibe plusieurs caspases, offrant une protection apoptotique étendue | Généralement stable dans diverses conditions | Particulièrement efficace sous stress, comme dans des conditions de bioréacteur à haute densité | Haute compatibilité avec les outils d'édition |
BCL-xL se distingue par son double rôle dans la promotion de la survie cellulaire et le soutien de l'activité métabolique, particulièrement pendant la phase critique de différenciation lorsque les protéines pro-apoptotiques comme BAK déclinent naturellement. BCL-2, en revanche, est idéal pour préserver les cellules indifférenciées, tandis que XIAP offre une protection large, surtout dans des environnements stressants comme les cultures à haute densité.
Aucun gène unique ne fonctionne mieux dans tous les scénarios. Par exemple, BIRC5 est particulièrement utile dans les situations nécessitant une division cellulaire rapide. En pratique, combiner deux ou plusieurs gènes offre souvent la protection la plus efficace, répondant simultanément à une variété de déclencheurs apoptotiques.
Ces découvertes fournissent une base pour l'incorporation de ces gènes dans des stratégies d'ingénierie de lignées cellulaires pour la production de viande cultivée. Cela inclut la sélection des bons intrants de viande cultivée pour assurer l'évolutivité.
Utilisation de ces gènes dans l'ingénierie des lignées cellulaires de viande cultivée
Pour améliorer la viabilité cellulaire dans la production de viande cultivée, l'intégration stratégique de gènes clés est cruciale.Il ne suffit pas d'identifier les gènes anti-apoptotiques - leur incorporation efficace dans les lignées cellulaires est ce qui fait la différence. Deux stratégies principales sont couramment employées : surexprimer les gènes anti-apoptotiques comme BCL-2, BCL-xL, et MCL-1 pour améliorer la survie cellulaire, ou éliminer les gènes pro-apoptotiques tels que BAX, BAK, et BOK pour éliminer les moteurs de la mort cellulaire. Combiner ces approches aboutit souvent à des lignées cellulaires mieux adaptées à la production à grande échelle [1].
Les outils modernes d'édition génétique comme CRISPR/Cas9 permettent des modifications simultanées, telles que l'élimination de Bak1, Bax, et Bok en une seule étape. Des alternatives comme ZFNs ou l'interférence ARN peuvent être utilisées pour réduire temporairement l'activité des caspases ( e.g. caspases-3, -7, -8, et -9). Pour les stratégies de surexpression, les promoteurs synthétiques assurent des niveaux d'expression constants et élevés de gènes comme BCL-2 lors de la montée en échelle, ce qui est crucial pour maintenir la performance cellulaire dans les systèmes de culture en fed-batch ou en continu . Ces méthodes combinées renforcent le développement de lignées cellulaires pour les applications de viande cultivée.
De telles modifications génétiques ont un impact direct sur l'amélioration de la concentration cellulaire viable intégrale (IVCC), un indicateur clé dans la production de viande cultivée. La mort cellulaire est la plus prononcée pendant les cinq premiers jours de différenciation, rendant les interventions précoces avec des gènes comme BCL-2 ou BCL-xL essentielles. La recherche publiée dans Cell Death & Disease souligne que l'expression de BCL-xL augmente à mesure que les cellules se différencient, indiquant que les progéniteurs plus matures dépendent fortement de son rôle protecteur [2] . En surveillant les niveaux d'expression des gènes de la famille BCL-2 tout au long des phases de croissance, les interventions peuvent être précisément programmées pour un effet maximal.
"En établissant des lignées cellulaires stables qui surexpriment des gènes anti-apoptotiques ou sous-régulent des gènes pro-apoptotiques, les rendements du produit final peuvent être améliorés car les cellules deviennent plus résistantes aux stress environnementaux." - Gyun Min Lee et al. [1]
Pour la production en bioréacteur, les cellules doivent également être conçues pour résister au stress hyperosmotique et à la privation de nutriments. Avant de passer à l'échelle supérieure, il est essentiel de valider les modifications génétiques à l'aide d'outils comme le Western blot ou le FACS. Pour les chercheurs à la recherche de lignées cellulaires spécialisées ou de matériaux génétiques adaptés aux environnements de bioréacteurs à haute densité, des plateformes comme
Conclusion
La sélection de gènes anti-apoptotiques pour les lignées cellulaires de viande cultivée nécessite une approche sur mesure. Des gènes comme BCL-2, BCL-xL, et MCL-1 jouent chacun des rôles uniques dans la protection des cellules, mais leur succès dépend de facteurs tels que le type de cellule, le stade de développement et les stress spécifiques rencontrés pendant la production. Comme le souligne la recherche :
"l'équilibre entre les membres anti-apoptotiques et pro-apoptotiques détermine en fin de compte si une cellule vit ou meurt" [2]
Au-delà de la survie, l'ingénierie anti-apoptotique préserve également les fonctions métaboliques. Par exemple, des protéines telles que BCL-xL sont étroitement liées au maintien de la glycolyse et de la phosphorylation oxydative. Cependant, des interventions mal exécutées peuvent perturber ces processus critiques [2]. Assurer que les lignées cellulaires ingénierées maintiennent leur identité et activité métabolique prévues tout au long de la production est une étape cruciale, bien que parfois négligée. Ces perspectives façonnent l'avenir de l'ingénierie des lignées cellulaires.
De nouvelles approches multi-gènes émergent, qui combinent la surexpression de gènes protecteurs avec des knockouts CRISPR de gènes pro-apoptotiques comme BAX, BAK1, et BOK pour créer des lignées cellulaires plus robustes pour une utilisation industrielle [1]. Les outils de profilage métabolique, tels que les tests bioénergétiques, deviennent essentiels pour confirmer que ces modifications génétiques améliorent la performance globale des cellules. Pour les chercheurs qui s'approvisionnent en lignées cellulaires porcines , matériaux génétiques, ou équipements de bioréacteur,
FAQs
Avec quel gène anti-apoptotique devrais-je commencer pour ma lignée cellulaire ?
Le BCL-2 est souvent suggéré comme point de départ lors du travail avec des lignées cellulaires. Ce gène anti-apoptotique bien étudié est reconnu pour sa capacité à améliorer la survie cellulaire, ce qui en fait une option populaire dans la recherche sur la viande cultivée. Sa fonction de soutien à la viabilité cellulaire en fait un choix pratique pour les expériences en phase initiale.
Est-il préférable de surexprimer les gènes anti-apoptotiques ou d'éliminer les gènes pro-apoptotiques ?
Dans la production de viande cultivée, augmenter l'expression des gènes anti-apoptotiques, tels que les membres de la famille BCL-2 comme BCL-xL, a tendance à donner de meilleurs résultats que de désactiver les gènes pro-apoptotiques. Cette stratégie soutient à la fois la survie et la prolifération des cellules - des facteurs clés pour l'augmentation de la production - tout en préservant les systèmes de régulation naturels de la cellule.
En augmentant l'activité des gènes anti-apoptotiques, les cellules acquièrent une plus grande résistance à l'apoptose, surtout dans des conditions stressantes. Cela en fait une approche plus fiable et plus sûre pour maintenir la viabilité cellulaire pendant le processus de culture.
Comment puis-je confirmer qu'une modification anti-apoptotique améliore l'IVCC dans mon bioréacteur ?
Pour déterminer si une modification d'un gène anti-apoptotique améliore in vitro la viabilité et la prolifération cellulaire (IVCC), vous aurez besoin d'une approche systématique :
- Évaluer les taux de viabilité et de prolifération: Utilisez des méthodes telles que le comptage cellulaire ou la cytométrie en flux pour mesurer ces taux avant et après la modification du gène.
- Vérifier l'expression génique: Des techniques telles que la qPCR ou le Western blotting peuvent confirmer l'expression réussie du gène ciblé.
- Surveiller les marqueurs d'apoptose: Vérifiez les marqueurs tels que l'activité des caspases pour s'assurer que la modification réduit efficacement l'apoptose.
Pour une évaluation complète, il est essentiel de tester la stabilité à long terme et la prolifération des cellules modifiées dans un bioréacteur. Cela garantit que les améliorations persistent à travers plusieurs cycles de culture.










